
CTLA-2 Alpha Is a Potent Inhibitor of Angiogenesis in Murine Ocular Tissue

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Hybridization Probes
2.3. Recombinant CTLA-2 Alpha
2.4. Anti-CTLA-2 Alpha Antibody
2.5. Corneal Suture Placement
2.6. Laser-Induced CNV
2.7. Determination of Hemangiogenesis
2.8. Determination of CNV
2.9. Mouse Pituitary Vasuclar Endothelail Cell Culture
2.10. Collection and Culture of Peritoneal Exudate Cells from the Peritoneal Cavity
2.11. ELISA for TNF-Alpha
2.12. Cell Proliferation Assay
2.13. CTLA-2 Alpha and Cath L Overexpression in mVECs
2.14. Statistical Analysis
3. Results
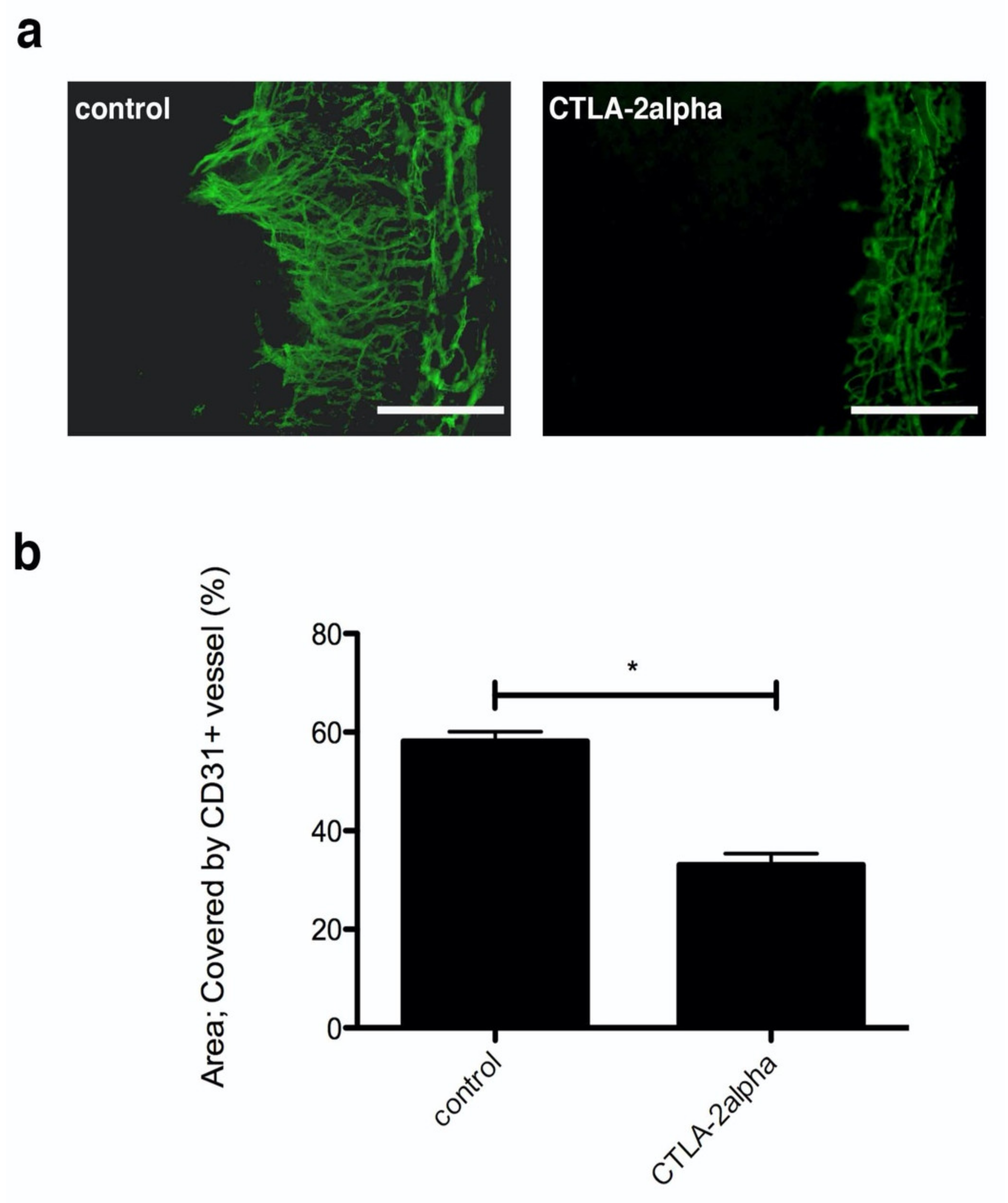
3.1. The Effects of CTLA-2 Alpha in the Corneal Suture Model
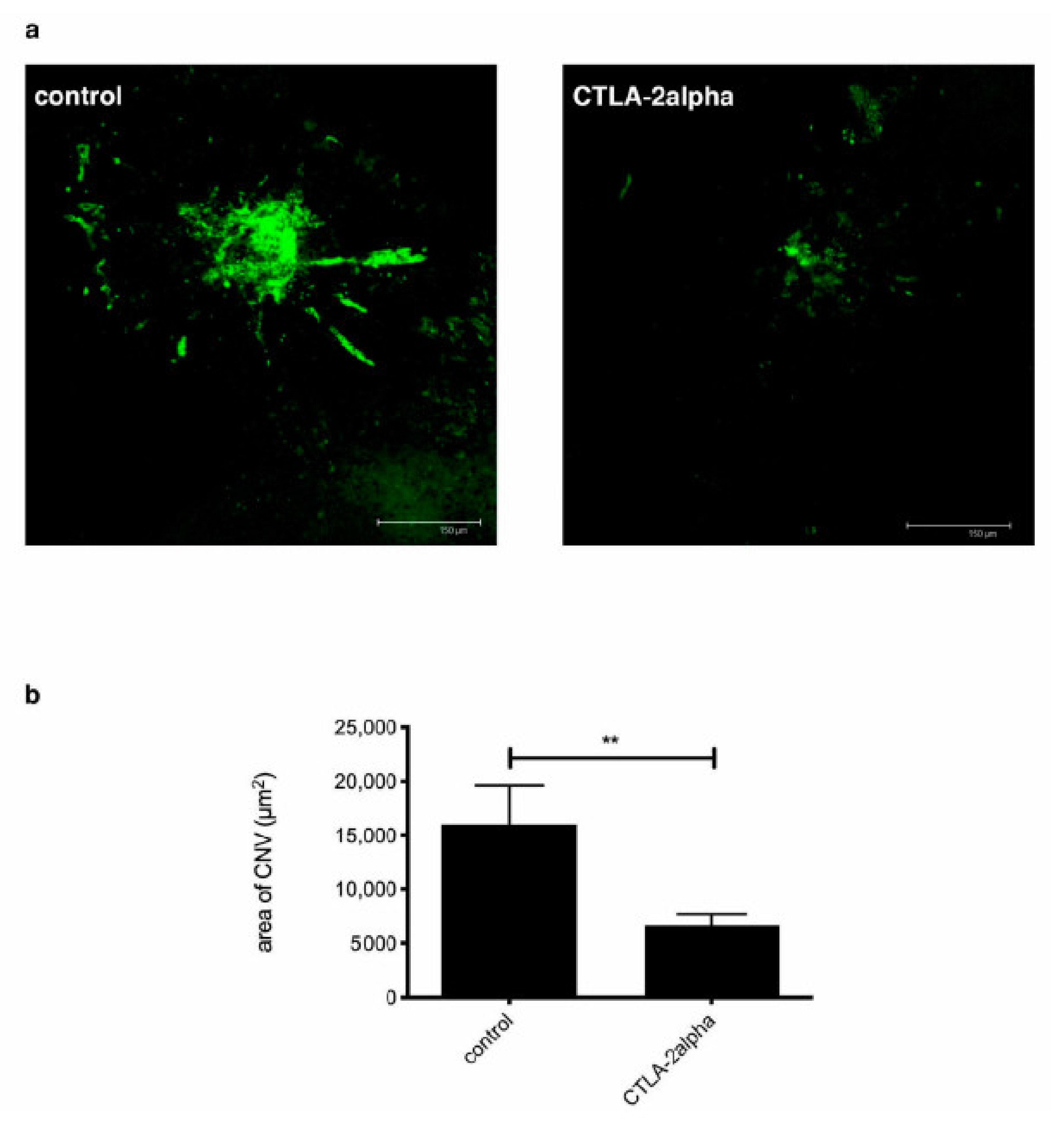
3.2. The Effects of CTLA-2 Alpha in the CNV Model
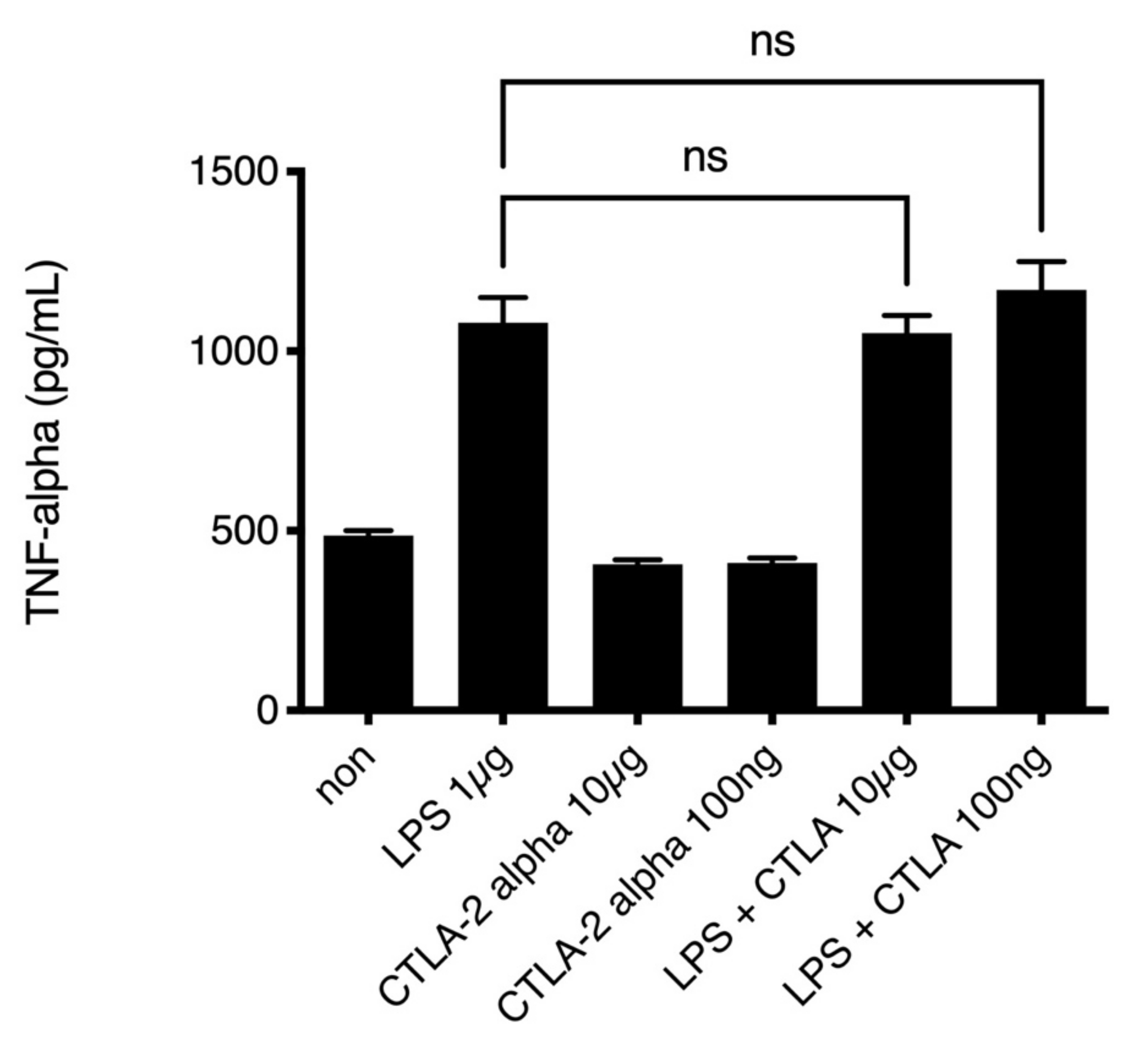
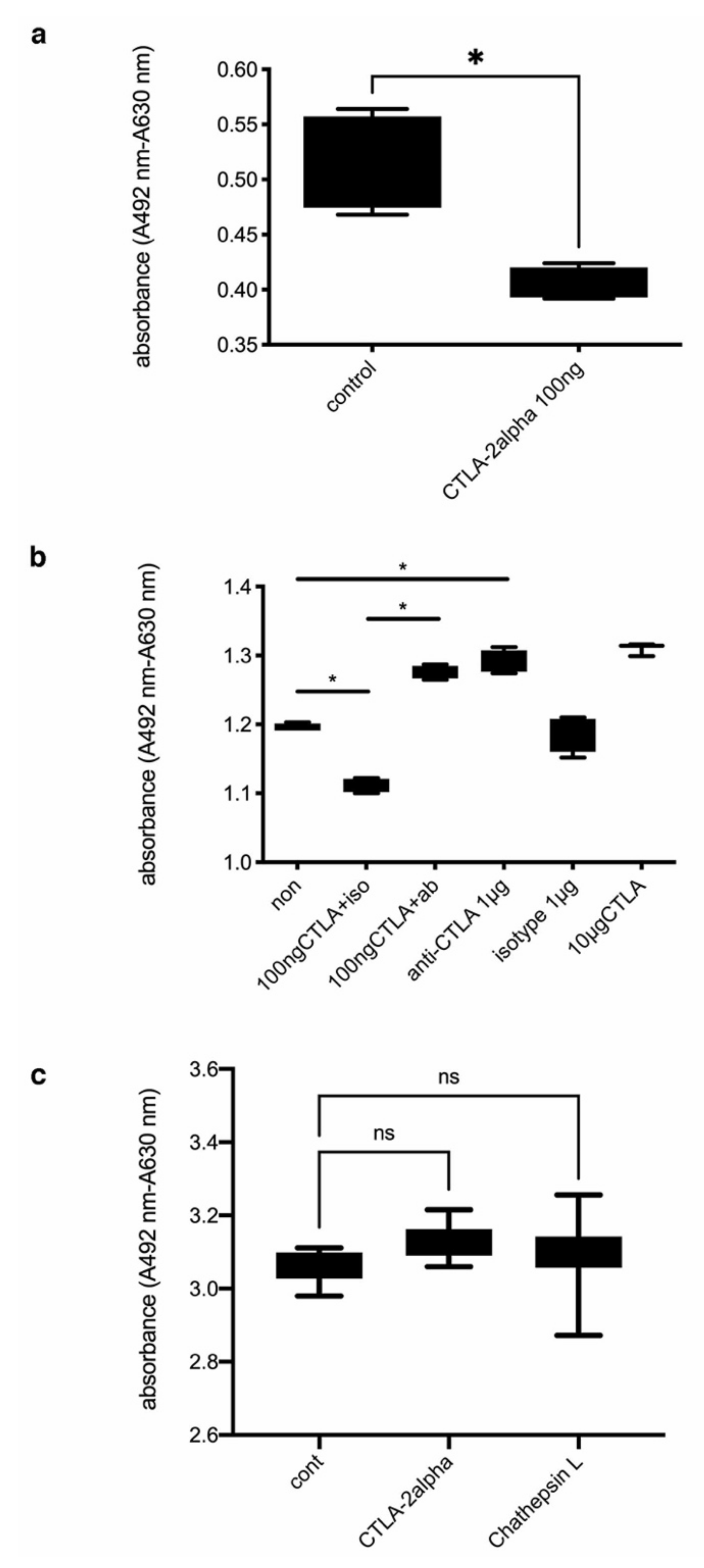
3.3. The Effects of CTLA-2 Alpha on PEC Activation In Vitro
3.4. The Effects of CTLA-2 Alpha on mVEC Proliferation In Vitro
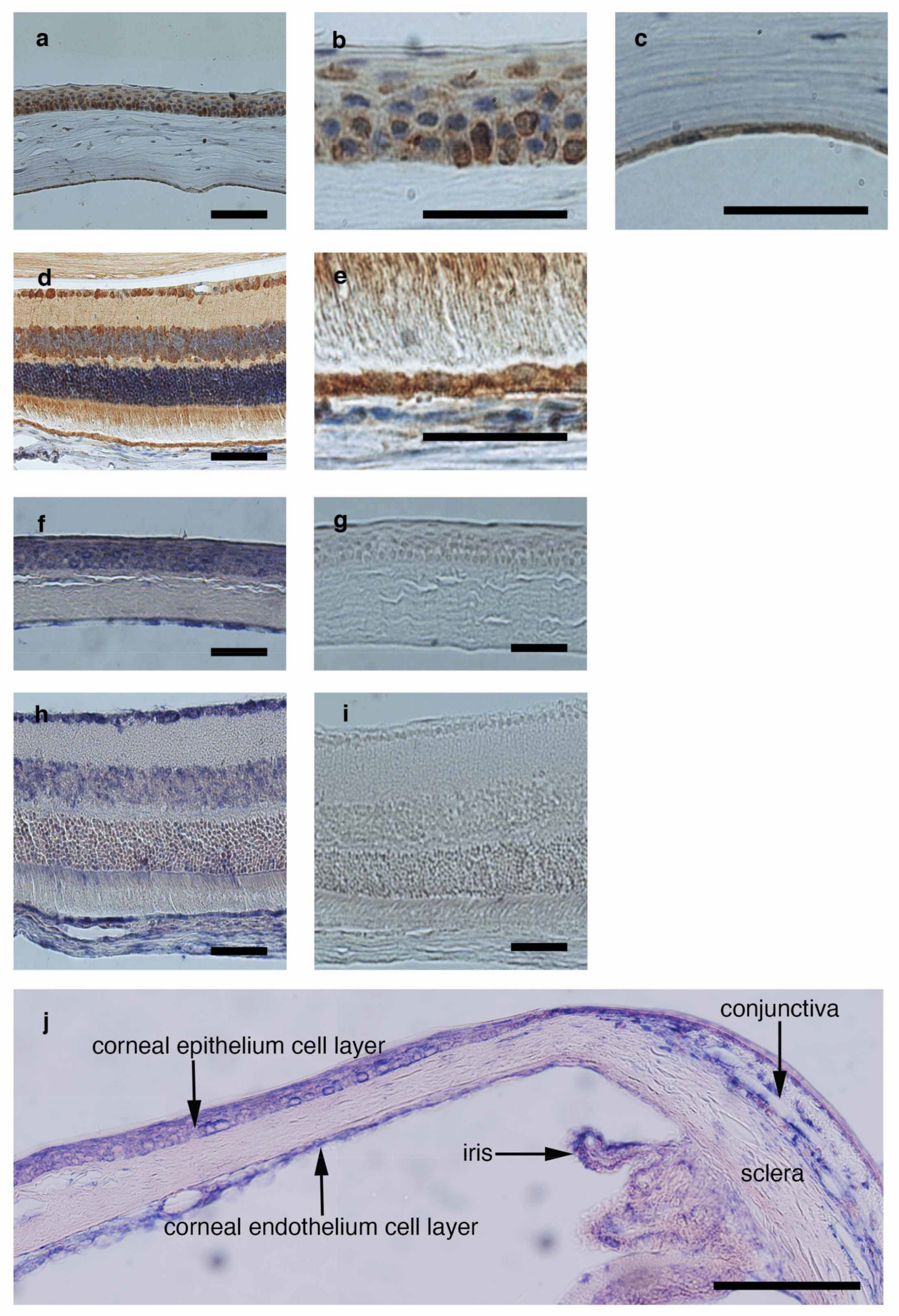
3.5. Detection of CTLA-2 Alpha Expression in the Ocular Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campochiaro, P.A. Retinal and Choroidal Neovascularization. J. Cell. Physiol. 2000, 184, 301–310. [Google Scholar] [CrossRef]
- Sekiyama, E.; Nakamura, T.; Cooper, L.J.; Kawasaki, S.; Hamuro, J.; Fullwood, N.J.; Kinoshita, S. Unique Distribution of thrombospondin-1 in Human Ocular Surface Epithelium. Investig. Ophthalmol. Vis. Sci. 2006, 47, 1352–1358. [Google Scholar] [CrossRef] [PubMed]
- Ambati, B.K.; Nozaki, M.; Singh, N.; Takeda, A.; Jani, P.D.; Suthar, T.; Albuquerque, R.J.; Richter, E.; Sakurai, E.; Newcomb, M.T.; et al. Corneal Avascularity Is Due to Soluble VEGF receptor-1. Nature 2006, 443, 993–997. [Google Scholar] [CrossRef] [Green Version]
- Cursiefen, C.; Seitz, B.; Dana, M.R.; Streilein, J.W. Angiogenesis and Lymphangiogenesis in the Cornea. Pathogenesis, Clinical Implications and Treatment Options. Ophthalmologe 2003, 100, 292–299. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, R.J.; Loughnan, M.S.; Flynn, E.; Folkman, J. Thalidomide Is an Inhibitor of Angiogenesis. Proc. Natl. Acad. Sci. USA 1994, 91, 4082–4085. [Google Scholar] [CrossRef] [Green Version]
- Cursiefen, C.; Chen, L.; Borges, L.P.; Jackson, D.; Cao, J.; Radziejewski, C.; D’Amore, P.A.; Dana, M.R.; Wiegand, S.J.; Streilein, J.W. VEGF-A Stimulates Lymphangiogenesis and Hemangiogenesis in Inflammatory Neovascularization via Macrophage Recruitment. J. Clin. Investig. 2004, 113, 1040–1050. [Google Scholar] [CrossRef] [Green Version]
- Cursiefen, C.; Cao, J.; Chen, L.; Liu, Y.; Maruyama, K.; Jackson, D.; Kruse, F.E.; Wiegand, S.J.; Dana, M.R.; Streilein, J.W. Inhibition of Hemangiogenesis and Lymphangiogenesis After Normal-Risk Corneal Transplantation by Neutralizing VEGF Promotes Graft Survival. Investig. Ophthalmol. Vis. Sci. 2004, 45, 2666–2673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalluri, R. Basement Membranes: Structure, Assembly and Role in Tumour Angiogenesis. Nat. Rev. Cancer 2003, 3, 422–433. [Google Scholar] [CrossRef]
- Akalu, A.; Roth, J.M.; Caunt, M.; Policarpio, D.; Liebes, L.; Brooks, P.C. Inhibition of Angiogenesis and Tumor Metastasis by Targeting a Matrix Immobilized Cryptic Extracellular Matrix Epitope in Laminin. Cancer Res. 2007, 67, 4353–4363. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.S.; Chang, J.H.; Yu, H.K.; Ahn, J.H.; Yum, J.S.; Lee, S.K.; Jung, K.H.; Park, D.H.; Yoon, Y.; Byun, S.M.; et al. Inhibition of Angiogenesis and Angiogenesis-Dependent Tumor Growth by the Cryptic Kringle Fragments of Human Apolipoprotein (a). J. Biol. Chem. 2003, 278, 29000–29008. [Google Scholar] [CrossRef] [Green Version]
- Colorado, P.C.; Torre, A.; Kamphaus, G.; Maeshima, Y.; Hopfer, H.; Takahashi, K.; Volk, R.; Zamborsky, E.D.; Herman, S.; Sarkar, P.K.; et al. Anti-Angiogenic Cues from Vascular Basement Membrane Collagen. Cancer Res. 2000, 60, 2520–2526. [Google Scholar]
- Rawlings, N.D.; Barrett, A.J. Families of Cysteine Peptidases. Methods Enzymol. 1994, 244, 461–486. [Google Scholar]
- Shaw, E.; Dean, R.T. The Inhibition of Macrophage Protein Turnover by a Selective Inhibitor of Thiol Proteinases. Biochem. J. 1980, 186, 385–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, A.J. Cellular Proteolysis. An Overview. Ann. N. Y. Acad. Sci. 1992, 674, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Kurata, M.; Watabe, S.; Murakami, R.; Takahashi, S.Y. Novel Cysteine Proteinase Inhibitors Homologous to the Proregions of Cysteine Proteinases. Curr. Protein Pept. Sci. 2002, 3, 231–238. [Google Scholar] [CrossRef]
- Joyce, J.A.; Baruch, A.; Chehade, K.; Meyer-Morse, N.; Giraudo, E.; Tsai, F.Y.; Greenbaum, D.C.; Hager, J.H.; Bogyo, M.; Hanahan, D. Cathepsin Cysteine Proteases Are Effectors of Invasive Growth and Angiogenesis During Multistage Tumorigenesis. Cancer Cell 2004, 5, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Berchem, G.; Glondu, M.; Gleizes, M.; Brouillet, J.P.; Vignon, F.; Garcia, M.; Liaudet-Coopman, E. Cathepsin-D Affects Multiple Tumor Progression Steps In Vivo: Proliferation, Angiogenesis and Apoptosis. Oncogene 2002, 21, 5951–5955. [Google Scholar] [CrossRef]
- Krueger, S.; Kellner, U.; Buehling, F.; Roessner, A. Cathepsin L Antisense Oligonucleotides in a Human Osteosarcoma Cell Line: Effects on the Invasive Phenotype. Cancer Gene Ther. 2001, 8, 522–528. [Google Scholar] [CrossRef] [Green Version]
- Urbich, C.; Heeschen, C.; Aicher, A.; Sasaki, K.; Bruhl, T.; Farhadi, M.R.; Vajkoczy, P.; Hofmann, W.K.; Peters, C.; Pennacchio, L.A.; et al. Cathepsin L Is Required for Endothelial Progenitor Cell-Induced Neovascularization. Nat. Med. 2005, 11, 206–213. [Google Scholar] [CrossRef] [Green Version]
- Kurata, M.; Hirata, M.; Watabe, S.; Miyake, M.; Takahashi, S.Y.; Yamamoto, Y. Expression, Purification, and Inhibitory Activities of Mouse Cytotoxic T-Lymphocyte antigen-2alpha. Protein Expr. Purif. 2003, 32, 119–125. [Google Scholar] [CrossRef]
- Maruyama, K.; Asai, J.; Ii, M.; Thorne, T.; Losordo, D.W.; D’Amore, P.A. Decreased Macrophage Number and Activation Lead to Reduced Lymphatic Vessel Formation and Contribute to Impaired Diabetic Wound Healing. Am. J. Pathol. 2007, 170, 1178–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobe, T.; Ortega, S.; Luna, J.D.; Ozaki, H.; Okamoto, N.; Derevjanik, N.L.; Vinores, S.A.; Basilico, C.; Campochiaro, P.A. Targeted Disruption of the FGF2 Gene Does Not Prevent Choroidal Neovascularization in a Murine Model. Am. J. Pathol. 1998, 153, 1641–1646. [Google Scholar] [CrossRef] [Green Version]
- Cursiefen, C.; Schlötzer-Schrehardt, U.; Küchle, M.; Sorokin, L.; Breiteneder-Geleff, S.; Alitalo, K.; Jackson, D. Lymphatic Vessels in Vascularized Human Corneas: Immunohistochemical Investigation Using LYVE-1 and Podoplanin. Investig. Ophthalmol. Vis. Sci. 2002, 43, 2127–2135. [Google Scholar]
- Chang, L.; Kaipainen, A.; Folkman, J. Lymphangiogenesis New Mechanisms. Ann. N. Y. Acad. Sci. 2002, 979, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Edelman, J.L.; Castro, M.R. Quantitative Image Analysis of Laser-Induced Choroidal Neovascularization in Rat. Exp. Eye Res. 2000, 71, 523–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, M.; Alard, P.; Streilein, J.W. TGF-Beta Promotes Immune Deviation by Altering Accessory Signals of Antigen-Presenting Cells. J. Immunol. 1998, 160, 1589–1597. [Google Scholar]
- Yi, H.; Nakamura, R.E.; Mohamed, O.; Dufort, D.; Hackam, A.S. Characterization of Wnt Signaling During Photoreceptor Degeneration. Investig. Ophthalmol. Vis. Sci. 2007, 48, 5733–5741. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, A.; Maruyama, K.; Walsh, T.; Kajiya, K.; Hamuro, J.; D’Amore, P.A.; Kinoshita, S. Intracellular Thiol Redox Status Regulates Lymphangiogenesis and Dictates Corneal Limbal Graft Survival. Investig. Ophthalmol. Vis. Sci. 2010, 51, 2450–2458. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, K.; Ii, M.; Cursiefen, C.; Jackson, D.G.; Keino, H.; Tomita, M.; Van Rooijen, N.; Takenaka, H.; D’Amore, P.A.; Stein-Streilein, J.; et al. Inflammation-Induced Lymphangiogenesis in the Cornea Arises from CD11b-Positive Macrophages. J. Clin. Investig. 2005, 115, 2363–2372. [Google Scholar] [CrossRef] [PubMed]
- Sugita, S.; Horie, S.; Nakamura, O.; Futagami, Y.; Takase, H.; Keino, H.; Aburatani, H.; Katunuma, N.; Ishidoh, K.; Yamamoto, Y.; et al. Retinal Pigment Epithelium-Derived CTLA-2alpha Induces TGFbeta-Producing T Regulatory Cells. J. Immunol. 2008, 181, 7525–7536. [Google Scholar] [CrossRef] [Green Version]
- Shimada, N.; Ohno-Matsui, K.; Iseki, S.; Koike, M.; Uchiyama, Y.; Wang, J.; Yoshida, T.; Sato, T.; Peters, C.; Mochizuki, M.; et al. Cathepsin L in Bone Marrow-Derived Cells Is Required for Retinal and Choroidal Neovascularization. Am. J. Pathol. 2010, 176, 2571–2580. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, T.; Hata, Y.; Yoshikawa, H.; Nakagawa, K.; Sueishi, K.; Inomata, H. Expression of Vascular Endothelial Growth Factor in Experimental Choroidal Neovascularization. Graefes Arch. Clin. Exp. Ophthalmol. 1997, 235, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Nakao, S.; Kuwano, T.; Tsutsumi-Miyahara, C.; Ueda, S.; Kimura, Y.N.; Hamano, S.; Sonoda, K.H.; Saijo, Y.; Nukiwa, T.; Strieter, R.M.; et al. Infiltration of COX-2-Expressing Macrophages Is a Prerequisite for IL-1 Beta-Induced Neovascularization and Tumor Growth. J. Clin. Investig. 2005, 115, 2979–2991. [Google Scholar] [CrossRef] [PubMed]
- Streilein, J.W.; Masli, S.; Takeuchi, M.; Kezuka, T. The Eye’s View of Antigen Presentation. Hum. Immunol. 2002, 63, 435–443. [Google Scholar] [CrossRef]
- Masli, S.; Turpie, B.; Hecker, K.H.; Streilein, J.W. Expression of Thrombospondin in TGFbeta-Treated APCs and Its Relevance to Their Immune Deviation-Promoting Properties. J. Immunol. 2002, 168, 2264–2273. [Google Scholar] [CrossRef] [Green Version]
- Murata, Y.; Shimamura, T.; Hamuro, J. The Polarization of T(h)1/T(h)2 Balance Is Dependent on the Intracellular Thiol Redox Status of Macrophages due to the Distinctive Cytokine Production. Int. Immunol. 2002, 14, 201–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.K.; Llewellyn, O.P.; Bates, D.O.; Nicholson, L.B.; Dick, A.D. IL-10 Regulation of Macrophage VEGF Production Is Dependent on Macrophage Polarisation and Hypoxia. Immunobiology 2010, 215, 796–803. [Google Scholar] [CrossRef] [PubMed]
- Roth, W.; Deussing, J.; Botchkarev, V.A.; Pauly-Evers, M.; Saftig, P.; Hafner, A.; Schmidt, P.; Schmahl, W.; Scherer, J.; Anton-Lamprecht, I.; et al. Cathepsin L Deficiency as Molecular Defect of Furless: Hyperproliferation of Keratinocytes and Pertubation of Hair Follicle Cycling. FASEB J. 2000, 14, 2075–2086. [Google Scholar] [CrossRef]
- Rebbaa, A.; Chu, F.; Sudha, T.; Gallati, C.; Dier, U.; Dyskin, E.; Yalcin, M.; Bianchini, C.; Shaker, O.; Mousa, S.A. The Anti-Angiogenic Activity of NSITC, a Specific Cathepsin L Inhibitor. Anticancer Res. 2009, 29, 4473–4481. [Google Scholar]
- Futagami, Y.; Sugita, S.; Vega, J.; Ishida, K.; Takase, H.; Maruyama, K.; Aburatani, H.; Mochizuki, M. Role of thrombospondin-1 in T Cell Response to Ocular Pigment Epithelial Cells. J. Immunol. 2007, 178, 6994–7005. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maruyama, K.; Yoneda, K.; Sugita, S.; Yamamoto, Y.; Koike, M.; Peters, C.; Uchiyama, Y.; Nishida, K. CTLA-2 Alpha Is a Potent Inhibitor of Angiogenesis in Murine Ocular Tissue. Antioxidants 2021, 10, 456. https://doi.org/10.3390/antiox10030456
Maruyama K, Yoneda K, Sugita S, Yamamoto Y, Koike M, Peters C, Uchiyama Y, Nishida K. CTLA-2 Alpha Is a Potent Inhibitor of Angiogenesis in Murine Ocular Tissue. Antioxidants. 2021; 10(3):456. https://doi.org/10.3390/antiox10030456
Chicago/Turabian StyleMaruyama, Kazuichi, Kazuhito Yoneda, Sunao Sugita, Yoshimi Yamamoto, Masato Koike, Christoph Peters, Yasuo Uchiyama, and Kohji Nishida. 2021. "CTLA-2 Alpha Is a Potent Inhibitor of Angiogenesis in Murine Ocular Tissue" Antioxidants 10, no. 3: 456. https://doi.org/10.3390/antiox10030456