

Antimicrobial and Adjuvant Potencies of Di-n-alkyl Substituted Diazalariat Ethers
 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. The Compounds and Organisms
2.2. Lariat Ethers and Derivatives’ Activities against E. coli DH5α
2.3. Solubility of Lariat Ethers and Salts
2.4. Minimum Inhibitory Concentration (MIC) Studies
2.5. Membrane Permeability
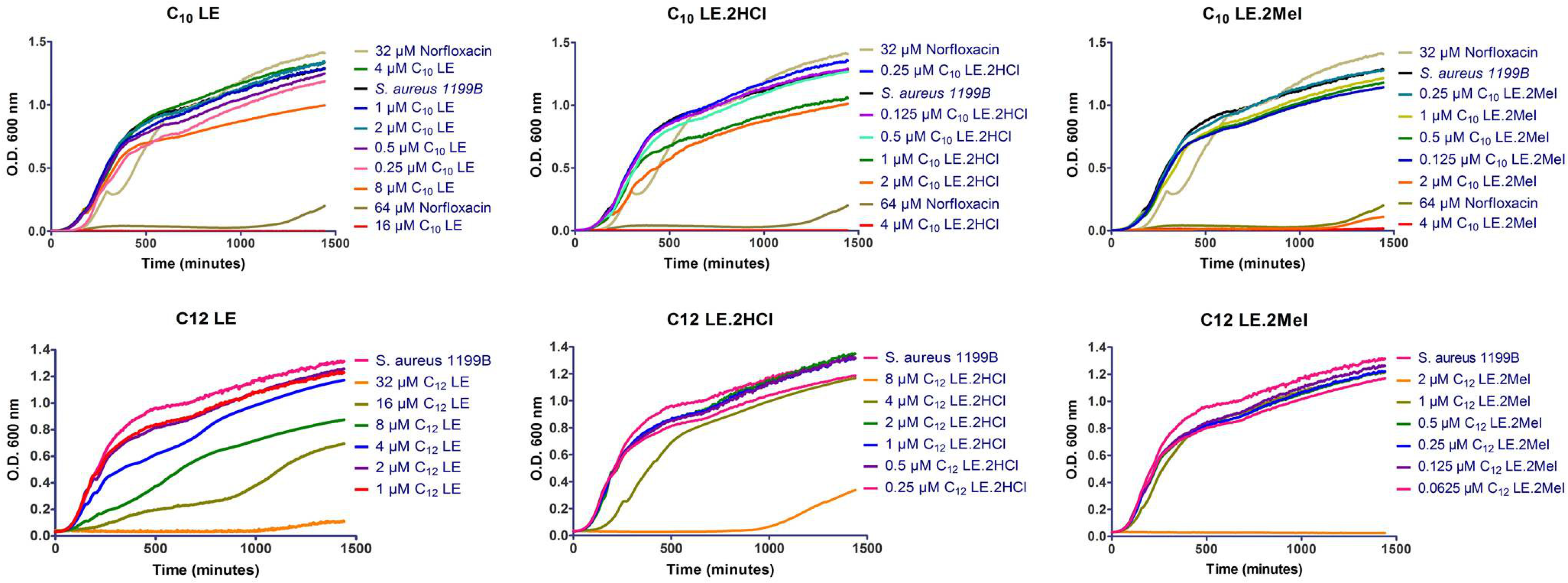
2.6. Growth Curves
2.7. Combination Studies
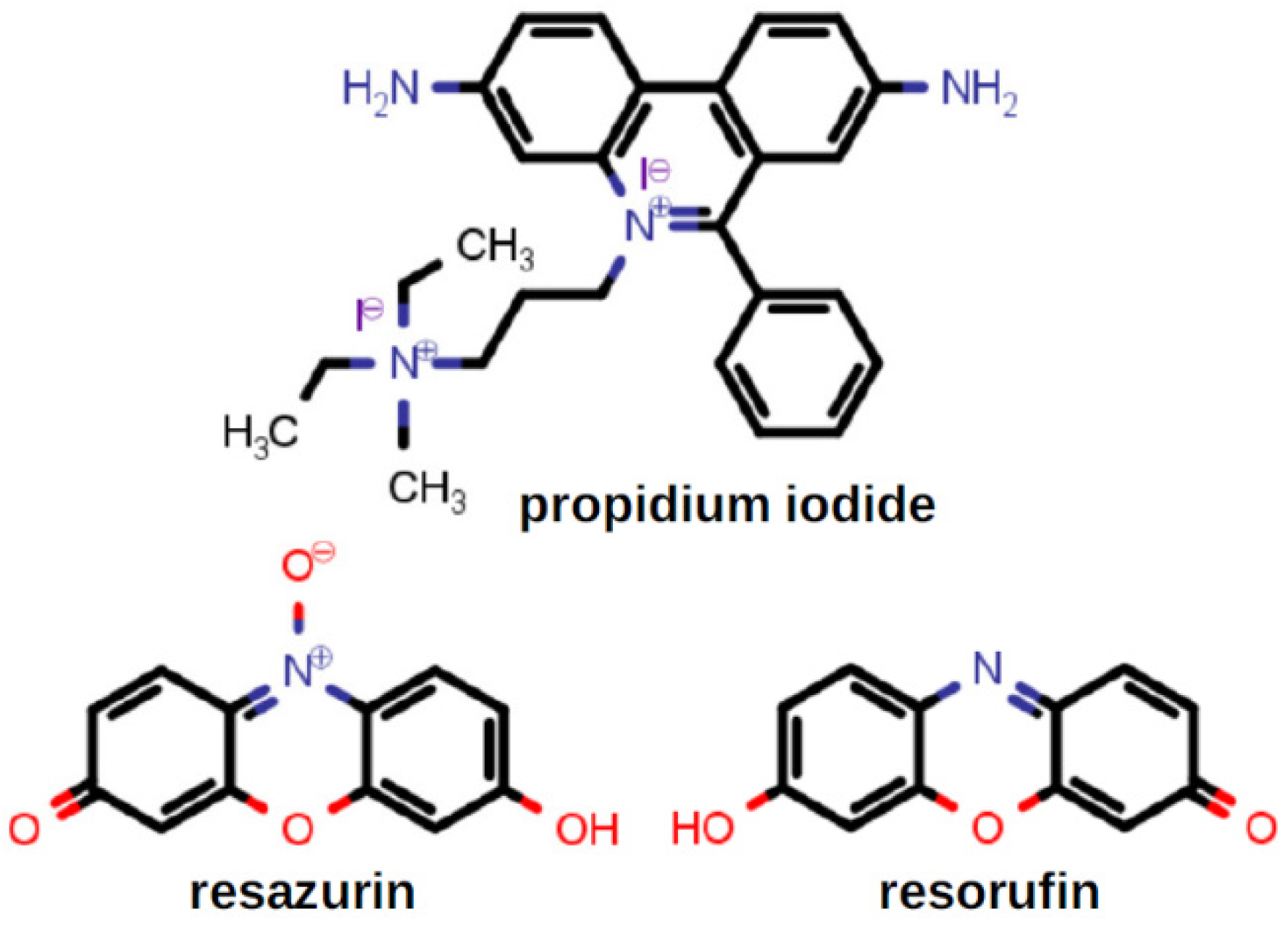
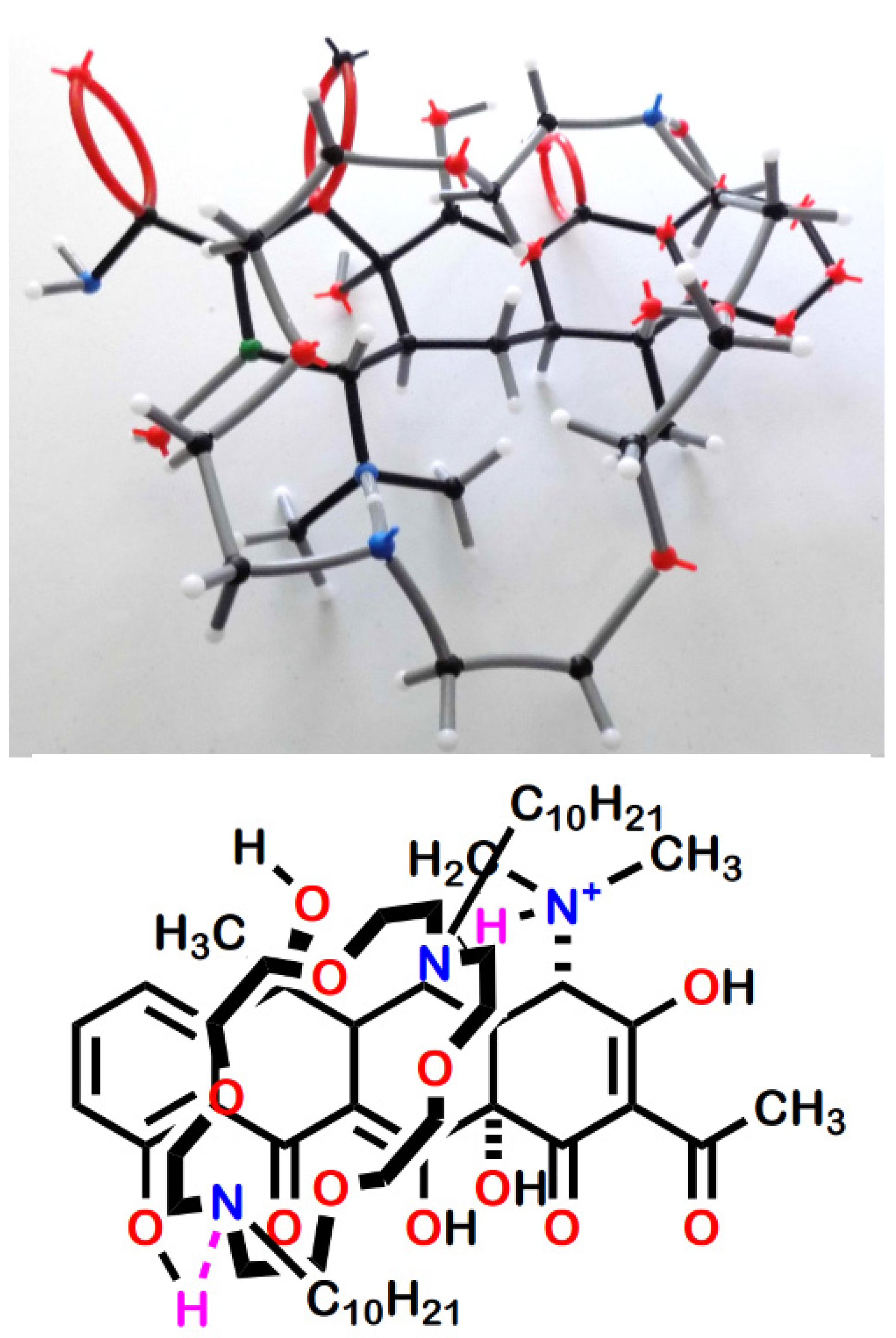
2.8. Complexation of Antimicrobials by Lariat Ethers
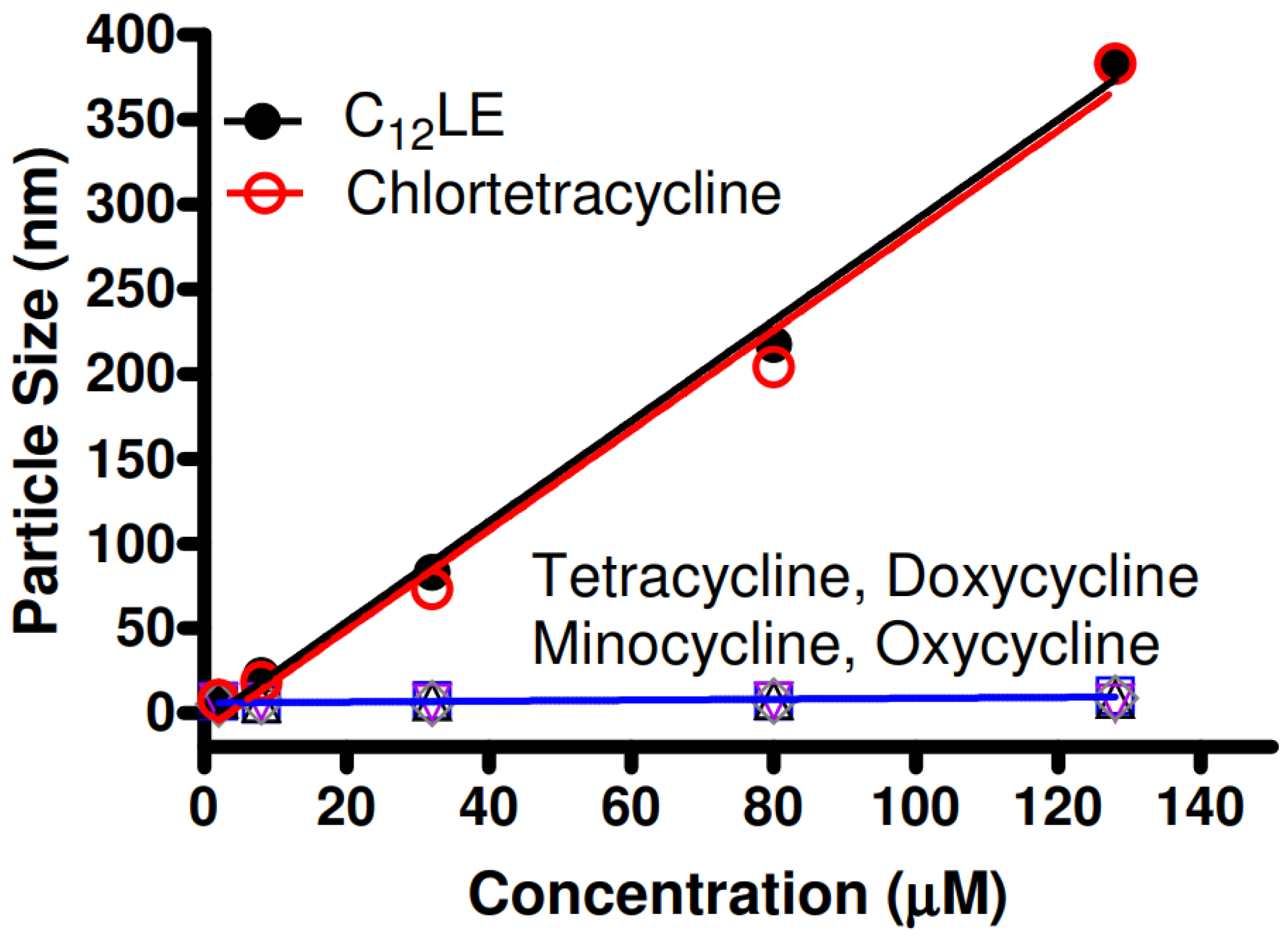
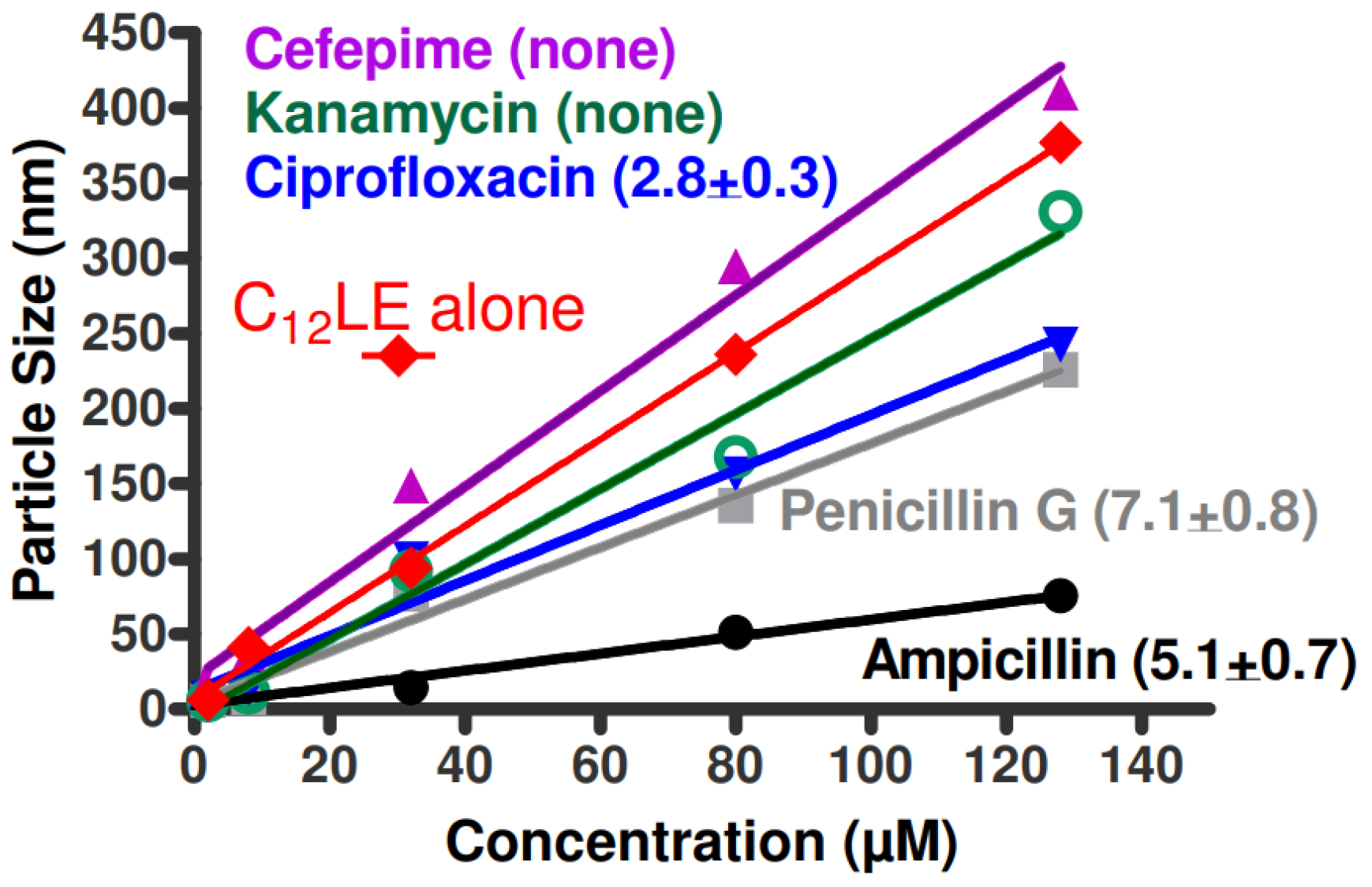
2.9. Dynamic Light Scattering (DLS)
2.10. Membrane Depolarization
2.11. Planar Bilayer Voltage Clamp Study
2.12. Efflux Pump Inhibition
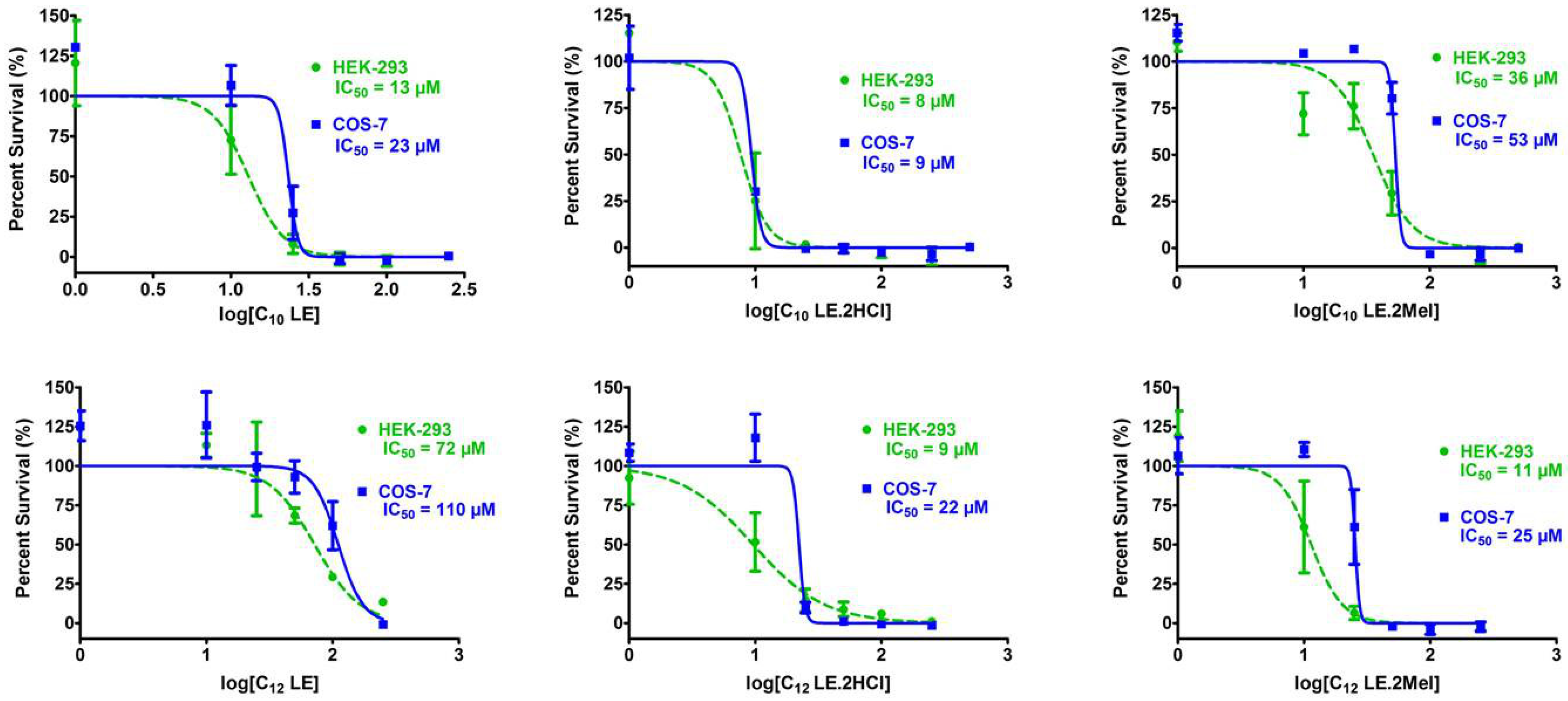
2.13. Cytotoxicity of 1–6
3. Materials and Methods
3.1. Compound Preparation
3.2. Minimum Inhibitory Concentrations
3.3. Checkerboard Experiments
3.4. Membrane Permeability Studies
3.5. pH Study
3.6. Growth Curves
3.7. Antimicrobial Complexation—Visual and NMR
3.8. Dynamic Light Scattering
3.9. Membrane Depolarization
3.10. Planar Bilayer Voltage Clamp Study
3.11. Efflux Pump Study
3.12. Cytotoxicity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- U.S. Department of Health and Human Services. Antibiotic Threats in the United States 2019; Centers for Disease Control and Prevention: Washington, DC, USA, 2019; 150p.
- World Health Organization. Antibiotic Resistance, 31 July 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/antibiotic-resistance (accessed on 20 July 2023).
- Akram, F.; Imtiaz, M.; Haq, I.U. Emergent crisis of antibiotic resistance: A silent pandemic threat to 21(st) century. Microb. Pathog. 2023, 174, 105923. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention, 2022, Antibiotic Resistance: A Global Threat. Available online: https://www.cdc.gov/drugresistance/solutions-initiative/stories/ar-global-threat.html (accessed on 8 July 2023).
- Rosen, T.J. Drugs Dermatol. Antibiotic resistance: An editorial review with recommendations. J. Drugs Dermatol. 2011, 10, 724–733. [Google Scholar] [PubMed]
- Hansson, K.; Brenthel, A. Imagining a post-antibiotic era: A cultural analysis of crisis and antibiotic resistance. Med. Humanit. 2022, 48, 381–388. [Google Scholar] [CrossRef]
- Zasloff, M. Magainins, a class of antimicrobial peptides from Xenopus skin: Isolation, characterization of two active forms, and partial cDNA sequence of a precursor. Proc. Natl. Acad. Sci. USA 1987, 84, 5449. [Google Scholar] [CrossRef]
- Wang, G. (Ed.) Antimicrobial Peptides: Discovery, Design, and Novel Therapeutic Strategies; CABI: Boston, MA, USA, 2017. [Google Scholar]
- Marsault, E.; Petersen, M.L. (Eds.) Practical Medicinal Chemistry with Macrocycles; Wiley: New York, NY, USA, 2007. [Google Scholar]
- Dutton, C.J.; Haxell, M.A.; McArthur, H.A.I.; Wax, R.G. (Eds.) Peptide Antibiotics, Discovery, Modes of Action, and Applications; Marcel Dekker: New York, NY, USA, 2002; 296p. [Google Scholar]
- Wang, H.; Zhang, C.; Li, M.; Liu, C.; Wang, J.; Ou, X.; Han, Y. Antimicrobial Peptides Mediate Apoptosis by Changing Mitochondrial Membrane Permeability. Int. J. Mol. Sci. 2022, 23, 12732. [Google Scholar] [CrossRef] [PubMed]
- Mandity, I.M.; Fulop, F. An overview of peptide and peptoid foldamers in medicinal chemistry. Expert. Opin. Drug. Discov. 2015, 10, 1163. [Google Scholar] [CrossRef] [PubMed]
- Mangold, S.L.; Grubbs, R.H. Stereoselective synthesis of macrocyclic peptides via a dual olefin metathesis and ethenolysis approach. Chem. Sci. 2015, 6, 4561. [Google Scholar] [CrossRef] [PubMed]
- Leong, B.K.; Ts’o, T.O.; Chenoweth, M.B. Testicular atrophy from inhalation of ethylene oxide cyclic tetramer. Toxicol. Appl. Pharmacol. 1974, 27, 342–354. [Google Scholar] [CrossRef]
- Takayama, K.; Hasegawa, S.; Sasagawa, S.; Nambu, N.; Nagai, T. Apparent oral toxicity of 18-crown-6 in dogs. Chem. Pharm. Bull. 1977, 25, 3125–3130. [Google Scholar] [CrossRef]
- Hendrixson, R.R.; Mack, M.P.; Palmer, R.A.; Ottolenghi, A.; Ghirardelli, R.G. Oral toxicity of the cyclic polyethers—12-crown-4, 15-crown-5, and 18-crown-6--in mice. Toxicol. Appl. Pharmacol. 1978, 44, 263–268. [Google Scholar] [CrossRef]
- Yagi, K.; Garcia, V.; Rivas, M.E.; Salas, J.; Camargo, A.; Tabata, T. Antifungal activity of crown ethers. J. Incl. Phenom. 1984, 2, 179–184. [Google Scholar] [CrossRef]
- Kato, N. Antibacterial action of alkyl-substituted crown ethers. Kenkyu Kiyo-Konan Joshi Daigaku 1985, 585–596. [Google Scholar]
- Bethge, E.W.; Bohuslavizki, K.H.; Hänsel, W.; Kneip, A.; Koppenhôfer, E. Effects of some potassium channel blockers on the ionic currents in myelinated nerve. Gen. Physiol. Biophys. 1991, 10, 225–244. [Google Scholar] [PubMed]
- Ozay, H.; Yildiz, M.; Unver, H.; Dulger, B. Synthesis, spectral studies, antimicrobial activity and crystal structures of phosphaza-lariat ethers. Asian J. Chem. 2011, 23, 2430–2436. [Google Scholar]
- Eshghi, H.; Rahimizadeh, M.; Zokaei, M.; Eshghi, S.; Eshghi, S.; Faghihi, Z.; Tabasi, E.; Kihanyan, M. Synthesis and antimicrobial activity of some new macrocyclic bis-sulfonamide and disulfides. Eur. J. Chem. 2011, 2, 47–50. [Google Scholar] [CrossRef]
- Kralj, M.; Majerski, K.; Ramljak, S.; Marjanovic, M. Adamantane Derivatives of Aza-Crown Ethers and Their Use in Treatment of Tumor. U.S. Patent 8,389,505, 5 March 2013. [Google Scholar]
- Le, T.A.; Truong, H.H.; Thi, T.P.N.; Thi, N.D.; To, H.T.; Thia, H.P.; Soldatenkov, A.T. Synthesis and biological activity of (gamma-arylpyridino)-dibenzoaza-14-crown-4 ethers. Mendeleev Commun. 2015, 25, 224–225. [Google Scholar] [CrossRef]
- Buhner, S.H. Herbal Antibiotics: Natural Alternatives for Treating Drug-Resistant Bacteria, 2nd ed.; Storey Publishing, LLC: New York, NY, USA, 2012; 480p. [Google Scholar]
- Krishnamurthy, S.; Wohlford-Lenane, C.; Kandimalla, S.; Sartre, G.; Meyerholz, D.K.; Theberge, V.; Hallee, S.; Duperre, A.M.; Del’Guidice, T.; Lepetit-Stoffaes, J.P.; et al. Engineered amphiphilic peptides enable delivery of proteins and CRISPR-associated nucleases to airway epithelia. Nat. Commun. 2019, 10, 4906. [Google Scholar] [CrossRef]
- Riahifard, N.; Mozaffari, S.; Aldakhil, T.; Nunez, F.; Alshammari, Q.; Alshammari, S.; Yamaki, J.; Parang, K.; Tiwari, R.K. Design, Synthesis, and Evaluation of Amphiphilic Cyclic and Linear Peptides Composed of Hydrophobic and Positively-Charged Amino Acids as Antibacterial Agents. Molecules 2018, 23, 2722. [Google Scholar] [CrossRef]
- Kabelka, I.; Vacha, R. Optimal conditions for opening of membrane pore by amphiphilic peptides. J. Chem. Phys. 2015, 143, 243115. [Google Scholar] [CrossRef]
- Oh, D.; Sun, J.; Nasrolahi Shirazi, A.; LaPlante, K.L.; Rowley, D.C.; Parang, K. Antibacterial activities of amphiphilic cyclic cell-penetrating peptides against multidrug-resistant pathogens. Mol. Pharm. 2014, 11, 3528. [Google Scholar] [CrossRef]
- Wiradharma, N.; Khoe, U.; Hauser, C.A.; Seow, S.V.; Zhang, S.; Yang, Y.Y. Synthetic cationic amphiphilic alpha-helical peptides as antimicrobial agents. Biomaterials 2011, 32, 2204. [Google Scholar] [CrossRef]
- Dennison, S.R.; Phoenix, D.A. Amphiphilic peptides structures. Protein Pept. Lett. 2010, 17, 1311. [Google Scholar]
- Zasloff, M. Antimicrobial peptides and suppression of apoptosis in human skin. J. Investig. Dermatol. 2009, 129, 824. [Google Scholar] [CrossRef]
- Wolf, L.M.; Servoss, S.L.; Moss, M.A. Peptoids: Emerging therapeutics for eurodegeneration. J. Neurol. Neuromed. 2017, 2, 1–5. [Google Scholar]
- Zhang, Y.; Heidari, Z.; Su, Y.; Yu, T.; Xuan, S.; Omarova, M.; Aydin, Y.; Dash, S.; Zhang, D.; John, V. Amphiphilic Polypeptoids Rupture Vesicle Bilayers To Form Peptoid-Lipid Fragments Effective in Enhancing Hydrophobic Drug Delivery. Langmuir 2019, 35, 15335. [Google Scholar] [CrossRef] [PubMed]
- Spicer, S.K.; Subramani, A.; Aguila, A.L.; Green, R.M.; McClelland, E.E.; Bicker, K.L. Toward a clinical antifungal peptoid: Investigations into the therapeutic potential of AEC5. Biopolymers 2019, 110, e23276. [Google Scholar] [CrossRef] [PubMed]
- Mattes, D.S.; Streit, B.; Bhandari, D.R.; Greifenstein, J.; Foertsch, T.C.; Munch, S.W.; Ridder, B.; Bojničić-Kninski, C.V.; Nesterov-Mueller, A.; Spengler, B.; et al. Combinatorial Synthesis of Peptoid Arrays via Laser-Based Stacking of Multiple Polymer Nanolayers. Macromol. Rapid Commun. 2019, 40, e1800533. [Google Scholar] [CrossRef]
- Webster, A.M.; Cobb, S.L. Recent Advances in the Synthesis of Peptoid Macrocycles. Chemistry 2018, 24, 7560. [Google Scholar] [CrossRef]
- Zuckermann, R.N. Peptoid origins. Biopolymers 2011, 96, 545. [Google Scholar] [CrossRef]
- Gomes, A.M.M.; Costa, P.J.; Machuqueiro, M. Recent advances on molecular dynamics-based techniques to address drug membrane permeability with atomistic detail. BBA Adv. 2023, 4, 100099. [Google Scholar] [CrossRef]
- Li, J.; Lu, H.; Wang, A.; Wen, X.; Huang, Y.; Li, Q. The fates of antibiotic resistance genes and their association with cell membrane permeability in response to peroxydisulfate during composting. J. Environ. Manag. 2023, 344, 118659. [Google Scholar] [CrossRef]
- Baena-Santillan, E.S.; Piloni-Martini, J.; Rangel-Vargas, E.; Gomez-Aldapa, C.A.; Sanchez-Gutierrez, M.; Madrigal-Santillan, E.O.; Castro-Rosas, J. Comparison of the Antibacterial Activity and Effect on Membrane Permeability of Hibiscus Acid and a Commercial Chlorhexidine Mouthrinse Against Pathogenic Oral Bacteria and Determination of Hibiscus Acid Toxicity. J. Med. Food 2022, 25, 324–328. [Google Scholar] [CrossRef]
- Cui, P.; Niu, H.; Shi, W.; Zhang, S.; Zhang, H.; Margolick, J.; Zhang, W.; Zhang, Y. Disruption of Membrane by Colistin Kills Uropathogenic E. coli Persisters and Enhances Killing of Other Antibiotics. Antimicrob. Agents Chemother. 2016, 60, 6867–6871. [Google Scholar] [CrossRef] [PubMed]
- Kelesidis, T.; Falagas, M.E. The safety of polymyxin antibiotics. Expert Opin. Drug Saf. 2015, 14, 1687. [Google Scholar] [CrossRef]
- Gokel, G.W. Hydraphiles: Design, Synthesis, and Analysis of a Family of Synthetic, Cation-Conducting Channels. Chem. Comm. 2000, 1–9. [Google Scholar] [CrossRef]
- Patel, M.B.; Garrad, E.G.; Meisel, J.W.; Negin, S.; Gokel, M.R.; Gokel, G.W. Synthetic Ionophores as Non-Resistant Antibiotic Adjuvants. RSC Adv. 2019, 9, 2217–2230. [Google Scholar] [CrossRef] [PubMed]
- Gokel, G.W.; Gokel, M.R.; Negin, S.; Patel, M.B. Molecules That Inhibit Efflux Pumps in Multi-Drug Resistant Bacteria and Uses Thereof. United States Patent 10,463,044 B2, 5 November 2019. [Google Scholar]
- MIC. M07-A9, Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, 9th ed.; Approved standard; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012; ISBN 1-56238-784-7. Available online: www.clsi.org (accessed on 10 July 2023).
- Inoue, Y.; Gokel, G.W. (Eds.) Cation Binding by Macrocycles; Marcel Dekker: New York, NY, USA, 1990; 761p. [Google Scholar]
- Leevy, W.M.; Weber, M.E.; Gokel, M.R.; Hughes-Strange, G.B.; Daranciang, D.D.; Ferdani, R.; Gokel, G.W. Correlation of bilayer membrane cation transport and biological activity in alkyl-substituted lariat ethers. Org. Biomol. Chem. 2005, 3, 1647. [Google Scholar] [CrossRef] [PubMed]
- Carrasquel-Ursulaez, W.; Reeves, R.D.; Dehghany, M.; Jones, C.; Schomaker, J.M.; Chanda, B. Re-evaluation of the mechanism of cytotoxicity of dialkylated lariat ether compounds. RSC Adv. 2020, 10, 40391. [Google Scholar] [CrossRef]
- Kumar, G.; Kiran Tudu, A. Tackling multidrug-resistant Staphylococcus aureus by natural products and their analogues acting as NorA efflux pump inhibitors. Bioorg. Med. Chem. 2023, 80, 117187. [Google Scholar] [CrossRef]
- Meisel, J.W.; Patel, M.B.; Garrad, E.; Stanton, R.A.; Gokel, G.W. Reversal of Tetracycline Resistance in Escherichia coli by Noncytotoxic bis(Tryptophan)s. J. Am. Chem. Soc. 2016, 138, 10571–10577. [Google Scholar] [CrossRef]
- Berge, S.M.; Bighley, L.D.; Monkhouse, D.C. Pharmaceutical salts. J. Pharm. Sci. 1977, 66, 1–19. [Google Scholar] [CrossRef]
- Swarbrick, J.; Boylan, J.C. (Eds.) Encyclopedia of Pharmaceutical Technology; Marcel Dekker: New York, NY, USA, 1996; Volume 13, p. 453. [Google Scholar]
- Ahman, J.; Matuschek, E.; Kahlmeter, G. Evaluation of ten brands of pre-poured Mueller-Hinton agar plates for EUCAST disc diffusion testing, Clin. Microbiol. Infect. 2022, 28, 1499.e1. [Google Scholar]
- Negin, S.; Gokel, M.R.; Patel, M.B.; Sedinkin, S.L.; Osborn, D.C.; Gokel, G.W. The Aqueous Medium-Dimethylsulfoxide Conundrum in Biological Studies. RSC Adv. 2015, 5, 8088–8093. [Google Scholar] [CrossRef]
- O’Neil, O.J. The Merck Index, 14th ed.; Merck Research Laboratories: Boston, MA, USA, 2006. [Google Scholar]
- De Wall, S.W.; Barbour, L.; Gokel, G.W. Solid state bilayer formation from a dialkyl-substituted lariat ether that forms stable vesicles in aqueous suspension. J. Phys. Org. Chem. 2001, 14, 383–391. [Google Scholar] [CrossRef]
- Zhang, N.; Shan, W.; Gao, L.; Kou, S.H.; Lu, C.; Yang, H.; Peng, B.; Tam, K.Y.; Lee, L.T.O.; Zheng, J. Repurposing the Hedgehog pathway inhibitor, BMS-833923, as a phosphatidylglycerol-selective membrane-disruptive colistin adjuvant against ESKAPE pathogens. Int. J. Antimicrob. Agents 2023, 62, 106888. [Google Scholar] [CrossRef]
- Gokel, G.W.; Gokel, M.R.; Negin, S.; Patel, M.B. Enhancement of Antibiotic Activity. U.S. Patent 10,791,740 B2, 6 October 2020. [Google Scholar]
- Kumari, H.; Negin, S.; Eisenhart, A.; Patel, M.B.; Beck, T.L.; Heinrich, F.; Spikes, H.J.; Gokel, G.W. Assessment of a host–guest interaction in a bilayer membrane model. RSC Adv. 2022, 12, 32046–32055. [Google Scholar] [CrossRef] [PubMed]
- Clegg, W.; Teat, S.J. Tetracycline hydrochloride: A synchrotron microcrystal study. Acta Crystallogr. Sect. C 2000, 56, 1343–1345. [Google Scholar] [CrossRef]
- De Wall, S.L.; Wang, K.; Berger, D.L.; Watanabe, S.; Hernandez, J.C.; Gokel, G.W. Azacrown Ethers as Amphiphile Headgroups: Formation of Stable Aggregates from Two- and Three-Armed Lariat Ethers. J. Org. Chem. 1997, 62, 6784–6791. [Google Scholar] [CrossRef]
- Wu, M.; Maier, E.; Benz, R.; Hancock, R.E.W. Mechanism of interaction of different classes of cationic antimicrobial peptides with planar bilayers and with the cytoplasmic membrane of Escherichia coli. Biochemistry 1999, 38, 7235–7242. [Google Scholar] [CrossRef]
- Zaritsky, A.; Kihara, M.; Macnab, R.M. Measurement of membrane potential in Bacillus subtilis: A comparison of lipophilic cations, rubidium ion, and a cyanine dye as probes. J. Membr. Biol. 1981, 63, 215–231. [Google Scholar] [CrossRef]
- Bakker, E.P.; Mangerich, W.E. Interconversion of components of the bacterial proton motive force by electrogenic potassium transport. J. Bacteriol. 1981, 147, 820–826. [Google Scholar] [CrossRef] [PubMed]
- Dinh, M.N.; Hitomi, M.; Al-Turaihi, Z.A.; Scott, J.G. Alamar Blue assay optimization to minimize drug interference and inter-assay viability. Biorxiv 2023. [Google Scholar] [CrossRef]
- Negin, S.; Patel, M.B.; Gokel, M.R.; Meisel, J.W.; Gokel, G.W. Antibiotic Potency against E. coli is Enhanced by Channel-Forming Alkyl Lariat Ethers. Chembiochem 2016, 17, 2153–2161. [Google Scholar] [CrossRef] [PubMed]
- de Morais Oliveira-Tintino, C.D.; Muniz, D.F.; Dos Santos Barbosa, C.R.; Silva Pereira, R.L.; Begnini, I.M.; Rebelo, R.A.; da Silva, L.E.; Mireski, S.L.; Nasato, M.C.; Lacowicz Krautler, M.I.; et al. NorA, Tet(K), MepA, and MsrA Efflux Pumps in Staphylococcus aureus, their Inhibitors and 1,8-Naphthyridine Sulfonamides. Curr. Pharm. Des. 2023, 29, 323. [Google Scholar]





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LE Sidearms | LE | LE•HCl | LE•MeI |
|---|---|---|---|
| n-octyl | >128 | >128 | >128 |
| n-decyl | 11 | 4 | 24 |
| n-undecyl | 20 | nd a | nd |
| n-dodecyl | >128 | >128 | 2 |
| n-tetradecyl | >128 | >128 | >128 |
| Compounds | MHII a | PBS b |
|---|---|---|
| Verapamil | >128 | >128 |
| Reserpine | 16 | 16 |
| C10LE (1) | ≥128 | ≥128 |
| C10LE•2HCl (2) | 8 | 8 |
| C10LE•2MeI (3) | 64 | ≥128 |
| C12LE (4) | 16 | 16 |
| C12LE•2HCl (5) | 4 | 2 |
| C12LE•2MeI (6) | 1–2 | ≥128 |
| Compounds Used | K12 E. coli | TetR E. coli | S. aureus 1199B |
|---|---|---|---|
| Norfloxacin | 0.125 | nd | 64 |
| Tetracycline•HCl | 2 | 1000 | nd |
| C10LE (1) | 10 | 8 | 8 |
| C10LE•2HCl (2) | 4 | 8 | 2 |
| C10LE•2MeI (3) | 16 | 32 | 1–2 |
| C12LE (4) | >128 | >64 | 32 |
| C12LE•2HCl (5) | >128 | >64 | 4 |
| C12LE•2MeI (6) | 4 | 16 | 1 |
| Compounds Used | MHII | MH | MH + Ca2+ | ||
|---|---|---|---|---|---|
| pH 6.4 | pH 7.4 | pH 8.4 | |||
| C10LE (1) | 16 | ≥16 | 32 | 16 | 16 |
| C10LE•2HCl (2) | 2 | 2 | 4 | 2 | 8 |
| C10LE•2MeI (3) | 2 | 2 | 2 | 2 | 16 |
| C12LE (4) | >16 | >32 | >64 | nd | nd |
| C12LE•2HCl (5) | 4 | 4 | >64 | nd | nd |
| C12LE•2MeI (6) | 2 | 2 | 2 | nd | nd |
| Antimicrobial | Complexation Ratio b |
|---|---|
| Tetracycline•HCl | 1.03 ± 0.02 |
| Minocycline•HCl | 1.06 ± 0.05 |
| Doxycycline•HCl | 1.02 ± 0.04 |
| Oxycycline•HCl | 0.997 ± 0.01 |
| Chlortetracycline•HCl | No complexation observed |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patel, M.B.; Spikes, H.; Bailey, R.S.; Connell, T.; Gill, H.; Gokel, M.R.; Harris, R.; Meisel, J.W.; Negin, S.; Yin, S.A.; et al. Antimicrobial and Adjuvant Potencies of Di-n-alkyl Substituted Diazalariat Ethers. Antibiotics 2023, 12, 1513. https://doi.org/10.3390/antibiotics12101513
Patel MB, Spikes H, Bailey RS, Connell T, Gill H, Gokel MR, Harris R, Meisel JW, Negin S, Yin SA, et al. Antimicrobial and Adjuvant Potencies of Di-n-alkyl Substituted Diazalariat Ethers. Antibiotics. 2023; 12(10):1513. https://doi.org/10.3390/antibiotics12101513
Chicago/Turabian StylePatel, Mohit B., Helena Spikes, Robert S. Bailey, Thomas Connell, Hannah Gill, Michael R. Gokel, Rebecca Harris, Joseph W. Meisel, Saeedeh Negin, Shanheng Andrew Yin, and et al. 2023. "Antimicrobial and Adjuvant Potencies of Di-n-alkyl Substituted Diazalariat Ethers" Antibiotics 12, no. 10: 1513. https://doi.org/10.3390/antibiotics12101513