Cigarette Smoke Extract Activates Hypoxia-Inducible Factors in a Reactive Oxygen Species-Dependent Manner in Stroma Cells from Human Endometrium

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Tissue Collection
2.3. Cell Culture
2.4. Preparation of CSE
2.5. Cell Proliferation Assay
2.6. Western Blot Analysis
2.7. ROS Assay
2.8. Luciferase Assay
2.9. Semi-Quantitative Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.10. RNA-Seq
2.11. Transcriptomics Analysis
2.12. Statistics
3. Results
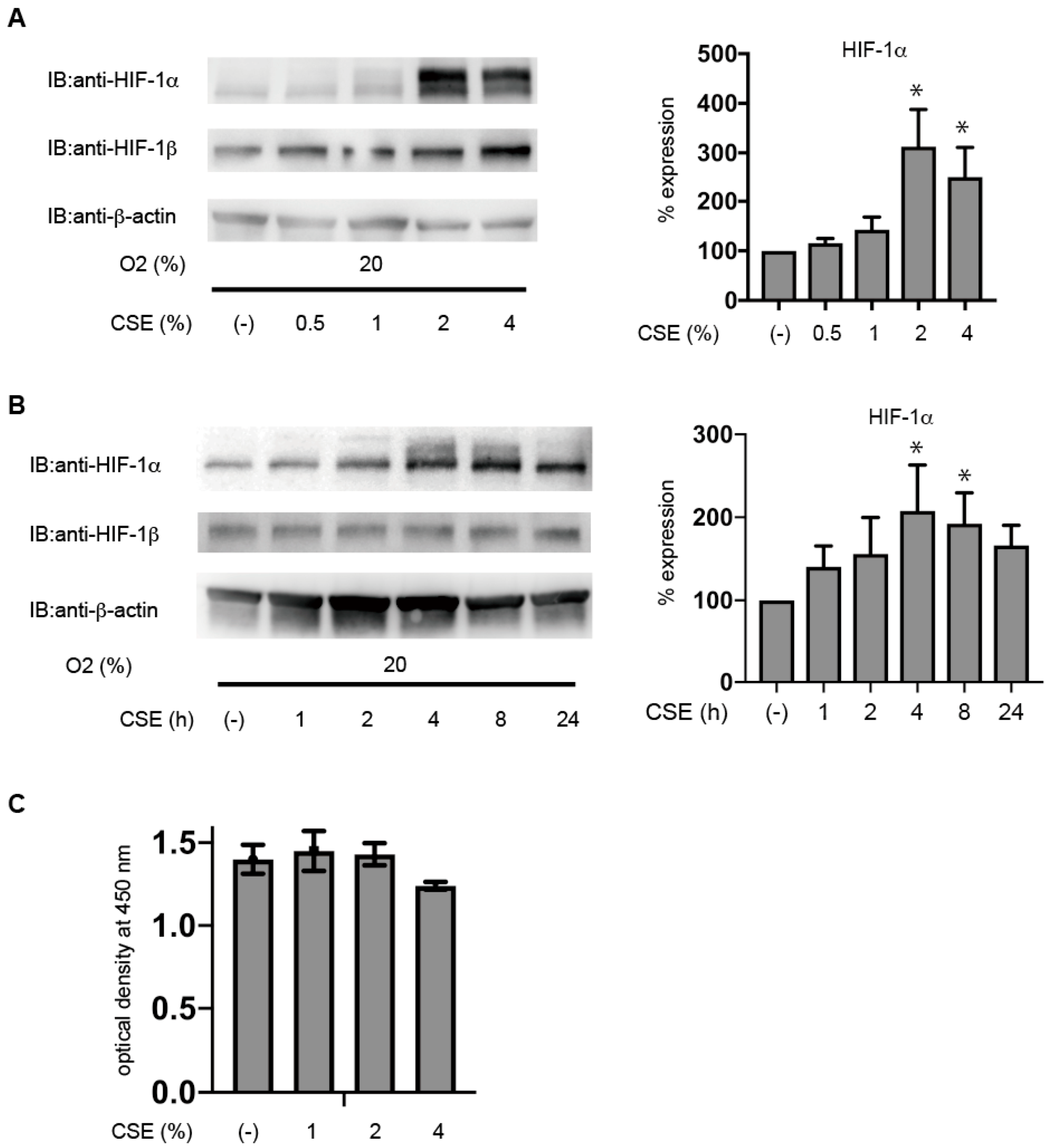
3.1. CSE Induces HIF-1α Protein Accumulation under Non-Hypoxic Conditions in Primary Cultured Human ESCs
3.2. CSE Induces HIF-1α Protein Accumulation under Non-Hypoxic Conditions in Immortalized KC02-44D Cells
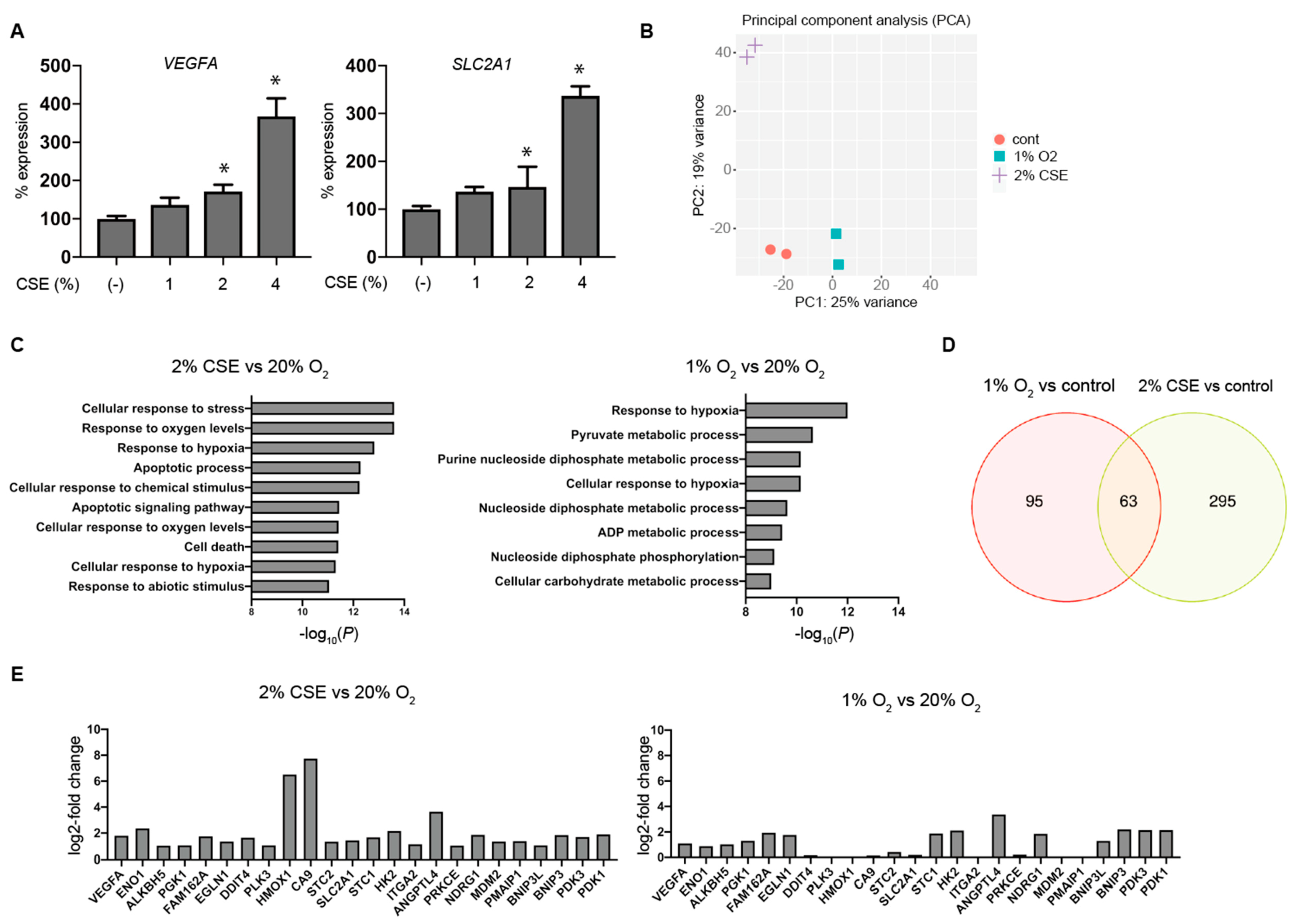
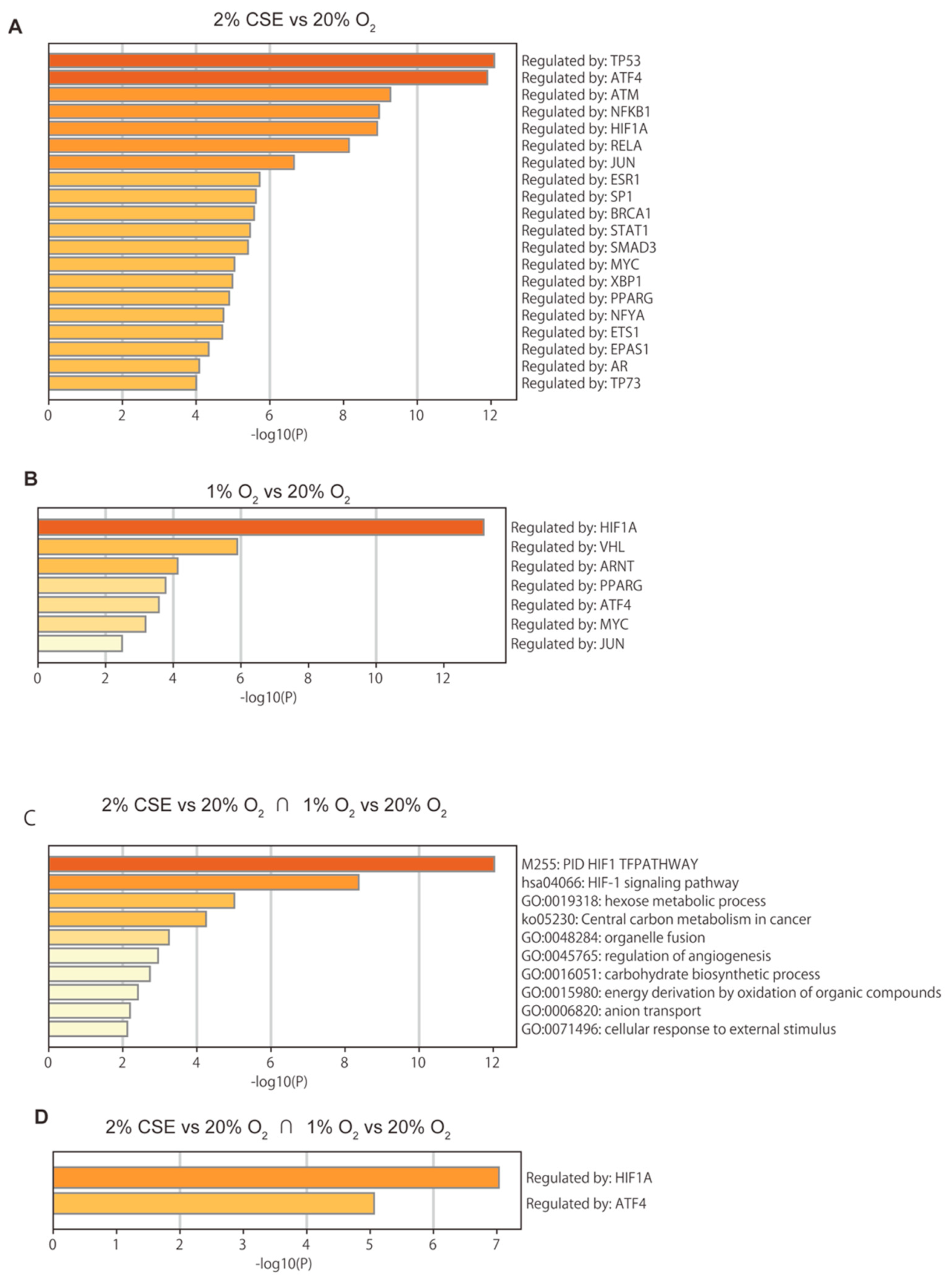
3.3. Global Gene-Expression Analysis Using RNA-Seq and Next-Generation Sequencing (NGS)
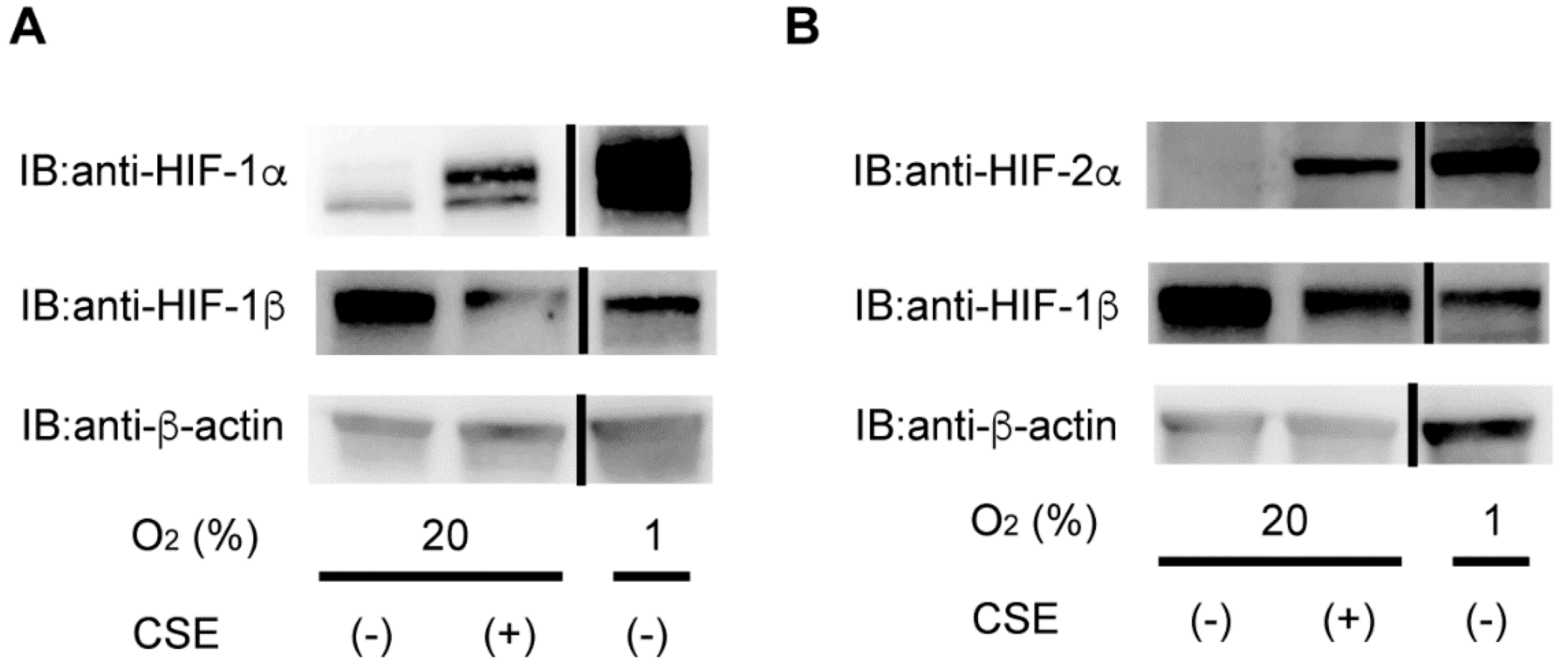
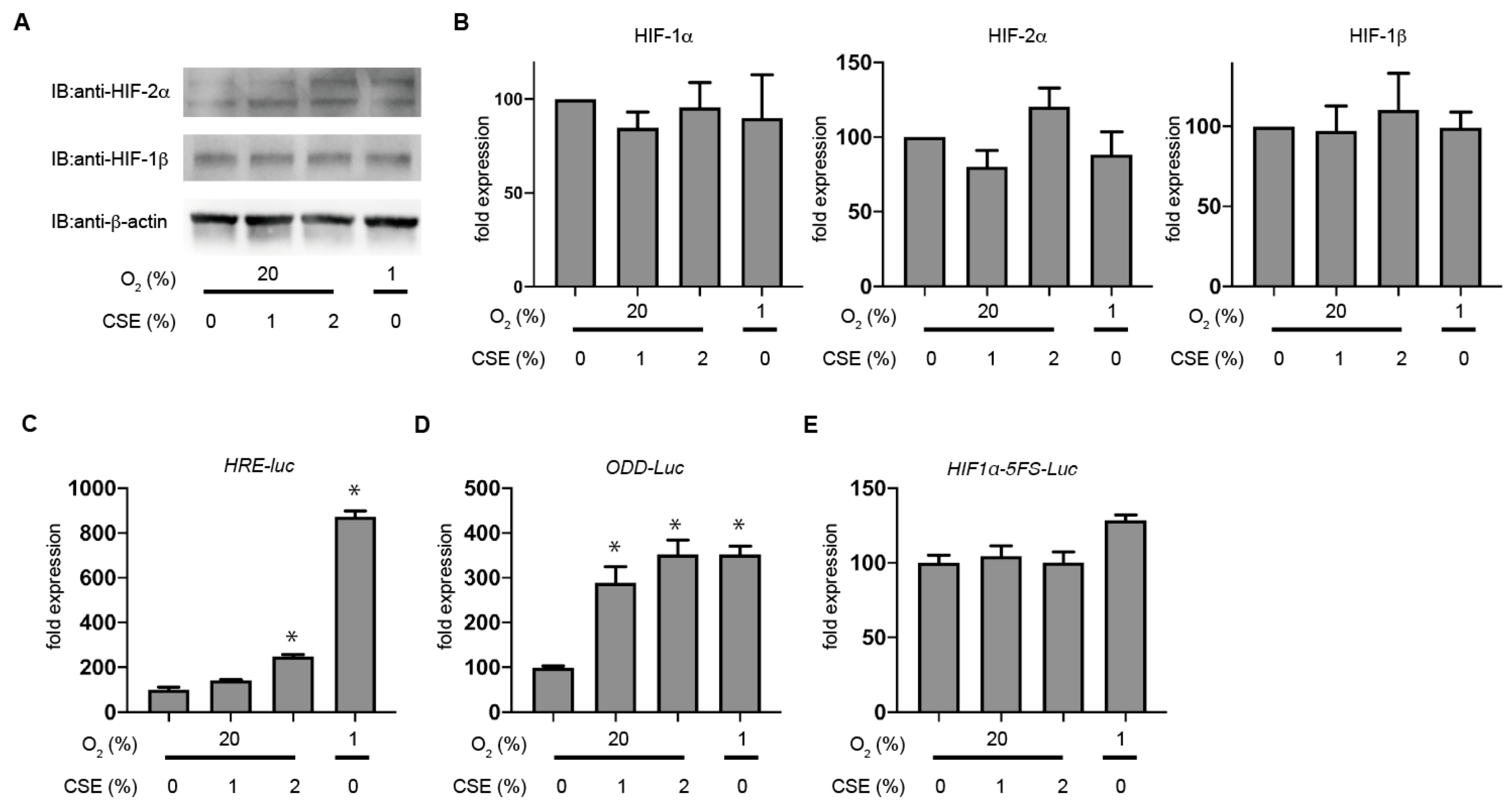
3.4. CSE Selectively Stabilizes HIF-1α Levels rather than HIF-2α
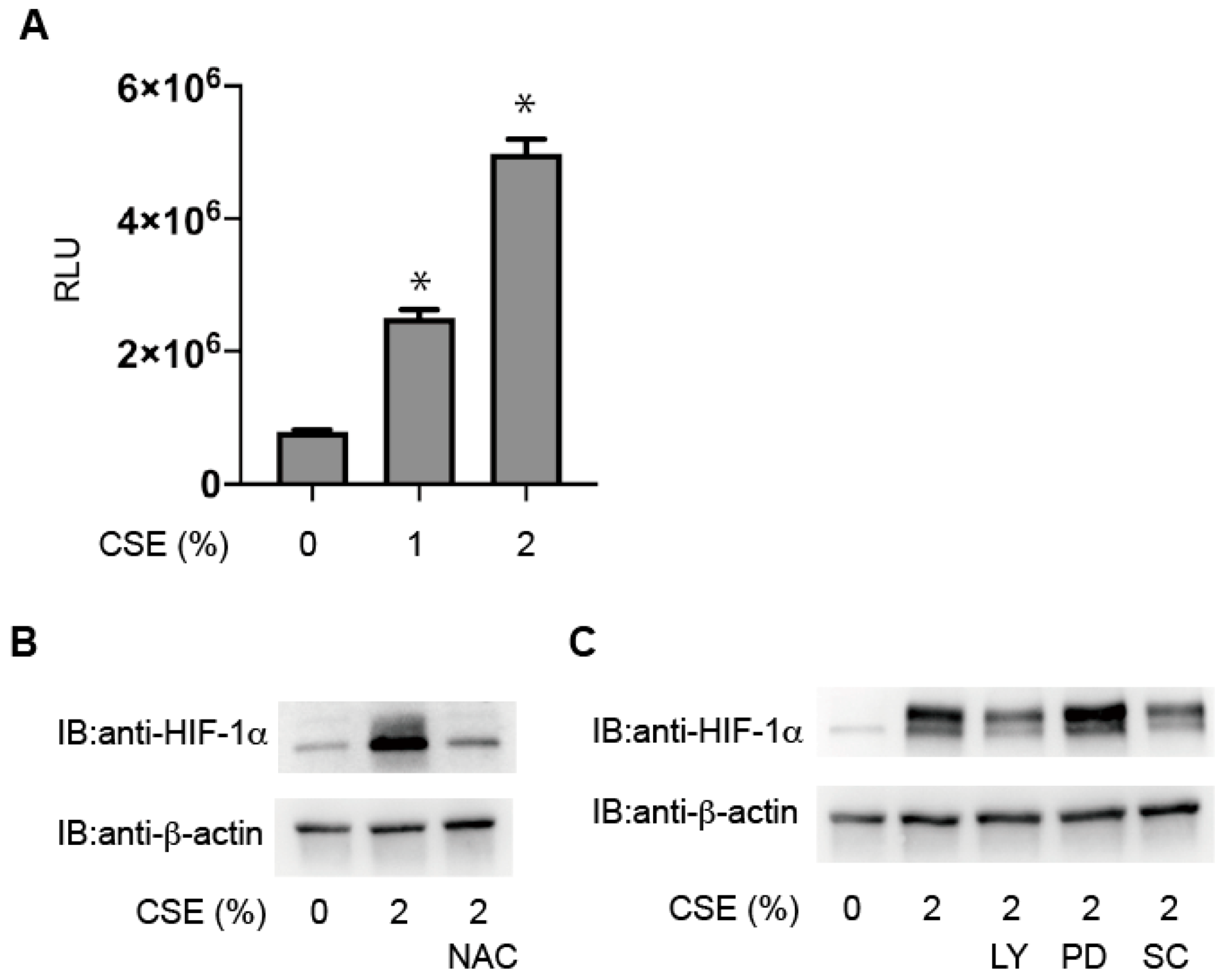
3.5. Critical Involvement of ROS in HIF-1 Activation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CSE | cigarette smoke extract |
| HIF | hypoxia-inducible factor |
| ESC | endometrial stromal cell |
| ROS | reactive oxygen species |
| GSEA | gene set enrichment analysis |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagents | Source | Identifier |
|---|---|---|
| 2-Amino-2-hydroxymethyl-1,3-propanediol | Wako | 207-06275 |
| 7.5% Mini-PROTEAN® TGXTM Precast Protein Gels | BIO-RAD | 4561023 |
| Anti-rabbit IgG (H + L), F(ab’)2 Fragment (Alexa Fluor® 488 Conjugate) | Cell Signaling TECHNOLOGY | 4412 |
| Anti-β-Actin antibody | Sigma-Aldrich | A5316 |
| Antibiotic-Antimycotic (100X) | Gibco | 5240-062 |
| Blocking One | Nacalai Tesque | 03953-95 |
| Bovine serum albumin | Wako | 017-23294 |
| Cell Counting Kit-8 | Dojindo | CK04 |
| DEXTRAN | Sigma-Aldrich | D4751 |
| Dithiothreitol (DTT) | BIO-RAD | 161-0610 |
| DMEM/F-12, HEPES, no phenol red | Gibco | 11039-021 |
| DMEM/F12, Phenol Red | Gibco | 11330-032 |
| ECLTM Prime Western Blotting Detection Reagents | GE Healthcare | RPN2232 |
| Ethanol (99.5) | Wako | 057-00456 |
| fetal bovine serum | Sigma-Aldrich | 172012 |
| Formaldehyde | Polysciences | 18814 |
| FuGENE® HD Transfection Reagent | Promega | E2311 |
| GlutaMAX™ Supplement | Gibco | 35050-061 |
| Glycine | Wako | 077-00735 |
| HIF-1β/ARNT(D28F3)XP rabbit monoclonal antibody | Cell Signaling TECHNOLOGY | 5537 |
| HIF-2 alpha/EPAS1 Antibody | Novus Biologicals | NB100-122 |
| Hydrochloric Acid | Wako | 084-05425 |
| LY294002 | Wako | 129-04861 |
| N-Acetyl-L-cysteine, ROS inhibitor | abcam | 143032 |
| M-PER™ Mammalian Protein Extraction Reagent | Thermo Fisher Scientific | 78501 |
| Mouse IgG HRP Linked Whole Ab | GE Healthcare | NA931 |
| normal rabbit IgG | SANTA CRUZ BIOTECHNOLOGY | sc-2027 |
| Opti-MEM™ I Reduced Serum Medium | Gibco | 31985-062 |
| ONE-Glo™ Luciferase Assay System | Promega | E6120 |
| PBS, pH 7.4 | Gibco | 10010-023 |
| PD98059 | Wako | 169-19211 |
| Pierce™ BCA Protein Assay Kit | Thermo Fisher Scientific | 23225 |
| Polyoxyethylene(20) Sorbitan Monolaurate | Wako | 167-11515 |
| Progesterone receptor Antibody | SANTA CRUZ BIOTECHNOLOGY | sc-166169 |
| Protease inhibitor cocktail | Calbiochem | 539134 |
| Purified Mouse Anti-Human HIF-1α | BD Transduction LaboratoriesTM | 610958 |
| ReverTra Ace® qPCR RT Master Mix with gDNA Remover | TOYOBO | FSQ-301 |
| RNeasy Mini Kit | Qiagen | 74106 |
| ROS-Glo™ H2O2 Assay | Promega | G8820 |
| Running Buffer Solution fro SDS-PAGE | Nacalai Tesque | 30329-61 |
| Tris Bufferd Saline with Tween® 20 (TBS-T) Tablets, pH 7.6 | TaKaRa | 79142 |
| THUNDERBIRD® SYBR qPCR Mix | TOYOBO | QPS-201 |
| Trans-Blot® Turbo™ Mini PVDF Transfer Packs | BIO-RAD | 1704156 |
| Trypsin-EDTA (0.25%) | Gibco | 25200-056 |
| SC-514 | Sigma | SML0557-5MG |
| X-tremeGENE HP DNA Transfection Reagent | Roche | 6366236001 |
| β-Mercaptoethanol | Wako | 131-14572 |
References
- Jha, P.; Peto, R. Global effects of smoking, of quitting, and of taxing tobacco. N. Engl. J. Med. 2014, 370, 60–68. [Google Scholar] [CrossRef] [Green Version]
- Shiverick, K.T.; Salafia, C. Cigarette smoking and pregnancy I: Ovarian, uterine and placental effects. Placenta 1999, 20, 265–272. [Google Scholar] [CrossRef]
- Orzabal, M.R.; Lunde-Young, E.R.; Ramirez, J.I.; Howe, S.Y.F.; Naik, V.D.; Lee, J.; Heaps, C.L.; Threadgill, D.W.; Ramadoss, J. Chronic exposure to e-cig aerosols during early development causes vascular dysfunction and offspring growth deficits. Transl. Res. 2019, 207, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, Y.; Tsuji, I.; Mizoue, T.; Inoue, M.; Sawada, N.; Matsuo, K.; Ito, H.; Naito, M.; Nagata, C.; Kitamura, Y.; et al. Cigarette smoking and cervical cancer risk: An evaluation based on a systematic review and meta-analysis among Japanese women. Jpn. J. Clin. Oncol. 2019, 49, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Jorgensen, E.M.; Gan, Y.; Taylor, H.S. Cigarette smoke increases progesterone receptor and homeobox A10 expression in human endometrium and endometrial cells: A potential role in the decreased prevalence of endometrial pathology in smokers. Biol. Reprod. 2011, 84, 1242–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamoto, T.; Yasuda, K.; Yasuhara, M.; Nakajima, T.; Mizokami, T.; Okada, H.; Kanzaki, H. Cigarette smoke extract enhances oxytocin-induced rhythmic contractions of rat and human preterm myometrium. Reproduction 2006, 132, 343–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, S.R.; Simon, C.; Remohi, J.; Pellicer, A. Cigarette smoking affects uterine receptiveness. Hum. Reprod. 2007, 22, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi, R.; Hiroi, H.; Momoeda, M.; Hosokawa, Y.; Nakazawa, F.; Yano, T.; Tsutsumi, O.; Taketani, Y. Induction of early decidualization by cadmium, a major contaminant of cigarette smoke. Fertil. Steril. 2009, 91, 1614–1617. [Google Scholar] [CrossRef] [PubMed]
- Dechanet, C.; Anahory, T.; Mathieu Daude, J.C.; Quantin, X.; Reyftmann, L.; Hamamah, S.; Hedon, B.; Dechaud, H. Effects of cigarette smoking on reproduction. Hum. Reprod. Update 2011, 17, 76–95. [Google Scholar] [CrossRef] [Green Version]
- Esposito, E.R.; Horn, K.H.; Greene, R.M.; Pisano, M.M. An animal model of cigarette smoke-induced in utero growth retardation. Toxicology 2008, 246, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Salafia, C.; Shiverick, K. Cigarette smoking and pregnancy II: Vascular effects. Placenta 1999, 20, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Rodesch, F.; Simon, P.; Donner, C.; Jauniaux, E. Oxygen measurements in endometrial and trophoblastic tissues during early pregnancy. Obstet. Gynecol. 1992, 80, 283–285. [Google Scholar] [PubMed]
- Gassmann, M.; Fandrey, J.; Bichet, S.; Wartenberg, M.; Marti, H.H.; Bauer, C.; Wenger, R.H.; Acker, H. Oxygen supply and oxygen-dependent gene expression in differentiating embryonic stem cells. Proc. Natl. Acad. Sci. USA 1996, 93, 2867–2872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirota, K. Basic Biology of Hypoxic Responses Mediated by the Transcription Factor HIFs and Its Implication for Medicine. Biomedicines 2020, 8, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daikoku, T.; Matsumoto, H.; Gupta, R.A.; Das, S.K.; Gassmann, M.; DuBois, R.N.; Dey, S.K. Expression of hypoxia-inducible factors in the peri-implantation mouse uterus is regulated in a cell-specific and ovarian steroid hormone-dependent manner. Evidence for differential function of HIFs during early pregnancy. J. Biol. Chem. 2003, 278, 7683–7691. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, L.; Hirota, Y.; Saito-Fujita, T.; Takeda, N.; Tanaka, T.; Hiraoka, T.; Akaeda, S.; Fujita, H.; Shimizu-Hirota, R.; Igaue, S.; et al. HIF2alpha in the uterine stroma permits embryo invasion and luminal epithelium detachment. J. Clin. Investig. 2018, 128, 3186–3197. [Google Scholar] [CrossRef] [Green Version]
- Majmundar, A.J.; Wong, W.J.; Simon, M.C. Hypoxia-inducible factors and the response to hypoxic stress. Mol. Cell 2010, 40, 294–309. [Google Scholar] [CrossRef] [Green Version]
- Ryan, H.E.; Lo, J.; Johnson, R.S. HIF-1 alpha is required for solid tumor formation and embryonic vascularization. EMBO J. 1998, 17, 3005–3015. [Google Scholar] [CrossRef] [Green Version]
- McGettrick, A.F.; O’Neill, L.A.J. The Role of HIF in Immunity and Inflammation. Cell Metab. 2020, 32, 524–536. [Google Scholar] [CrossRef]
- Daijo, H.; Hoshino, Y.; Kai, S.; Suzuki, K.; Nishi, K.; Matsuo, Y.; Harada, H.; Hirota, K. Cigarette smoke reversibly activates hypoxia-inducible factor 1 in a reactive oxygen species-dependent manner. Sci. Rep. 2016, 6, 34424. [Google Scholar] [CrossRef] [Green Version]
- Tsuzuki, T.; Okada, H.; Cho, H.; Shimoi, K.; Miyashiro, H.; Yasuda, K.; Kanzaki, H. Divergent regulation of angiopoietin-1, angiopoietin-2, and vascular endothelial growth factor by hypoxia and female sex steroids in human endometrial stromal cells. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 168, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Tsuzuki, T.; Okada, H.; Cho, H.; Tsuji, S.; Nishigaki, A.; Yasuda, K.; Kanzaki, H. Hypoxic stress simultaneously stimulates vascular endothelial growth factor via hypoxia-inducible factor-1alpha and inhibits stromal cell-derived factor-1 in human endometrial stromal cells. Hum. Reprod. 2012, 27, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Okada, H.; Nakajima, T.; Sanezumi, M.; Ikuta, A.; Yasuda, K.; Kanzaki, H. Progesterone enhances interleukin-15 production in human endometrial stromal cells in vitro. J. Clin. Endocrinol. Metab. 2000, 85, 4765–4770. [Google Scholar] [CrossRef] [PubMed]
- Yuhki, M.; Kajitani, T.; Mizuno, T.; Aoki, Y.; Maruyama, T. Establishment of an immortalized human endometrial stromal cell line with functional responses to ovarian stimuli. Reprod. Biol. Endocrinol. 2011, 9, 104. [Google Scholar] [CrossRef] [Green Version]
- Rius, C.; Company, C.; Piqueras, L.; Cerda-Nicolas, J.M.; Gonzalez, C.; Servera, E.; Ludwig, A.; Morcillo, E.J.; Sanz, M.J. Critical role of fractalkine (CX3CL1) in cigarette smoke-induced mononuclear cell adhesion to the arterial endothelium. Thorax 2013, 68, 177–186. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Han, W.; Giraldo, C.; De Li, Y.; Block, E.R. Effect of cigarette smoke extract on nitric oxide synthase in pulmonary artery endothelial cells. Am. J. Respir. Cell Mol. Biol. 1998, 19, 819–825. [Google Scholar] [CrossRef] [Green Version]
- Wakamatsu, T.; Tanaka, T.; Oda, S.; Nishi, K.; Harada, H.; Daijo, H.; Takabuchi, S.; Kai, S.; Fukuda, K.; Hirota, K. The intravenous anesthetics barbiturates inhibit hypoxia-inducible factor 1 activation. Eur. J. Pharmacol. 2009, 617, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Morinibu, A.; Koyasu, S.; Goto, Y.; Hiraoka, M.; Harada, H. A circadian clock gene, PER2, activates HIF-1 as an effector molecule for recruitment of HIF-1alpha to promoter regions of its downstream genes. FEBS J. 2017, 284, 3804–3816. [Google Scholar] [CrossRef] [Green Version]
- Zeng, L.; Morinibu, A.; Kobayashi, M.; Zhu, Y.; Wang, X.; Goto, Y.; Yeom, C.J.; Zhao, T.; Hirota, K.; Shinomiya, K.; et al. Aberrant IDH3alpha expression promotes malignant tumor growth by inducing HIF-1-mediated metabolic reprogramming and angiogenesis. Oncogene 2015, 34, 4758–4766. [Google Scholar] [CrossRef] [Green Version]
- Kakita-Kobayashi, M.; Murata, H.; Nishigaki, A.; Hashimoto, Y.; Komiya, S.; Tsubokura, H.; Kido, T.; Kida, N.; Tsuzuki-Nakao, T.; Matsuo, Y.; et al. Thyroid Hormone Facilitates in vitro Decidualization of Human Endometrial Stromal Cells via Thyroid Hormone Receptors. Endocrinology 2020, 161, bqaa049. [Google Scholar] [CrossRef]
- Sumi, C.; Matsuo, Y.; Kusunoki, M.; Shoji, T.; Uba, T.; Iwai, T.; Bono, H.; Hirota, K. Cancerous phenotypes associated with hypoxia-inducible factors are not influenced by the volatile anesthetic isoflurane in renal cell carcinoma. PLoS ONE 2019, 14, e0215072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiraoka, Y.; Yamada, K.; Kawasaki, Y.; Hirose, H.; Matsumoto, K.; Ishikawa, K.; Yasumizu, Y. Ikra: RNAseq pipeline centered on Salmon. Zenodo 2019. [Google Scholar] [CrossRef]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krueger, F. Trim Galore. Available online: https://www.bioinformatics.babraham.ac.uk/projects/trim_galore/ (accessed on 14 October 2020).
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Ge, S.X.; Son, E.W.; Yao, R. iDEP: An integrated web application for differential expression and pathway analysis of RNA-Seq data. BMC Bioinform. 2018, 19, 534. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Han, H.; Cho, J.W.; Lee, S.; Yun, A.; Kim, H.; Bae, D.; Yang, S.; Kim, C.Y.; Lee, M.; Kim, E.; et al. TRRUST v2: An expanded reference database of human and mouse transcriptional regulatory interactions. Nucleic Acids Res. 2018, 46, D380–D386. [Google Scholar] [CrossRef]
- Han, H.; Shim, H.; Shin, D.; Shim, J.E.; Ko, Y.; Shin, J.; Kim, H.; Cho, A.; Kim, E.; Lee, T.; et al. TRRUST: A reference database of human transcriptional regulatory interactions. Sci. Rep. 2015, 5, 11432. [Google Scholar] [CrossRef]
- Kim, S.M.; Hwang, K.A.; Go, R.E.; Sung, J.H.; Choi, D.W.; Choi, K.C. Exposure to cigarette smoke via respiratory system may induce abnormal alterations of reproductive organs in female diabetic rats. Environ. Toxicol. 2019, 34, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Chandel, N.S. There Is No Smoke without Mitochondria. Am. J. Respir. Cell Mol. Biol. 2019, 60, 489–491. [Google Scholar] [CrossRef]
- Jaimes, E.A.; DeMaster, E.G.; Tian, R.X.; Raij, L. Stable compounds of cigarette smoke induce endothelial superoxide anion production via NADPH oxidase activation. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1031–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Z.; Han, Z.; Li, Z.; Hu, H.; Patel, J.M.; Antony, V.; Block, E.R.; Su, Y. Involvement of calpain-calpastatin in cigarette smoke-induced inhibition of lung endothelial nitric oxide synthase. Am. J. Respir. Cell Mol. Biol. 2005, 33, 513–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, S.; Das, H.; Das, U.; Sengupta, A.; Dey Sharma, R.; Biswas, S.C.; Dey, S. Smokeless tobacco induces toxicity and apoptosis in neuronal cells: A mechanistic evaluation. Free Radic. Res. 2020, 54, 477–496. [Google Scholar] [CrossRef] [PubMed]
- Perez-Rial, S.; Barreiro, E.; Fernandez-Acenero, M.J.; Fernandez-Valle, M.E.; Gonzalez-Mangado, N.; Peces-Barba, G. Early detection of skeletal muscle bioenergetic deficit by magnetic resonance spectroscopy in cigarette smoke-exposed mice. PLoS ONE 2020, 15, e0234606. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L.; Prabhakar, N.R. The role of hypoxia-inducible factors in oxygen sensing by the carotid body. Adv. Exp. Med. Biol. 2012, 758, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenza, G.L.; Prabhakar, N.R. The role of hypoxia-inducible factors in carotid body (patho) physiology. J. Physiol. 2018, 596, 2977–2983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maybin, J.A.; Murray, A.A.; Saunders, P.T.K.; Hirani, N.; Carmeliet, P.; Critchley, H.O.D. Hypoxia and hypoxia inducible factor-1alpha are required for normal endometrial repair during menstruation. Nat. Commun. 2018, 9, 295. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G.L. Life with oxygen. Science 2007, 318, 62–64. [Google Scholar] [CrossRef]
- Semenza, G.L. HIF-1 and human disease: One highly involved factor. Genes Dev. 2000, 14, 1983–1991. [Google Scholar]
- Zhan, L.; Wang, W.; Zhang, Y.; Song, E.; Fan, Y.; Wei, B. Hypoxia-inducible factor-1alpha: A promising therapeutic target in endometriosis. Biochimie 2016, 123, 130–137. [Google Scholar] [CrossRef]
- Sava, R.I.; March, K.L.; Pepine, C.J. Hypertension in pregnancy: Taking cues from pathophysiology for clinical practice. Clin. Cardiol. 2018, 41, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.; Landers, K.; Mortimer, R.H.; Richard, K. Regulation of hypoxia inducible factors (HIF) in hypoxia and normoxia during placental development. Placenta 2010, 31, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.M.; Choi, K.C. Cigarette smoke extract and isoprene resulted in the induction of apoptosis and autophagy in human placenta choriocarcinoma JEG-3 cells. Environ. Toxicol. 2018, 33, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.D.; Lee, D.S.; Chun, Y.G.; Shim, T.S.; Lim, C.M.; Koh, Y.; Kim, W.S.; Kim, D.S.; Kim, W.D. Cigarette smoke extract induces endothelin-1 via protein kinase C in pulmonary artery endothelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2001, 281, L403–L411. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.M.; Kim, C.W.; Hwang, K.A.; Sung, J.H.; Lee, J.K.; Choi, K.C. Cigarette smoke impaired maturation of ovarian follicles and normal growth of uterus inner wall of female wild-type and hypertensive rats. Reprod. Toxicol. 2017, 73, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Bernhard, D.; Wang, X.L. Smoking, oxidative stress and cardiovascular diseases—Do anti-oxidative therapies fail? Curr. Med. Chem. 2007, 14, 1703–1712. [Google Scholar] [CrossRef]






| Target | Gene Symbol | Forward Primer 5′–3′ | Reverse Primer 5′–3′ |
|---|---|---|---|
| VEGF | VEGFA | CGAAACCATGAACTTTCTGC | CCTCAGTGGGCACACACTCC |
| GLUT1 | SLC2A1 | TCCACGAGCATCTTCGAGA | ATACTGGAAGCACATGCCC |
| HIF-1a | HIF1A | ACACACAGAAATGGCCTTGTGA | CCTGTGCAGTGCAATACCTTC |
| HIF-2a | EPAS1 | ATGGGACTTACACAGGTGGAG | GCTCTGTGGACATGTCTTTGC |
| HIF-1b | ARNT | TGTTGGCTACCAGCCACAGGAACT | ACCGGAACCGGAACATGACAGA |
| EF1α | EEF1A1 | TCTGGTTGGAATGGTGACAACATGC | AGAGCTTCACTCAAAGCTTCATGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kida, N.; Matsuo, Y.; Hashimoto, Y.; Nishi, K.; Tsuzuki-Nakao, T.; Bono, H.; Maruyama, T.; Hirota, K.; Okada, H. Cigarette Smoke Extract Activates Hypoxia-Inducible Factors in a Reactive Oxygen Species-Dependent Manner in Stroma Cells from Human Endometrium. Antioxidants 2021, 10, 48. https://doi.org/10.3390/antiox10010048
Kida N, Matsuo Y, Hashimoto Y, Nishi K, Tsuzuki-Nakao T, Bono H, Maruyama T, Hirota K, Okada H. Cigarette Smoke Extract Activates Hypoxia-Inducible Factors in a Reactive Oxygen Species-Dependent Manner in Stroma Cells from Human Endometrium. Antioxidants. 2021; 10(1):48. https://doi.org/10.3390/antiox10010048
Chicago/Turabian StyleKida, Naoko, Yoshiyuki Matsuo, Yoshiko Hashimoto, Kenichiro Nishi, Tomoko Tsuzuki-Nakao, Hidemasa Bono, Tetsuo Maruyama, Kiichi Hirota, and Hidetaka Okada. 2021. "Cigarette Smoke Extract Activates Hypoxia-Inducible Factors in a Reactive Oxygen Species-Dependent Manner in Stroma Cells from Human Endometrium" Antioxidants 10, no. 1: 48. https://doi.org/10.3390/antiox10010048