Bactericidal Activity of Lactic Acid against Clinical, Carbapenem-Hydrolyzing, Multi-Drug-Resistant Klebsiella pneumoniae Planktonic and Biofilm-Forming Cells

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. A Broad Range of Meropenem (MRP) Minimum Inhibitory Concentration (MIC) Values and Moderate Biofilm-Formation Was Determined across the K. pneumoniae Isolates
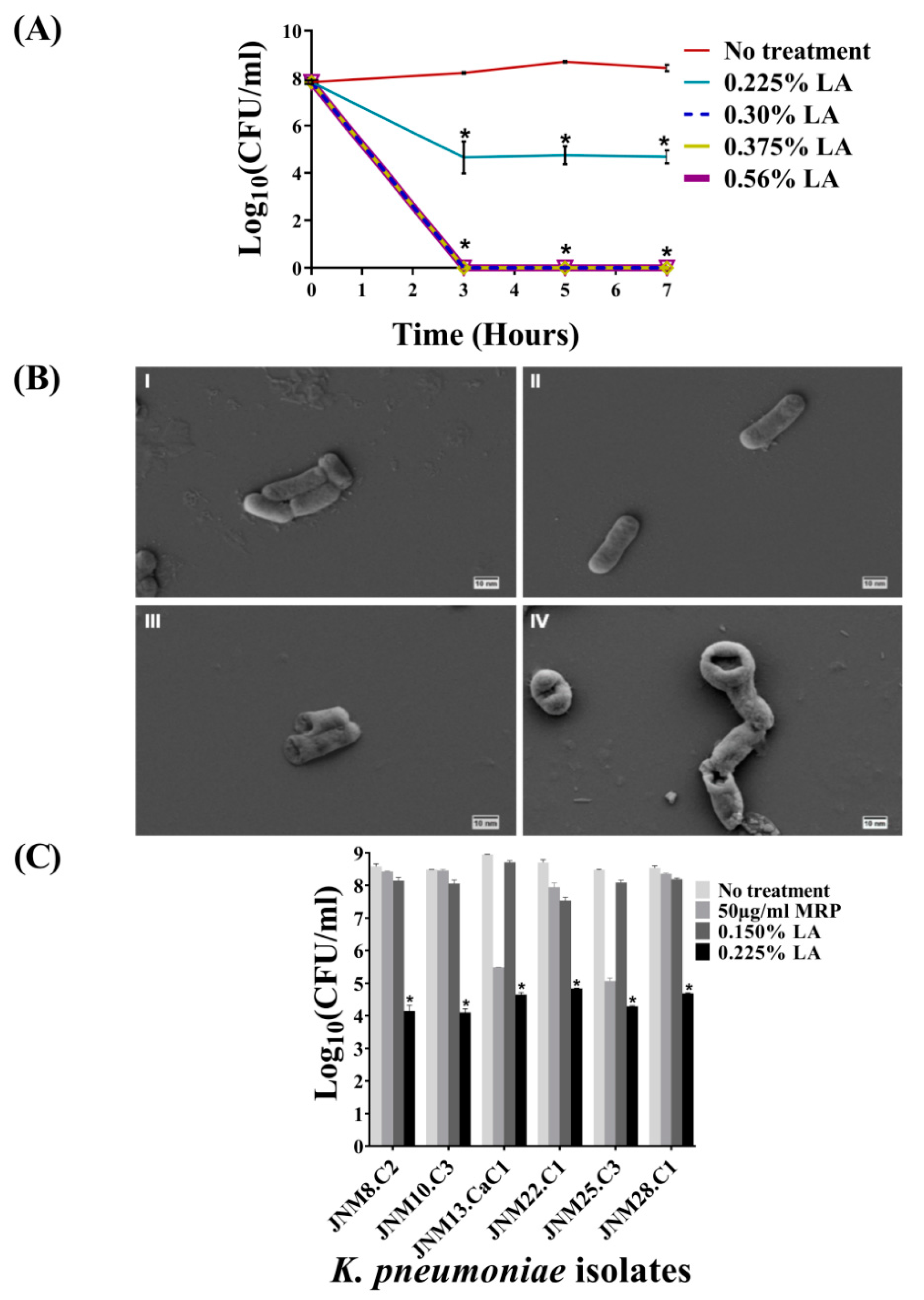
2.2. Bactericidal Activity of LA against Planktonic K. pneumoniae, JNM11.C4 Cells In Vitro
2.3. Treatment with LA Caused Membrane Damage
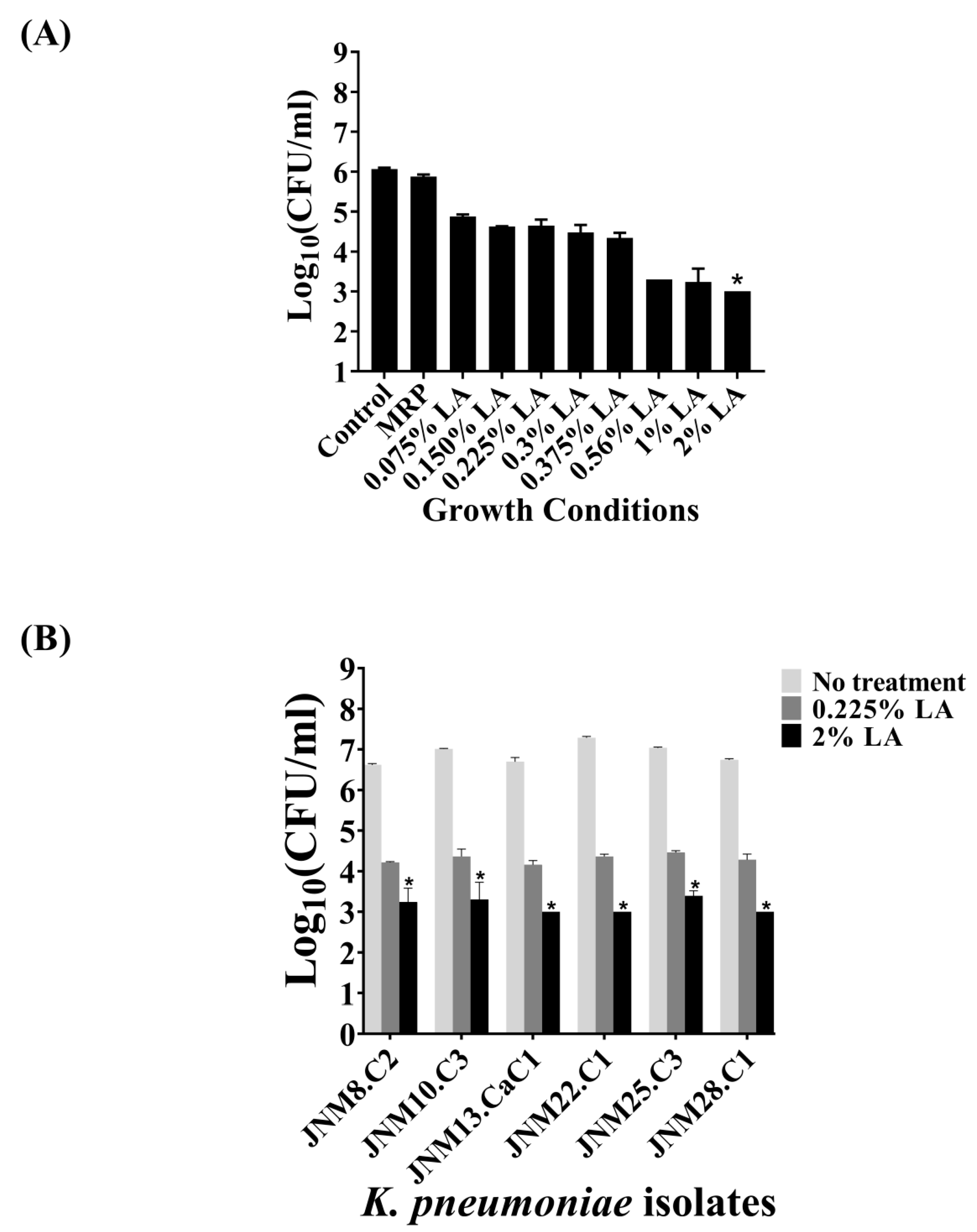
2.4. Bactericidal Activity of LA across K. pneumoniae Isolates
2.5. Bactericidal Activity of LA against Sessile K. pneumoniae Cells Forming Biofilms
3. Discussion
4. Materials and Methods
4.1. Determination of Minimum Inhibitory Concentration (MIC) Values of Meropenem (MRP)
4.2. Analyses of Biofilm Formation
4.3. Time Course LA MIC Determination Assays on Planktonic Cells
4.4. Scanning Electron Microscopy (SEM)
4.5. Validation of Bactericidal Activity
4.6. LA MIC Determination Assays on Biofilm-Forming Cells and Validation
4.7. Statistical Tests
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgements
Conflicts of Interest
References
- Lomonaco, S.; Crawford, M.A.; Lascols, C.; Timme, R.E.; Anderson, K.; Hodge, D.R.; Fisher, D.J.; Pillai, S.P.; Morse, S.A.; Khan, E.; et al. Resistome of carbapenem- and colistin-resistant Klebsiella pneumoniae clinical isolates. PLoS ONE 2018, 13, e0198526. [Google Scholar] [CrossRef] [PubMed]
- Holt, K.E.; Wertheim, H.; Zadoks, R.N.; Baker, S.; Whitehouse, C.A.; Dance, D.; Jenney, A.; Connor, T.R.; Hsu, L.Y.; Severin, J.; et al. Genomic analysis of diversity, population structure, virulence, and antimicrobial resistance in Klebsiella pneumoniae, an urgent threat to public health. Proc. Natl. Acad. Sci. USA 2015, 112, E3574–E3581. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, B.; Bardhan, T.; Chakraborty, M.; Basu, M. Resistance profiles and resistome mapping of multidrug resistant carbapenem-hydrolyzing Klebsiella pneumoniae strains isolated from the nares of preterm neonates. Int. J. Antimicrob. Agents 2019, 53, 535–537. [Google Scholar] [CrossRef] [PubMed]
- Paczosa, M.K.; Mecsas, J. Klebsiella pneumosniae: Going on the Offense with a Strong Defense. Microbiol. Mol. Biol. Rev. 2016, 80, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Choudhuri, A.H.; Chakravarty, M.; Uppal, R. Epidemiology and characteristics of nosocomial infections in critically ill patients in a tertiary care Intensive Care Unit of Northern India. Saudi J. Anaesth. 2017, 11, 402–407. [Google Scholar] [CrossRef]
- Selina, F.; Talha, K.A.; Islam, A.; Hasan, Z.; Hyder, M.; Selvapandian, S. Organisms associated with ventilator associated pneumonia (VAP) in intensive care units (ICU). J. Bangladesh Soc. Anaesth. 2014, 22, 72–77. [Google Scholar] [CrossRef]
- Magill, S.S.; Edwards, J.R.; Bamberg, W.; Beldavs, Z.G.; Dumyati, G.; Kainer, M.A.; Lynfield, R.; Maloney, M.; McAllister-Hollod, L.; Nadle, J.; et al. Multistate point-prevalence survey of health care-associated infections. N. Engl. J. Med. 2014, 370, 1198–1208. [Google Scholar] [CrossRef]
- Podschun, R.; Ullmann, U. Klebsiella spp. as nosocomial pathogens: Epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin. Microbiol. Rev. 1998, 11, 589–603. [Google Scholar] [CrossRef]
- Lee, C.; Lee, J.; Park, K.; Kim, Y.; Jeong, B.; Lee, S. Global Dissemination of Carbapenemase-Producing Klebsiella pneumoniae: Epidemiology, Genetic Context, Treatment Options, and Detection Methods. Front. Microbiol. 2016, 3, 895. [Google Scholar] [CrossRef]
- Balagopal, A.B.; Sundar, K.; Shetty, P.H. Effect of organic acids on biofilm formation and quorum signaling of pathogens from fresh fruits and vegetables. Microb. Pathog. 2017, 111, 156–162. [Google Scholar] [CrossRef]
- Komesu, A.; de Oliveira, J.A.R.; da Silva Martins, L.H.; Wolf Maciel, M.R.; Filho, R.M. Lactic acid production to purification: A review. BioRes 2017, 12, 4364–4383. [Google Scholar] [CrossRef]
- Alsaheb, R.A.A.; Aladdin, A.; Othman, N.Z.; Malek, R.A.; Leng, O.M.; Aziz, R.; Enshasy, H.A. Lactic acid applications in pharmaceutical and cosmeceutical industries. J. Chem. Pharm. Res. 2015, 7, 729–735. [Google Scholar]
- De Oliveira, E.F.; Cossu, A.; Tikekar, R.V; Nitin, N. Enhanced Antimicrobial Activity Based on a Synergistic Combination of Sublethal Levels of Stresses Induced by UV-A Light and Organic Acids. Appl. Environ. Microbiol. 2017, 83, e00383-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chotigarpa, R.; Lampang, K.N.; Pikulkaew, S.; Okonogi, S.; Ajariyakhajorn, K.; Mektrirat, R. Inhibitory Effects and Killing Kinetics of Lactic Acid Rice Gel Against Pathogenic Bacteria Causing Bovine Mastitis. Sci. Pharm. 2018, 86, 29. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chang, T.; Yang, H.; Cui, M. Antibacterial mechanism of lactic acid on physiological and morphological properties of Salmonella enteritidis, Escherichia coli and Listeria monocytogenes. Food Control 2015, 47, 231–236. [Google Scholar] [CrossRef]
- Limoli, D.H.; Jones, C.J.; Wozniak, D.J. Bacterial extracellular polysaccharides in biofilm formation and function. Microbiol. Spectrum 2015, 3, MB-0011-2014. [Google Scholar] [CrossRef]
- Bridier, A.; Briandet, R.; Thomas, V.; Dubois-Brissonnet, F. Resistance of bacterial biofilms to disinfectants: A review. Biofouling 2011, 27, 1017–1032. [Google Scholar] [CrossRef]
- Du, X.; He, F.; Shi, Q.; Zhao, F.; Xu, J.; Fu, Y.; Yu, Y. The Rapid Emergence of Tigecycline Resistance in blaKPC–2 Harboring Klebsiellapneumoniae, as Mediated in Vivo by Mutation in tetA During Tigecycline Treatment. Front. Microbiol. 2018, 9, 648. [Google Scholar] [CrossRef]
- Davies, M.; Walsh, T.R. A colistin crisis in India. Lancet Infect. Dis. 2018, 18, 256–257. [Google Scholar] [CrossRef]
- World Health Organization. Best Practice Protocols Clinical Procedures Safety. Available online: https://www.who.int/surgery/publications/OrganizationManagement.pdf (accessed on 14 August 2019).
- Cowan, R.E. Cleaning and disinfection of equipment for gastrointestinal endoscopy. Report of a Working Party of the British Society of Gastroenterology Endoscopy Committee. Gut 1998, 42, 585–593. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; 23rd informational supplement; CLSI M100-S27; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- De Araújo, L.C.A.; da Purificação-Júnior, A.F.; da Silva, S.M.; Lopes, A.C.S.; Veras, D.L.; Alves, L.C.; Dos Santos, F.B.; Napoleão, T.H.; Dos Santos Correia, M.T.; da Silva, M.V.; et al. In vitro evaluation of mercury (Hg2+) effects on biofilm formation by clinical and environmental isolates of Klebsiella pneumoniae. Ecotoxicol. Environ. Saf. 2019, 169, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Alcántar-Curiel, M.D.; Blackburn, D.; Saldaña, Z.; Gayosso-Vázquez, C.; Iovine, N.M.; De la Cruz, M.A.; Girón, J.A. Multi-functional analysis of Klebsiella pneumoniae fimbrial types in adherence and biofilm formation. Virulence 2013, 4, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Pournaras, S.; Kristo, I.; Vrioni, G.; Ikonomidis, A.; Poulou, A.; Petropoulou, D.; Tsakris, A. Characteristics of meropenemheteroresistance in Klebsiella pneumoniae carbapenemase (KPC)-producing clinical isolates of K. pneumoniae. J. Clin. Microbiol. 2010, 48, 2601–2604. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bardhan, T.; Chakraborty, M.; Bhattacharjee, B. Bactericidal Activity of Lactic Acid against Clinical, Carbapenem-Hydrolyzing, Multi-Drug-Resistant Klebsiella pneumoniae Planktonic and Biofilm-Forming Cells. Antibiotics 2019, 8, 181. https://doi.org/10.3390/antibiotics8040181
Bardhan T, Chakraborty M, Bhattacharjee B. Bactericidal Activity of Lactic Acid against Clinical, Carbapenem-Hydrolyzing, Multi-Drug-Resistant Klebsiella pneumoniae Planktonic and Biofilm-Forming Cells. Antibiotics. 2019; 8(4):181. https://doi.org/10.3390/antibiotics8040181
Chicago/Turabian StyleBardhan, Taniya, Madhurima Chakraborty, and Bornali Bhattacharjee. 2019. "Bactericidal Activity of Lactic Acid against Clinical, Carbapenem-Hydrolyzing, Multi-Drug-Resistant Klebsiella pneumoniae Planktonic and Biofilm-Forming Cells" Antibiotics 8, no. 4: 181. https://doi.org/10.3390/antibiotics8040181