
Effects of Supplementing Rumen-Protected Methionine and Lysine on Milk Performance and Oxidative Status of Dairy Ewes

 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Diets and Experimental Design
2.2. Sample Collection
2.3. Feed Analyses
2.4. Milk Chemical Composition and FA Profile
2.5. Antioxidant Enzymes Activities and Oxidative Status Indicators
2.6. Statistics
3. Results
3.1. Animals and Milk Performance
3.2. Milk Fatty Acids Profile
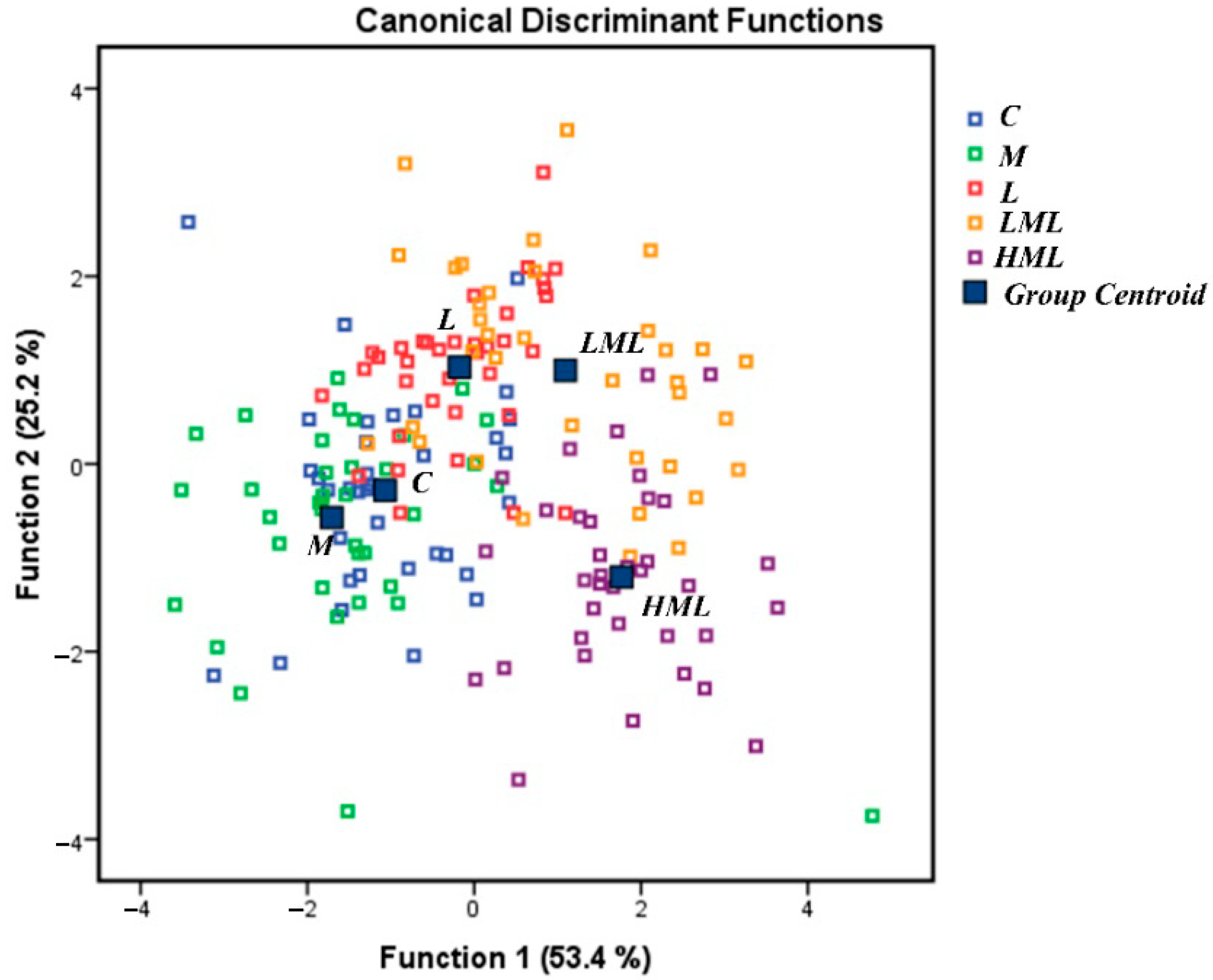
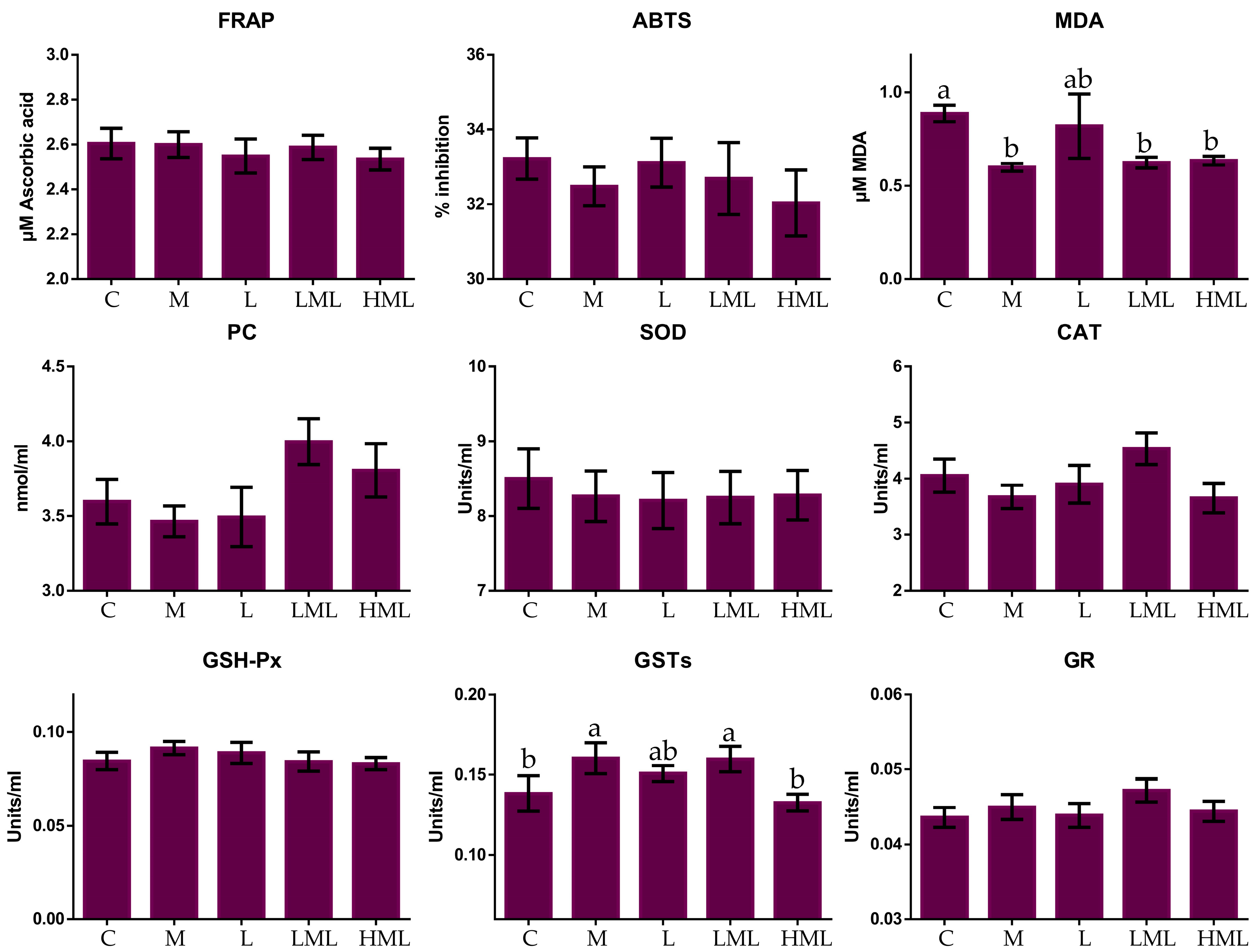
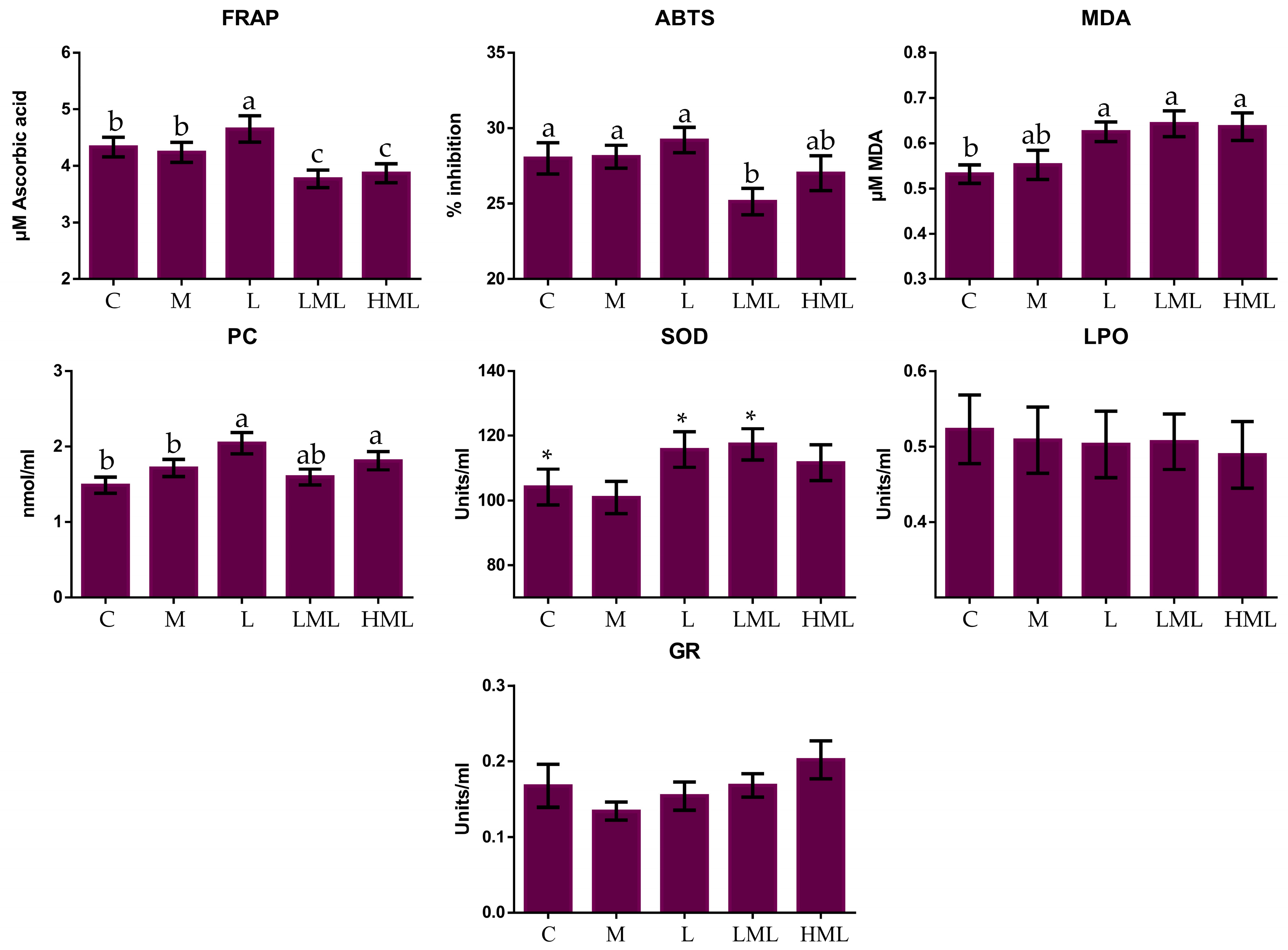
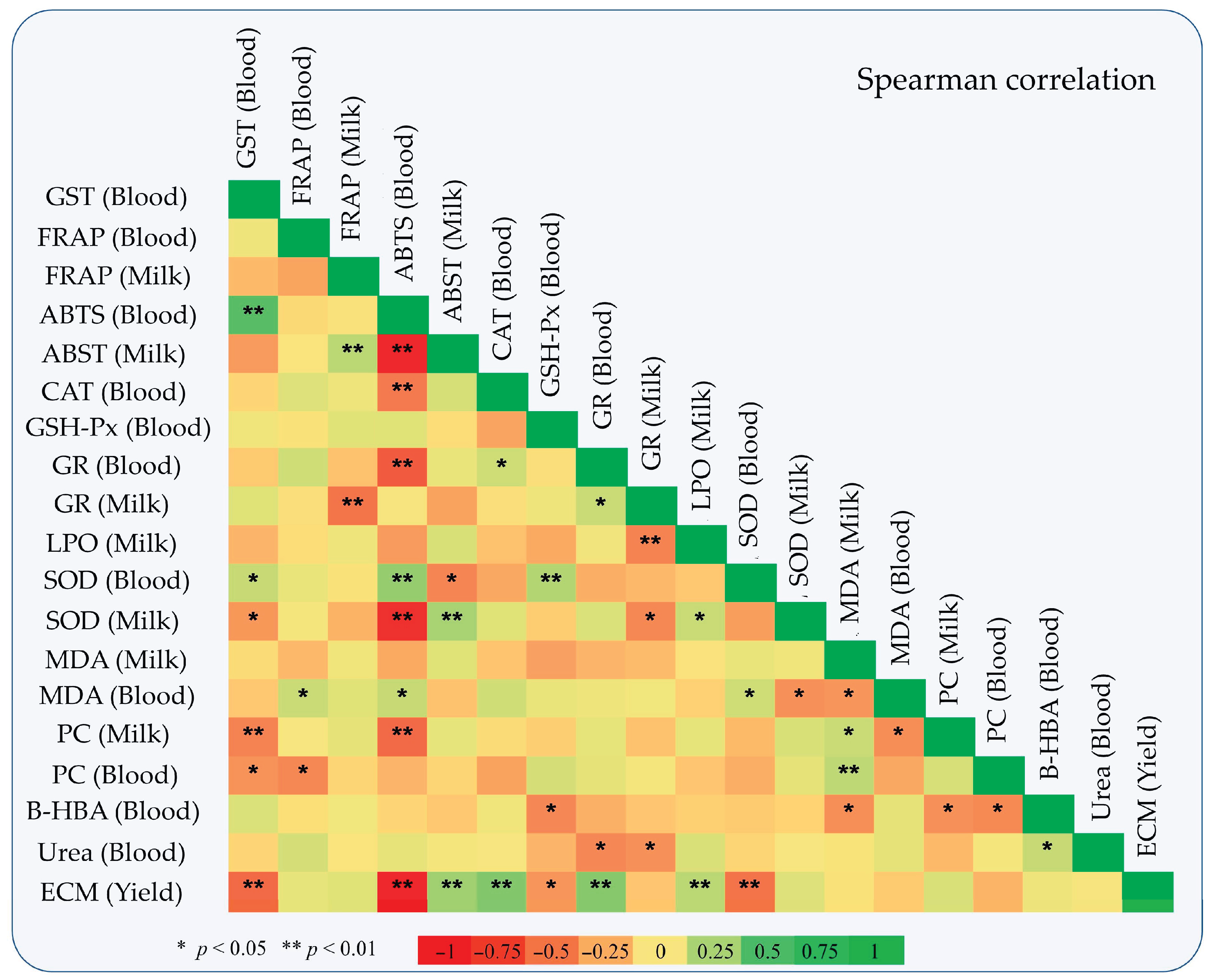
3.3. Oxidative Status
4. Discussion
4.1. Animals’ Performance
4.2. Fatty Acid Profile
4.3. Oxidative Status
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schwab, C.G.; Broderick, G.A. A 100-Year Review: Protein and amino acid nutrition in dairy cows. J. Dairy Sci. 2017, 100, 10094–10112. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.G.; Peng, H.H.; Schwab, C.G. Enhancing the productivity of dairy cows using amino acids. Anim. Prod. Sci. 2013, 53, 1156. [Google Scholar] [CrossRef]
- Tsiplakou, E.; Mavrommatis, A.; Kalogeropoulos, T.; Chatzikonstantinou, M.; Koutsouli, P.; Sotirakoglou, K.; Labrou, N.; Zervas, G. The effect of dietary supplementation with rumen-protected methionine alone or in combination with rumen-protected choline and betaine on sheep milk and antioxidant capacity. J. Anim. Physiol. Anim. Nutr. 2017, 101, 1004–1013. [Google Scholar] [CrossRef] [PubMed]
- Arriola Apelo, S.I.; Bell, A.L.; Estes, K.; Ropelewski, J.; de Veth, M.J.; Hanigan, M.D. Effects of reduced dietary protein and supplemental rumen-protected essential amino acids on the nitrogen efficiency of dairy cows. J. Dairy Sci. 2014, 97, 5688–5699. [Google Scholar] [CrossRef] [PubMed]
- Bröer, S.; Bröer, A. Amino acid homeostasis and signalling in mammalian cells and organisms. Biochem. J. 2017, 474, 1935–1963. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.; Li, T.; Yin, Y. Methionine and Antioxidant Potential. J. Antioxid. Act. 2016, 1, 12–17. [Google Scholar] [CrossRef]
- Bequette, B.J.; Backwell, F.R.; Kyle, C.E.; Calder, A.G.; Buchan, V.; Crompton, L.A.; France, J.; MaCrae, J.C. Vascular sources of phenylalanine, tyrosine, lysine, and methionine for casein synthesis in lactating goats. J. Dairy Sci. 1999, 82, 362–377. [Google Scholar] [CrossRef]
- Glascock, R.F.; Welch, V.A. Contribution of the fatty acids of three low density serum lipoproteins to bovine milk fat. J. Dairy Sci. 1974, 57, 1364–1370. [Google Scholar] [CrossRef]
- Park, J.K.; Yeo, J.M.; Bae, G.S.; Kim, E.J.; Kim, C.H. Effects of supplementing limiting amino acids on milk production in dairy cows consuming a corn grain and soybean meal-based diet. J. Anim. Sci. Technol. 2020, 624, 485–494. [Google Scholar] [CrossRef]
- Chow, J.M.; DePeters, E.J.; Baldwin, R.L. Two Rumen-Protected Amino Acids in Dairy Cows’ Feed Change the Protein Content Of Milk. Calif. Agric. 1990, 44, 12–14. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Yin, Y.L.; Li, D.; Kim, S.W.; Wu, G. Amino acids and immune function. Br. J. Nutr. 2007, 98, 237–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, F.; Cao, Y.; Cai, C.; Li, S.; Yu, C.; Yao, J. Regulation of Nutritional Metabolism in Transition Dairy Cows: Energy Homeostasis and Health in Response to Post-Ruminal Choline and Methionine. PLoS ONE 2016, 11, e0160659. [Google Scholar] [CrossRef] [Green Version]
- Lopes, M.G.; Dominguez, J.H.E.; Corrêa, M.N.; Schmitt, E.; Fischer, G. Rumen-protected methionine in cattle: Influences on reproduction, immune response, and productive performance. Arq. Inst. Biol. 2019, 86, e1292018. [Google Scholar] [CrossRef]
- Awawdeh, M.S. Rumen-protected methionine and lysine: Effects on milk production and plasma amino acids of dairy cows with reference to metabolisable protein status. J. Dairy Res. 2016, 83, 151–155. [Google Scholar] [CrossRef]
- Vailati-Riboni, M.; Zhou, Z.; Jacometo, C.B.; Minuti, A.; Trevisi, E.; Luchini, D.N.; Loor, J.J. Supplementation with rumen-protected methionine or choline during the transition period influences whole-blood immune response in periparturient dairy cows. J. Dairy Sci. 2017, 100, 3958–3968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, D.N.; Lopreiato, V.; Alharthi, A.; Loor, J.J. Amino acids and the regulation of oxidative stress and immune function in dairy cattle. J. Anim. Sci. 2020, 98, S175–S193. [Google Scholar] [CrossRef]
- Toledo, M.Z.; Baez, G.M.; Garcia-Guerra, A.; Lobos, N.E.; Guenther, J.N.; Trevisol, E.; Luchini, D.; Shaver, R.D.; Wiltbank, M.C. Effect of feeding rumen-protected methionine on productive and reproductive performance of dairy cows. PLoS ONE 2017, 12, e0189117. [Google Scholar] [CrossRef] [Green Version]
- Tsiplakou, E.; Mavrommatis, A.; Skliros, D.; Righi, F.; Flemetakis, E. The impact of rumen-protected amino acids on the expression of key- genes involved in the innate immunity of dairy sheep. PLoS ONE 2020, 15, e0233192. [Google Scholar] [CrossRef]
- Tsiplakou, E.; Mavrommatis, A.; Skliros, D.; Sotirakoglou, K.; Flemetakis, E.; Zervas, G. The effects of dietary supplementation with rumen-protected amino acids on the expression of several genes involved in the immune system of dairy sheep. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1437–1449. [Google Scholar] [CrossRef]
- Ali, C.S.; Islam-ud-Din; Sharif, M.; Nisa, M.; Javaid, A.; Hashmi, N.; Sarwar, M. Supplementation of ruminally protected proteins and amino acids: Feed consumption, digestion and performance of cattle and sheep. Int. J. Agric. Biol. 2009, 11, 477–482. [Google Scholar]
- Cannas, A.; Tedeschi, L.O.; Fox, D.G.; Pell, A.N.; Van Soest, P.J. A mechanistic model for predicting the nutrient requirements and feed biological values for sheep1. J. Anim. Sci. 2004, 82, 149–169. [Google Scholar] [CrossRef] [Green Version]
- IDF: Bulletin of the IDF No. 285/1993—Reference Materials and Interlaboratory Collaborative Studies (Third Series), by Various Groups of Experts (See Also Bulletins Nos 207/1986, 235/1988); The International Dairy Federation: Brussels, Belgium, 1993; ISSN 0250-5118.
- The Royal Society of Chemistry. British Standards Institution. Available online: https://pubs.rsc.org/en/Content/ArticleLanding/1952/AN/an952770546a#!divAbstract (accessed on 15 September 2020).
- ISO (International Organization for Standardization). Milk—Determination of Casein-Nitrogen Content—Part 1: Indirect Method (ISO 17997-1/IDF 29); ISO: Geneva, Switzerland, 2004. [Google Scholar]
- Bocquier, F.; Barillet, F.; Guillouet, P.; Jacquin, M. Pre’vision de l’energie du lair de brebis a` partir de diffe’rents re’sultats d’ analyses: Proposition de lait standard pour les brebis laitie`res. Ann. Zootech. 1993, 42, 57–66. [Google Scholar] [CrossRef]
- Mavrommatis, A.; Tsiplakou, E. The impact of the dietary supplementation level with Schizochytrium sp. on milk chemical composition and fatty acid profile, of both blood plasma and milk of goats. Small Rumin. Res. 2020, 106252. [Google Scholar] [CrossRef]
- Tsiplakou, E.; Mitsiopoulou, C.; Mavrommatis, A.; Karaiskou, C.; Chronopoulou, E.G.; Mavridis, G.; Sotirakoglou, K.; Labrou, N.E.; Zervas, G. Effect of under- and overfeeding on sheep and goat milk and plasma enzymes activities related to oxidation. J. Anim. Physiol. Anim. Nutr. 2018, 102, 288–298. [Google Scholar] [CrossRef]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar]
- Nielsen, F.; Mikkelsen, B.B.; Nielsen, J.B.; Andersen, H.R.; Grand-Jean, P. Plasma malondialdehyde as bio-marker for oxidative stress: Reference interval and effects of life-style factors. Clin. Chem. 1997, 43, 1209–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mavrommatis, A.; Mitsiopoulou, C.; Christodoulou, C.; Karabinas, D.; Nenov, V.; Zervas, G.; Tsiplakou, E. Dietary Supplementation of a Live Yeast Product on Dairy Sheep Milk Performance, Oxidative and Immune Status in Peripartum Period. J. Fungi 2020, 6, 334. [Google Scholar] [CrossRef] [PubMed]
- Patsoukis, N.; Zervoudakis, G.; Panagopoulos, N.T.; Georgiou, C.D.; Angelatou, F.; Matsokis, N.A. Thiol redox state (TRS) and oxidative stress in the mouse hippocampus after pentylenetetrazolinduced epileptic seizure. Neurosci. Lett. 2004, 357, 83–86. [Google Scholar] [CrossRef]
- Pellegrini, N.; Serafini, M.; Colombi, B.; Del Rio, D.; Salvatore, S.; Bianchi, M.; Brighenti, F. Total antioxidant capacity of plant foods, beverages and oils consumed in Italy assessed by three different in vitro assays. J. Nutr. 2003, 133, 2812–2819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Huo, L.; Su, W.; Lu, R.; Deng, C.; Li, L.; He, C. Free radical scavenging capacity, antioxidant activity and phenolic content of Pouzolzia zeylanica. J. Serbian Chem. Soc. 2011, 76, 709–717. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, J.A.; Horton, G.M.J.; Wohlt, J.E.; Palatini, D.D.; Emanuele, S.M. Rumen-protected methionine for lactation, wool and growth in sheep. Small Rumin. Res. 1993, 12, 125–132. [Google Scholar] [CrossRef]
- Lara, A.; Mendoza, G.D.; Landois, L.; Barcena, R.; Sánchez-Torres, M.T.; Rojo, R.; Vega, S. Milk production in Holstein cows supplemented with different levels of ruminally protected methionine. Livest. Sci. 2006, 105, 105–108. [Google Scholar] [CrossRef]
- Třináctý, J.; Křížová, L.; Richter, M.; Černý, V.; Říha, J. Effect of rumen-protected methionine, lysine or both on milk production and plasma amino acids of high-yielding dairy cows. Czech J. Anim. Sci. 2009, 54, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.H.; Broderick, G.A.; Luchini, N.D.; Sloan, B.K.; Devillard, E. Effect of feeding different sources of rumen-protected methionine on milk production and N-utilization in lactating dairy cows1. J. Dairy Sci. 2011, 94, 1978–1988. [Google Scholar] [CrossRef] [PubMed]
- Rulquin, H.; Vérité, R. Amino acid nutrition of dairy cows: Productive effects and animal requirements. In Recent Developments in Animal Nutrition; Garnsworthy, P.C., Cole, D.J.A., Eds.; Nottingham University Press: Nottingham, UK, 1996; pp. 71–94. [Google Scholar]
- Oldham, J.D. Protein requirement system for ruminants. In Progress in Dairy Science; Phillips, C.J.C., Ed.; CAB International: Wallingford, UK, 1996; pp. 3–27. [Google Scholar]
- St-Gelais, D.; Haché, S. Effect of β-casein concentration in cheese milk on rennet coagulation properties, cheese composition and cheese ripening. Food Res. Int. 2005, 38, 523–531. [Google Scholar] [CrossRef]
- Goulas, C.; Zervas, G.; Papadopoulos, G. Effect of dietary animal fat and methionine on dairy ewes milk yield and milk composition. Anim. Feed Sci. Technol. 2003, 105, 43–54. [Google Scholar] [CrossRef]
- Flores, A.; Mendoza, G.; Pinos-Rodrıguez, J.M.; Plata, F.; Vega, S.; Barcena, R. Effects of rumen-protected methionine on milk production of dairy goats. Ital. J. Anim. Sci. 2009, 8, 271–275. [Google Scholar] [CrossRef]
- Poljicak-Milas, N.; Marenjak, T.S. Dietary supplement of the rumen protected methionine and milk yield in dairy Goats. Arch. Tierz. 2007, 50, 273–278. [Google Scholar] [CrossRef] [Green Version]
- Al-Qaisi, M.A.; Titi, H.H. Effect of rumen-protected methionine on production and composition of early lactating Shami goats milk and growth performance of their kids. Arch. Tierz. 2014, 57, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kudrna, V.; Illek, J.; Marounek, M.; Nguyen Ngoc, A. Feeding ruminally protected methionine to pre- and postpartum dairy cows: Effect on milk performance, milk composition and blood parameters. Czech J. Anim. Sci. 2009, 9, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.R.; Sun, H.; Wang, Q.Y.; Liu, F.X.; Yang, Z.B. Effects of rumen protected methionine on dairy performance and amino acid metabolism in lactating cows. Am. J. Anim. Vet. Sci. 2010, 5, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Soltan, M.A.; Mujalli, A.M.; Mandour, M.A.; El-Shinaway Abeer, M. Effect of dietary rumen protected methionine and/or choline supplementation on rumen fermentation characteristics and productive performance of early lactating cows. Pak. J. Nutr. 2012, 11, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Mepham, T.B. Amino acid utilization by lactating mammary gland. J. Dairy Sci. 1982, 65, 287–298. [Google Scholar] [CrossRef]
- Bequette, B.J.; Backwell, F.R.; Crompton, L.A. Current concepts of amino acid and protein metabolism in the mammary gland of the lactating ruminant. J. Dairy Sci. 1998, 81, 2540–2559. [Google Scholar] [CrossRef]
- Bionaz, M.; Periasamy, K.; Rodriguez-Zas, S.L.; Everts, R.E.; Lewin, H.A.; Hurley, W.L.; Loor, J.J. Old and new stories: Revelations from functional analysis of the bovine mammary transcriptome during the lactation cycle. PLoS ONE 2012, 7, e33268. [Google Scholar] [CrossRef]
- Garnsworthy, P.C.; Masson, L.L.; Lock, A.L.; Mottram, T.T. Variation of milk citrate with stage of lactation and de novo fatty acid synthesis in dairy cow. J. Dairy Sci. 2006, 89, 1604–1612. [Google Scholar] [CrossRef] [Green Version]
- Faulkner, A.; Peaker, M. Reviews of the progress of dairy science: Secretion of citrate into milk. J. Dairy Res. 1982, 49, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Sevi, A.; Rotunno, T.; Di Caterina, R.; Muscio, A. Rumen-protected methionine or lysine supplementation of Comisana ewes’ diets: Effects on milk fatty acid composition. J. Dairy Res. 1998, 65, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Giallongo, F.; Harper, M.T.; Oh, J.; Lopes, J.C.; Lapierre, H.; Patton, R.A.; Parys, C.; Shinzato, I.; Hristov, A.N. Effects of rumen-protected methionine, lysine, and histidine on lactation performance of dairy cows. J Dairy Sci. 2016, 99, 4437–4452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, L.; Xiao, X.X.; LIU, Y.; Jiang, J.; Hu, K.; Jiang, W.D.; Li, S.H.; Zhou, X.Q. Methionine hydroxy analogue prevents oxidative damage and improves antioxidant status of intestine and hepatopancreas for juvenile Jian carp (Cyprinus carpio var. Jian). Aquac. Nutr. 2011, 17, 595–604. [Google Scholar] [CrossRef]
- Osorio, J.S.; Trevisi, E.; Ji, P.; Drackley, J.K.; Luchini, D.; Bertoni, G.; Loor, J.J. Biomarkers of inflammation, metabolism, and oxidative stress in blood, liver, and milk reveal a better immunometabolic status in peripartal cows supplemented with Smartamine M or MetaSmart. J. Dairy Sci. 2014, 97, 7437–7450. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.T.; Chen, H.W.; Sheen, L.Y.; Lii, C.K. Methionine and Cysteine Affect Glutathione Level, Glutathione-Related Enzyme Activities and the Expression of Glutathione S-Transferase Isozymes in Rat Hepatocytes. J. Nutr. 1997, 127, 2135–2141. [Google Scholar] [CrossRef]
- D’Angelo, A.; Selhub, J. Homocysteine and thrombotic disease. Blood 1997, 90, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pizzorno, J. Glutathione! Integr. Med. (Encinitas) 2014, 13, 8–12. [Google Scholar] [PubMed]
- Gould, R.L.; Pazdro, R. Impact of Supplementary Amino Acids, Micronutrients, and Overall Diet on Glutathione Homeostasis. Nutrients 2019, 11, 1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jankowski, J.; Ognik, K.; Kubińska, M.; Czech, A.; Juśkiewicz, J.; Zduńczyk, Z. The effect of DL-, L-isomers and DL-hydroxy analog administered at 2 levels as dietary sources of methionine on the metabolic and antioxidant parameters and growth performance of turkeys. Poult. Sci. 2017, 96, 3229–3238. [Google Scholar] [CrossRef] [PubMed]
- Kapusta, A.; Kuczyńska, B.; Puppel, K. Relationship between the degree of antioxidant protection and the level of malondialdehyde in high-performance Polish Holstein-Friesian cows in peak of lactation. PLoS ONE 2018, 13, e0193512. [Google Scholar] [CrossRef] [Green Version]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef] [PubMed]
- Jové, M.; Mota-Martorell, N.; Pradas, I.; Martín-Gari, M.; Ayala, V.; Pamplona, R. The Advanced Lipoxidation End-Product Malondialdehyde-Lysine in Aging and Longevity. Antioxidants 2020, 9, 1132. [Google Scholar] [CrossRef]
- Chio, K.S.; Tappel, A.L. Synthesis and characterization of the fluorescent products derived from malonaldehyde and amino acids. Biochemistry 1969, 8, 2821–2826. [Google Scholar] [CrossRef] [PubMed]
- Sayre, L.M.; Moreira, P.I.; Smith, M.A.; Perry, G. Metal ions and oxidative protein modification in neurological disease. Ann. Ist. Super. Sanita 2005, 41, 143–164. [Google Scholar] [PubMed]
- Yuan, S.B.; Chen, D.W.; Zhang, K.Y.; Yu, B. Effects of oxidative stress on growth performance, nutrient digestibilities and activities of antioxidative enzymes of weanling pigs. Asian Australas. J. Anim. Sci. 2007, 20, 1600–1605. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | |||||
|---|---|---|---|---|---|
| C | M | L | LML | HML | |
| Diet components (% Dry Matter) | |||||
| Alfalfa hay 1st cut | 35.36 | 35.3 | 35.31 | 35.24 | 35.17 |
| Wheat straw | 19,00 | 18.96 | 18.97 | 18.93 | 18.89 |
| Concentrate mix | 45.63 | 45.54 | 45.55 | 45.46 | 45.37 |
| Rumen-protected methionine commercial product | - | 0.2 | - | 0.2 | 0.4 |
| Rumen-protected lysine commercial product | - | - | 0.17 | 0.17 | 0.17 |
| Diet chemical composition (% Dry Matter) | |||||
| Dry Matter | 90.39 | 90.40 | 90.40 | 90.41 | 90.41 |
| Ash | 6.85 | 6.91 | 6.86 | 6.92 | 6.99 |
| Ash-free NDF treated with amylase | 38.01 | 37.93 | 37.94 | 37.86 | 37.86 |
| Acid Detergent Fiber | 26.29 | 26.24 | 26.24 | 26.20 | 26.14 |
| ADL (lignin) * | 5.61 | 5.60 | 5.60 | 5.59 | 5.59 |
| Non Fibrous Carbohydrate | 39.69 | 39.64 | 39.63 | 39.57 | 39.57 |
| Starch * | 23.03 | 23.01 | 23.00 | 22.98 | 22.98 |
| Sugars * | 3.22 | 3.22 | 3.22 | 3.21 | 3.21 |
| Ether Extract | 1.67 | 1.66 | 1.75 | 1.75 | 1.74 |
| Crude Protein | 13.92 | 13.99 | 13.97 | 14.03 | 14.03 |
| Soluble Crude Protein * | 3.69 | 3.78 | 3.69 | 3.78 | 3.78 |
| Rumen Degradable Protein at 3 times the maintenance intake * | 8.92 | 8.90 | 8.94 | 8.92 | 8.92 |
| Methionine %, (g/d) * | 0.23 (6.5) | 0.35 (7.8) | 0.23 (6.5) | 0.35 (7.8) | 0.35 (9.2) |
| Methionine, % Metabolizable Protein * | 2.18 | 2.60 | 2.17 | 2.59 | 3.04 |
| Lysine %, (g/d) * | 0.60 (20.3) | 0.60 (20.3) | 0.72 (21.3) | 0.72 (21.3) | 0.72 (21.3) |
| Lysine, % Metabolizable Protein * | 6.81 | 6.77 | 7.12 | 7.08 | 7.05 |
| Lysine: Methionine * | 3.13:1 | 2.60:1 | 3.28:1 | 2.72:1 | 2.33:1 |
| Net Energy of lactation (Mcal/kg) * | 1.53 | 1.53 | 1.53 | 1.53 | 1.53 |
| Treatment (T) | Sampling Time (S) | Effect b | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C | M | L | LML | HML | SEM a | 25 | 50 | 75 | SEM a | T | S | T × S | |
| Milk yield (kg) | 1.68 | 1.84 | 1.64 | 1.72 | 1.60 | 0.090 | 2.13 A | 1.52 B | 1.44 C | 0.058 | 0.439 | <0.001 | 0.996 |
| FCM6% (kg) | 1.60 | 1.71 | 1.73 | 1.82 | 1.78 | 0.092 | 2.17 A | 1.62 B | 1.42 C | 0.045 | 0.588 | <0.001 | 0.485 |
| ΕCM (kg) | 1.46 | 1.56 | 1.56 | 1.64 | 1.63 | 0.086 | 1.92 A | 1.49 A | 1.30 B | 0.040 | 0.570 | <0.001 | 0.955 |
| Fat content % | 5.64 c | 6.18 b | 6.60 ab | 6.34 ab | 6.72 a | 0.160 | 6.17 B | 6.70 A | 6.02 B | 0.105 | <0.001 | <0.001 | 0.389 |
| Fat yield (g) | 94.34 | 108.18 | 106.17 | 110.35 | 108.58 | 5.426 | 129.58 A | 101.14 B | 85.87 C | 3.014 | 0.216 | <0.001 | 0.223 |
| Protein content % | 5.98 b | 6.72 a | 6.56 a | 6.18 b | 6.23 b | 0.099 | 6.04 B | 6.29 A | 6.32 A | 0.067 | 0.002 | <0.001 | 0.001 |
| Protein yield (g) | 99.35 | 112.36 | 105.40 | 108.32 | 100.71 | 5.028 | 128.09 A | 95.94 B | 91.66 C | 2.384 | 0.401 | <0.001 | 0.136 |
| Caseins (%) | 4.69 b | 4.82 ab | 5.16 a | 4.89 ab | 5.09 ab | 0.033 | 4.83 B | 4.98 A | 4.98 A | 0.017 | 0.039 | <0.001 | 0.894 |
| Lactose % | 5.85 | 5.87 | 5.76 | 5.86 | 5.83 | 0.059 | 6.05 A | 5.75 B | 5.69 C | 0.033 | 0.661 | <0.001 | 0.003 |
| Total solid % | 18.27 c | 19.05 b | 19.68 ab | 19.18 b | 19.85 a | 0.235 | 19.10 A | 19.68 B | 18.85 A | 0.133 | <0.001 | <0.001 | 0.546 |
| Total solid no-fat % | 12.62 b | 12.87 ab | 13.07 a | 12.83 ab | 13.12 a | 0.104 | 12.93 AB | 12.98 A | 12.82 B | 0.056 | 0.011 | <0.001 | 0.606 |
| Body weight (Kg) | 63.75 | 61.79 | 61.04 | 61.81 | 66.33 | 1.744 | 62.91 | - | 62.98 | 0.792 | 0.212 | 0.798 | 0.510 |
| Treatment (T) | Sampling Time a (S) | Effect c | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C | M | L | LML | HML | SEM b | 25 | 50 | 75 | SEM b | T | S | T × S | |
| C4:0 | 2.88 bc | 3.16 a | 3.05 ab | 2.67 c | 2.05 d | 0.077 | 2.60 C | 2.76 B | 2.97 A | 0.052 | <0.001 | <0.001 | <0.001 |
| C6:0 | 2.77 a | 2.80 a | 2.81 a | 2.68 a | 2.28 b | 0.056 | 2.64 | 2.65 | 2.72 | 0.033 | <0.001 | 0.113 | <0.001 |
| C8:0 | 2.93 | 2.87 | 2.89 | 3.01 | 2.83 | 0.069 | 3.09 A | 2.84 B | 2.78 B | 0.036 | 0.409 | <0.001 | 0.409 |
| C10:0 | 9.54 | 9.45 | 9.52 | 9.9 | 9.52 | 0.188 | 10.09 A | 9.33 B | 9.35 B | 0.111 | 0.398 | <0.001 | 0.368 |
| C11:0 | 0.43 | 0.41 | 0.44 | 0.42 | 0.41 | 0.019 | 0.40 B | 0.42 B | 0.44 A | 0.009 | 0.638 | <0.001 | 0.404 |
| C12:0 | 5.34 | 5.53 | 5.51 | 5.78 | 5.64 | 0.167 | 5.87 A | 5.47 B | 5.33 B | 0.090 | 0.423 | <0.001 | 0.347 |
| C13:0 | 0.09 | 0.10 | 0.10 | 0.11 | 0.12 | 0.015 | 0.10 | 0.11 | 0.10 | 0.009 | 0.506 | 0.428 | 0.080 |
| C14:0 | 12.51 | 12.64 | 12.54 | 12.71 | 12.68 | 0.259 | 11.99 C | 12.37 B | 13.49 A | 0.131 | 0.977 | <0.001 | 0.263 |
| C14:1 | 0.48 b | 0.57 a | 0.44 b | 0.36 c | 0.42 b | 0.021 | 0.51 A | 0.44 B | 0.41 B | 0.013 | <0.001 | <0.001 | <0.001 |
| C15:0 | 1.02 | 1.07 | 1.05 | 1.05 | 1.15 | 0.039 | 1.05 B | 1.04 B | 1.11 A | 0.020 | 0.238 | <0.001 | 0.174 |
| C15:1 | 0.29 | 0.31 | 0.30 | 0.30 | 0.31 | 0.013 | 0.30 | 0.30 | 0.30 | 0.007 | 0.686 | 0.514 | 0.168 |
| C16:0 | 28.59 | 28.04 | 28.60 | 27.83 | 28.17 | 0.455 | 26.68 B | 27.02 B | 31.04 A | 0.248 | 0.691 | <0.001 | 0.381 |
| C16:1 | 1.53 | 1.51 | 1.67 | 1.56 | 1.63 | 0.053 | 1.43 C | 1.59 B | 1.72 A | 0.030 | 0.157 | <0.001 | 0.232 |
| C17:0 | 0.74 | 0.76 | 0.80 | 0.75 | 0.82 | 0.046 | 0.84 A | 0.86 A | 0.61 B | 0.024 | 0.707 | <0.001 | 0.550 |
| C17:1 | 0.28 | 0.28 | 0.30 | 0.28 | 0.31 | 0.017 | 0.32 A | 0.32 A | 0.23 B | 0.009 | 0.704 | <0.001 | 0.531 |
| C18:0 | 5.84 | 6.19 | 5.55 | 6.11 | 6.51 | 0.247 | 6.12 B | 6.36 A | 5.64 C | 0.129 | 0.066 | <0.001 | 0.208 |
| C18:1 Σ trans d | 0.46 | 0.42 | 0.42 | 0.44 | 0.44 | 0.019 | 0.43 | 0.45 | 0.42 | 0.011 | 0.511 | 0.121 | 0.023 |
| C18:1 t11 | 1.55 | 1.32 | 1.25 | 1.28 | 1.18 | 0.092 | 1.41 A | 1.34 A | 1.20 B | 0.047 | 0.068 | <0.001 | 0.859 |
| C18:1 c9 | 16.87 | 17.06 | 17.78 | 17.46 | 17.92 | 0.475 | 18.23 A | 18.54 A | 15.24 B | 0.295 | 0.577 | <0.001 | 0.294 |
| C18:2n6t | 0.32 a | 0.29 ab | 0.28 b | 0.27 b | 0.27 b | 0.012 | 0.30 A | 0.29 AB | 0.27 B | 0.007 | 0.021 | <0.001 | 0.051 |
| C18:2n6c | 3.38 | 3.29 | 3.14 | 3.14 | 3.30 | 0.120 | 3.58 A | 3.42 B | 2.76 C | 0.058 | 0.558 | <0.001 | 0.067 |
| C18:3n6 | 0.03 a | 0.01 b | 0.01 bc | 0.03 a | 0.03 ac | 0.007 | 0.04 A | 0.02 A | 0.01 B | 0.004 | 0.010 | <0.001 | 0.024 |
| C20:0 | 0.11 a | 0.10 ab | 0.10 ab | 0.10 b | 0.09 c | 0.004 | 0.10 B | 0.09 C | 0.11 A | 0.003 | 0.002 | <0.001 | <0.001 |
| C18:3n3 | 0.81 b | 0.82 ab | 0.78 b | 0.74 b | 0.90 a | 0.030 | 0.79 B | 0.84 A | 0.80 B | 0.015 | 0.007 | <0.001 | <0.001 |
| c9, t11 C18:2 | 0.94 | 0.80 | 0.84 | 0.79 | 0.76 | 0.055 | 0.83 B | 0.88 A | 0.76 C | 0.028 | 0.207 | <0.001 | 0.170 |
| C20:3n3 | 0.29 a | 0.28 a | 0.26 a | 0.22 b | 0.28 a | 0.011 | 0.27 A | 0.25 B | 0.28 A | 0.007 | <0.001 | 0.006 | <0.001 |
| SCFA | 18.41 a | 18.43 a | 18.55 a | 18.55 a | 16.95 b | 0.305 | 18.83 A | 18.00 B | 17.72 B | 0.182 | 0.001 | <0.001 | 0.034 |
| ΜCFA | 47.55 | 47.37 | 47.80 | 47.49 | 47.76 | 0.672 | 45.69 B | 46.02 B | 51.07 A | 0.357 | 0.991 | <0.001 | 0.131 |
| LCFA | 6.68 | 7.05 | 6.44 | 6.96 | 7.42 | 0.268 | 7.06 A | 7.31 A | 6.36 B | 0.140 | 0.099 | <0.001 | 0.237 |
| ΜUFA | 21.44 | 21.47 | 21.75 | 21.67 | 22.20 | 0.500 | 22.63 A | 22.98 A | 19.51 B | 0.299 | 0.824 | <0.001 | 0.247 |
| PUFA | 5.7 | 5.49 | 5.31 | 5.19 | 5.53 | 0.186 | 5.80 A | 5.70 A | 4.87 B | 0.090 | 0.241 | <0.001 | 0.023 |
| SFA | 27.20 | 26.96 | 27.06 | 26.86 | 27.74 | 0.583 | 28.43 A | 28.68 A | 24.38 B | 0.329 | 0.842 | <0.001 | 0.152 |
| UFA | 72.65 | 72.86 | 72.78 | 72.99 | 72.12 | 0.582 | 71.57 B | 71.32 B | 75.16 A | 0.328 | 0.849 | <0.001 | 0.139 |
| SFA/UFA | 2.72 | 2.76 | 2.76 | 2.77 | 2.64 | 0.080 | 2.57 B | 2.51 B | 3.11 A | 0.043 | 0.768 | <0.001 | 0.039 |
| AI | 3.15 | 3.20 | 3.22 | 3.22 | 3.11 | 0.115 | 2.90 B | 2.89 B | 3.74 A | 0.060 | 0.948 | <0.001 | 0.051 |
| Δ−9 desaturase index | |||||||||||||
| C14:1/C14:0 | 0.04 b | 0.05 a | 0.04 bc | 0.03 d | 0.03 c | 0.001 | 0.04 A | 0.04 B | 0.03 C | 0.001 | <0.001 | <0.001 | <0.001 |
| C16:1/C16:0 | 0.05 | 0.05 | 0.06 | 0.06 | 0.06 | 0.002 | 0.05 B | 0.06 A | 0.06 B | 0.001 | 0.160 | <0.001 | 0.005 |
| C18:1 c9/C18:0 | 2.92 ab | 2.78 b | 3.18 a | 2.89 b | 2.80 b | 0.094 | 3.02 A | 2.97 A | 2.75 B | 0.054 | 0.035 | <0.001 | 0.752 |
| c9, t11C18:2/t11 C18:1 | 0.61 b | 0.62 ab | 0.69 a | 0.62 b | 0.67 ab | 0.020 | 0.60 B | 0.67 A | 0.65 A | 0.012 | 0.030 | <0.001 | 0.663 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mavrommatis, A.; Mitsiopoulou, C.; Christodoulou, C.; Kariampa, P.; Simoni, M.; Righi, F.; Tsiplakou, E. Effects of Supplementing Rumen-Protected Methionine and Lysine on Milk Performance and Oxidative Status of Dairy Ewes. Antioxidants 2021, 10, 654. https://doi.org/10.3390/antiox10050654
Mavrommatis A, Mitsiopoulou C, Christodoulou C, Kariampa P, Simoni M, Righi F, Tsiplakou E. Effects of Supplementing Rumen-Protected Methionine and Lysine on Milk Performance and Oxidative Status of Dairy Ewes. Antioxidants. 2021; 10(5):654. https://doi.org/10.3390/antiox10050654
Chicago/Turabian StyleMavrommatis, Alexandros, Christina Mitsiopoulou, Christos Christodoulou, Paraskevi Kariampa, Marica Simoni, Federico Righi, and Eleni Tsiplakou. 2021. "Effects of Supplementing Rumen-Protected Methionine and Lysine on Milk Performance and Oxidative Status of Dairy Ewes" Antioxidants 10, no. 5: 654. https://doi.org/10.3390/antiox10050654