
Virucidal Activity of Lemon Essential Oil against Feline Calicivirus Used as Surrogate for Norovirus
 , ,
, ,  , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Analysis of LEO
2.2. Gas Chromatography/Mass Spectrophotometry (GC/MS)
2.3. Compound Identification
2.4. Cells and Virus
2.5. Cytotoxicity Assay
2.6. Virucidal Activity Assay
2.7. Viral Titration
2.8. Data Analysis
3. Results
3.1. Analytical Details of LEO
3.2. Cytotoxicity
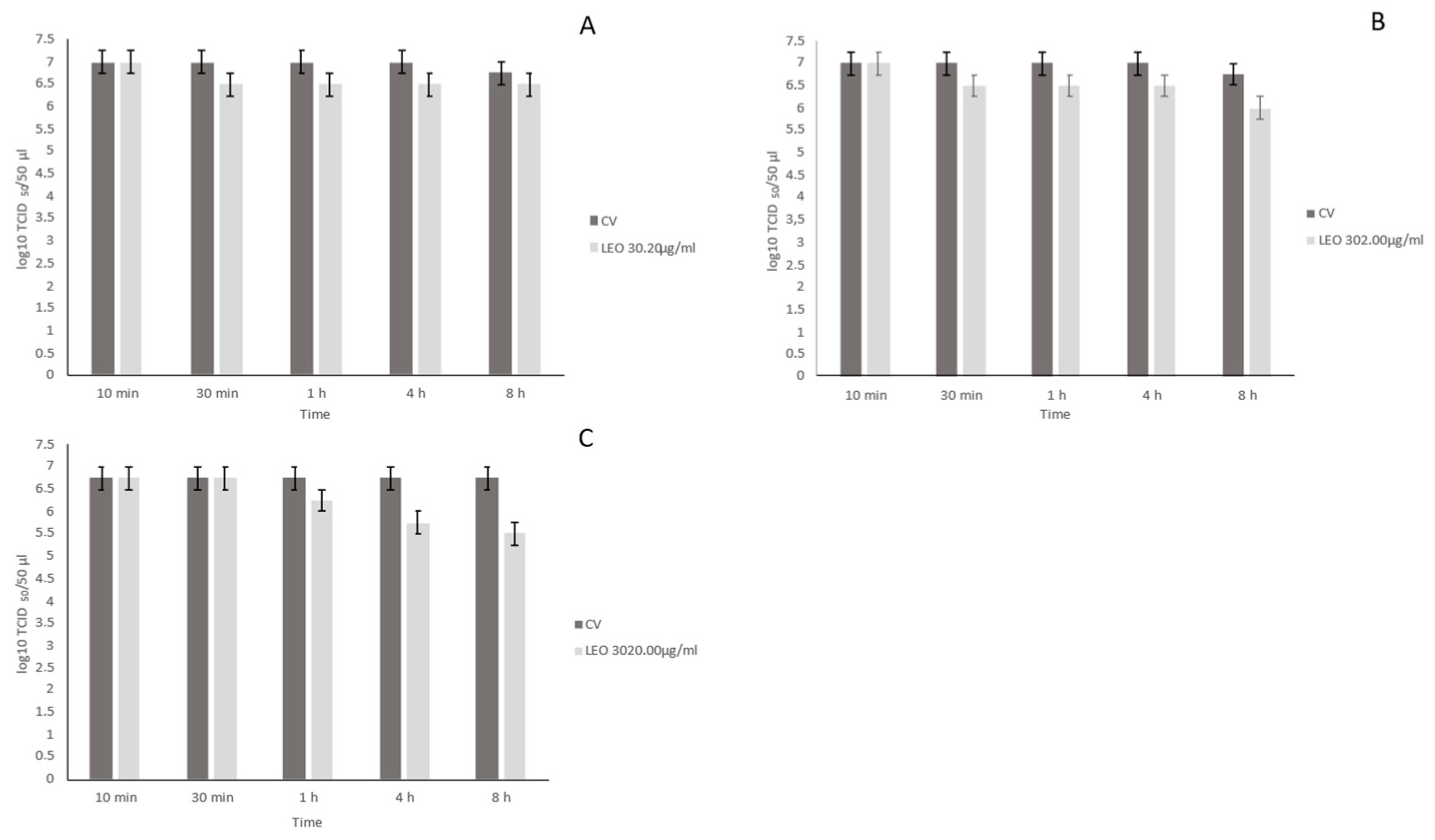
3.3. Virucidal Activity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO (Ed.) World Health Organization WHO Director-General’s Opening Remarks at the Media Briefing on COVID-19; 11 March 2020. Available online: https://www.who.int/director-general/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefing-on-covid-19---11-march-2020 (accessed on 11 January 2023).
- Mathur, S.; Hoskins, C. Drug development: Lessons from nature. Biomed. Rep. 2017, 6, 612–614. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, R.N.; Agra, M.D.F.; Maior, F.N.S.; de Sousa, D. Essential Oils and Their Constituents: Anticonvulsant Activity. Molecules 2011, 16, 2726–2742. [Google Scholar] [CrossRef] [PubMed]
- Amorati, R.; Foti, M.C.; Valgimigli, L. Antioxidant Activity of Essential Oils. J. Agric. Food Chem. 2013, 61, 10835–10847. [Google Scholar] [CrossRef] [PubMed]
- Wani, A.R.; Yadav, K.; Khursheed, A.; Rather, M.A. An updated and comprehensive review of the antiviral potential of essential oils and their chemical constituents with special focus on their mechanism of action against various influenza and coronaviruses. Microb. Pathog. 2021, 152, 104620. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, Y.; Paidari, S.; Baghbaderani, S.A.; Nateghi, L.; Al-Hassan, A.A.; Ariffin, F. Essential oils as natural antimicrobial agents in postharvest treatments of fruits and vegetables: A review. J. Food Meas. Charact. 2022, 16, 507–522. [Google Scholar] [CrossRef]
- Singh, I.; Vimanpreet, K.; Navendu, S.; Parneet, K.; Udesh, K. Essential Oils in Treatment and Management of Dental Diseases. Biointerface Res. Appl. Chem. 2021, 12, 7267–7286. [Google Scholar] [CrossRef]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential Oils in Food Preservation: Mode of Action, Synergies, and Interactions with Food Matrix Components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef]
- World Health Organization (Ed.) Food Safety 2022. Available online: https://www.who.int/news-room/fact-sheets/detail/food-safety#:~:text=Food%20safety%2C%20nutrition%20and%20food,healthy%20life%20years%20(DALYs) (accessed on 11 January 2023).
- Reichling, J.; Schnitzler, P.; Suschke, U.; Saller, R. Essential Oils of Aromatic Plants with Antibacterial, Antifungal, Antiviral, and Cytotoxic Properties—An Overview. Complement. Med. Res. 2009, 16, 79–90. [Google Scholar] [CrossRef]
- Camero, M.; Lanave, G.; Catella, C.; Capozza, P.; Gentile, A.; Fracchiolla, G.; Britti, D.; Martella, V.; Buonavoglia, C.; Tempesta, M. Virucidal activity of ginger essential oil against caprine alphaherpesvirus-1. Vet. Microbiol. 2019, 230, 150–155. [Google Scholar] [CrossRef]
- Catella, C.; Camero, M.; Lucente, M.S.; Fracchiolla, G.; Sblano, S.; Tempesta, M.; Martella, V.; Buonavoglia, C.; Lanave, G. Virucidal and antiviral effects of Thymus vulgaris essential oil on feline coronavirus. Res. Vet. Sci. 2021, 137, 44–47. [Google Scholar] [CrossRef]
- Romeo, A.; Iacovelli, F.; Scagnolari, C.; Scordio, M.; Frasca, F.; Condò, R.; Ammendola, S.; Gaziano, R.; Anselmi, M.; Divizia, M.; et al. Potential Use of Tea Tree Oil as a Disinfectant Agent against Coronaviruses: A Combined Experimental and Simulation Study. Molecules 2022, 27, 3786. [Google Scholar] [CrossRef] [PubMed]
- Cermelli, C.; Fabio, A.; Fabio, G.; Quaglio, P. Effect of Eucalyptus Essential Oil on Respiratory Bacteria and Viruses. Curr. Microbiol. 2008, 56, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Garozzo, A.; Timpanaro, R.; Bisignano, B.; Furneri, P.; Bisignano, G.; Castro, A. In vitro antiviral activity of Melaleuca alternifolia essential oil. Lett. Appl. Microbiol. 2009, 49, 806–808. [Google Scholar] [CrossRef]
- Kovač, K.; Diez-Valcarce, M.; Raspor, P.; Hernández, M.; Rodríguez-Lázaro, D. Natural Plant Essential Oils Do Not Inactivate Non-enveloped Enteric Viruses. Food Environ. Virol. 2012, 4, 209–212. [Google Scholar] [CrossRef]
- Búfalo, M.; Figueiredo, A.; de Sousa, J.; Candeias, J.; Bastos, J.; Sforcin, J. Anti-poliovirus activity of Baccharis dracunculifolia and propolis by cell viability determination and real-time PCR. J. Appl. Microbiol. 2009, 107, 1669–1680. [Google Scholar] [CrossRef]
- Elizaquível, P.; Azizkhani, M.; Aznar, R.; Sánchez, G. The effect of essential oils on norovirus surrogates. Food Control 2013, 32, 275–278. [Google Scholar] [CrossRef]
- Pilau, M.R.; Alves, S.H.; Weiblen, R.; Arenhart, S.; Cueto, A.P.; Lovato, L.T. Antiviral activity of the Lippia graveolens (Mexican oregano) essential oil and its main compound carvacrol against human and animal viruses. Braz. J. Microbiol. 2011, 42, 1616–1624. [Google Scholar] [CrossRef]
- Ahmed, S.M.; Hall, A.J.; Robinson, A.E.; Verhoef, L.; Premkumar, P.; Parashar, U.D.; Koopmans, M.; Lopman, B.A. Global prevalence of norovirus in cases of gastroenteritis: A systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 725–730. [Google Scholar] [CrossRef]
- Robilotti, E.; Deresinski, S.; Pinsky, B.A. Norovirus. Clin. Microbiol. Rev. 2015, 28, 134–164. [Google Scholar] [CrossRef]
- Netzler, N.E.; Tuipulotu, D.E.; White, P.A. Norovirus antivirals: Where are we now? Med. Res. Rev. 2019, 39, 860–886. [Google Scholar] [CrossRef]
- Terio, V.; Martella, V.; Moschidou, P.; Di Pinto, P.; Tantillo, G.; Buonavoglia, C. Norovirus in retail shellfish. Food Microbiol. 2010, 27, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Winder, N.; Gohar, S.; Muthana, M. Norovirus: An Overview of Virology and Preventative Measures. Viruses 2022, 14, 2811. [Google Scholar] [CrossRef] [PubMed]
- Parrón, I.; Barrabeig, I.; Soldevila, N.; Bartolomé, R.; Guix, S.; Rius, C.; Cornejo-Sánchez, T.; Izquierdo, C.; Domínguez, À. Outbreaks of Gastroenteritis Due to Norovirus in Schools and Summer Camps in Catalonia, 2017–2019. Microbiol. Spectr. 2022, 10, e00119-22. [Google Scholar] [CrossRef] [PubMed]
- Kanerva, M.; Maunula, L.; Lappalainen, M.; Mannonen, L.; von Bonsdorff, C.-H.; Anttila, V.-J. Prolonged norovirus outbreak in a Finnish tertiary care hospital caused by GII.4-2006b subvariants. J. Hosp. Infect. 2009, 71, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Stals, A.; Baert, L.; Van Coillie, E.; Uyttendaele, M. Evaluation of a norovirus detection methodology for soft red fruits. Food Microbiol. 2011, 28, 52–58. [Google Scholar] [CrossRef]
- Wobus, C.E.; Karst, S.M.; Thackray, L.B.; Chang, K.-O.; Sosnovtsev, S.V.; Belliot, G.; Krug, A.; Mackenzie, J.M.; Green, K.Y.; Virgin, H.W., IV. Replication of Norovirus in Cell Culture Reveals a Tropism for Dendritic Cells and Macrophages. PLOS Biol. 2004, 2, e432. [Google Scholar] [CrossRef]
- Lindesmith, L.C.; Donaldson, E.; Leon, J.; Moe, C.L.; Frelinger, J.A.; Johnston, R.E.; Weber, D.J.; Baric, R.S. Heterotypic Humoral and Cellular Immune Responses following Norwalk Virus Infection. J. Virol. 2010, 84, 1800–1815. [Google Scholar] [CrossRef]
- Ettayebi, K.; Tenge, V.R.; Cortes-Penfield, N.W.; Crawford, S.E.; Neill, F.H.; Zeng, X.-L.; Yu, X.; Ayyar, B.V.; Burrin, D.; Ramani, S.; et al. New Insights and Enhanced Human Norovirus Cultivation in Human Intestinal Enteroids. Msphere 2021, 6, e01136-22. [Google Scholar] [CrossRef]
- Hoover, E.A.; Kahn, D.E. Experimentally induced feline calicivirus infection: Clinical signs and lesions. J. Am. Vet. Med. Assoc. 1975, 166, 463–468. [Google Scholar]
- Jimenez, L.; Chiang, M. Virucidal activity of a quaternary ammonium compound disinfectant against feline calicivirus: A surrogate for norovirus. Am. J. Infect. Control 2006, 34, 269–273. [Google Scholar] [CrossRef]
- Cromeans, T.; Park, G.W.; Costantini, V.; Lee, D.; Wang, Q.; Farkas, T.; Lee, A.; Vinjé, J. Comprehensive Comparison of Cultivable Norovirus Surrogates in Response to Different Inactivation and Disinfection Treatments. Appl. Environ. Microbiol. 2014, 80, 5743–5751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, M.S. Antiviral activities of Artemisia princeps var. orientalis essential oil and its α-thujone against norovirus surrogates. Food Sci. Biotechnol. 2017, 26, 1457–1461. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.W.; You, H.J.; Lee, S.; Kim, B.; Kim, D.K.; Choi, J.-B.; Kim, J.-A.; Lee, H.J.; Joo, I.S.; Lee, J.S.; et al. Inactivation of Norovirus by Lemongrass Essential Oil Using a Norovirus Surrogate System. J. Food Prot. 2017, 80, 1293–1302. [Google Scholar] [CrossRef] [PubMed]
- Battistini, R.; Rossini, I.; Ercolini, C.; Goria, M.; Callipo, M.R.; Maurella, C.; Pavoni, E.; Serracca, L. Antiviral Activity of Essential Oils Against Hepatitis A Virus in Soft Fruits. Food Environ. Virol. 2019, 11, 90–95. [Google Scholar] [CrossRef]
- Koromyslova, A.D.; White, P.A.; Hansman, G.S. Treatment of norovirus particles with citrate. Virology 2015, 485, 199–204. [Google Scholar] [CrossRef]
- Rosato, A.; Carocci, A.; Catalano, A.; Clodoveo, M.L.; Franchini, C.; Corbo, F.; Carbonara, G.G.; Carrieri, A.; Fracchiolla, G. Elucidation of the synergistic action of Mentha Piperita essential oil with common antimicrobials. PLoS ONE 2018, 13, e0200902. [Google Scholar] [CrossRef]
- Rosato, A.; Maggi, F.; Cianfaglione, K.; Conti, F.; Ciaschetti, G.; Rakotosaona, R.; Fracchiolla, G.; Clodoveo, M.L.; Franchini, C.; Corbo, F. Chemical composition and antibacterial activity of seven uncommon essential oils. J. Essent. Oil Res. 2018, 30, 233–243. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectorscopy, 4th ed.; Allured Pub. Corp: Carol Stream, IL, USA, 2007. [Google Scholar]
- Waseem, R.; Low, K.H. Advanced analytical techniques for the extraction and characterization of plant-derived essential oils by gas chromatography with mass spectrometry. J. Sep. Sci. 2015, 38, 483–501. [Google Scholar] [CrossRef]
- Koo, I.; Kim, S.; Zhang, X. Comparative analysis of mass spectral matching-based compound identification in gas chromatography–mass spectrometry. J. Chromatogr. A 2013, 1298, 132–138. [Google Scholar] [CrossRef]
- Wan, K.X.; Vidavsky, I.; Gross, M.L. Comparing similar spectra: From similarity index to spectral contrast angle. J. Am. Soc. Mass Spectrom. 2002, 13, 85–88. [Google Scholar] [CrossRef]
- Lanave, G.; Cavalli, A.; Martella, V.; Fontana, T.; Losappio, R.; Tempesta, M.; Decaro, N.; Buonavoglia, D.; Camero, M. Ribavirin and boceprevir are able to reduce Canine distemper virus growth in vitro. J. Virol. Methods 2017, 248, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Gilling, D.; Kitajima, M.; Torrey, J.; Bright, K. Antiviral efficacy and mechanisms of action of oregano essential oil and its primary component carvacrol against murine norovirus. J. Appl. Microbiol. 2014, 116, 1149–1163. [Google Scholar] [CrossRef] [PubMed]
- Baert, L.; Debevere, J.; Uyttendaele, M. The efficacy of preservation methods to inactivate foodborne viruses. Int. J. Food Microbiol. 2009, 131, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Butot, S.; Putallaz, T.; Sanchez, G. Effects of sanitation, freezing and frozen storage on enteric viruses in berries and herbs. Int. J. Food Microbiol. 2008, 126, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Tursun, A.O. Impact of soil types on chemical composition of essential oil of purple basil. Saudi J. Biol. Sci. 2022, 29, 103314. [Google Scholar] [CrossRef]
- Ravishankar, S.; Zhu, L.; Reyna-Granados, J.; Law, B.; Joens, L.; Friedman, M. Carvacrol and Cinnamaldehyde Inactivate Antibiotic-Resistant Salmonella enterica in Buffer and on Celery and Oysters. J. Food Prot. 2010, 73, 234–240. [Google Scholar] [CrossRef]
- Mieres-Castro, D.; Ahmar, S.; Shabbir, R.; Mora-Poblete, F. Antiviral Activities of Eucalyptus Essential Oils: Their Effectiveness as Therapeutic Targets against Human Viruses. Pharmaceuticals 2021, 14, 1210. [Google Scholar] [CrossRef]
- Reichling, J. Antiviral and Virucidal Properties of Essential Oils and Isolated Compounds—A Scientific Approach. Planta Med. 2022, 88, 587–603. [Google Scholar] [CrossRef]
- Su, X.; Howell, A.B.; D’Souza, D.H. Antiviral effects of cranberry juice and cranberry proanthocyanidins on foodborne viral surrogates—A time dependence study in vitro. Food Microbiol. 2010, 27, 985–991. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Baert, L.; Zhang, D.; Xia, M.; Zhong, W.; Van Coillie, E.; Jiang, X.; Uyttendaele, M. Effect of Grape Seed Extract on Human Norovirus GII.4 and Murine Norovirus 1 in Viral Suspensions, on Stainless Steel Discs, and in Lettuce Wash Water. Appl. Environ. Microbiol. 2012, 78, 7572–7578. [Google Scholar] [CrossRef] [PubMed]
- Schnitzler, P. Essential Oils for the Treatment of Herpes Simplex Virus Infections. Chemotherapy 2019, 64, 1–7. [Google Scholar] [CrossRef]
- Shogan, B.; Kruse, L.; Mulamba, G.B.; Hu, A.; Coen, D.M. Virucidal Activity of a GT-Rich Oligonucleotide against Herpes Simplex Virus Mediated by Glycoprotein B. J. Virol. 2006, 80, 4740–4747. [Google Scholar] [CrossRef] [PubMed]
- Hofmann-Lehmann, R.; Hosie, M.J.; Hartmann, K.; Egberink, H.; Truyen, U.; Tasker, S.; Belák, S.; Boucraut-Baralon, C.; Frymus, T.; Lloret, A.; et al. Calicivirus Infection in Cats. Viruses 2022, 14, 937. [Google Scholar] [CrossRef]
- Chiu, S.; Skura, B.; Petric, M.; McIntyre, L.; Gamage, B.; Isaac-Renton, J. Efficacy of common disinfectant/cleaning agents in inactivating murine norovirus and feline calicivirus as surrogate viruses for human norovirus. Am. J. Infect. Control. 2015, 43, 1208–1212. [Google Scholar] [CrossRef]
- Barker, J.; Vipond, I.; Bloomfield, S. Effects of cleaning and disinfection in reducing the spread of Norovirus contamination via environmental surfaces. J. Hosp. Infect. 2004, 58, 42–49. [Google Scholar] [CrossRef]
- Said, M.A.; Perl, T.M.; Sears, C.L. Healthcare Epidemiology: Gastrointestinal Flu: Norovirus in Health Care and Long-Term Care Facilities. Clin. Infect. Dis. 2008, 47, 1202–1208. [Google Scholar] [CrossRef]
- Williamson, E.M. Aromatherapy Science: Maria Lis-Balchin. Publisher: Pharmaceutical Press. UK, pp 496. Phytother. Res. 2006, 20, 333. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Solis-Sanchez, D.; Rivera-Piza, A.; Lee, S.; Kim, J.; Kim, B.; Choi, J.B.; Kim, Y.W.; Ko, G.P.; Song, M.J.; Lee, S.-J. Antiviral Effects of Lindera obtusiloba Leaf Extract on Murine Norovirus-1 (MNV-1), a Human Norovirus Surrogate, and Potential Application to Model Foods. Antibiotics 2020, 9, 697. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| N | Components | LRI | AI | Citrus lemon | |
|---|---|---|---|---|---|
| Area ± SEM | SI/MS | ||||
| 1 | Ethyl propanoate | 714 | 714 | 0.1 ± 0.010 | 91 |
| 2 | α-pinene a | 930 | 931 | 2.4 ± 0.5 | 95 |
| 3 | β-thujene | 968 | 968 | 1.94 ± 0.2 | 86 |
| 4 | β-pinene a | 982 | 980 | 14.5 ± 1 | 94 |
| 5 | Limonene a | 1030 | 1032 | 53 ± 5 | 93 |
| 6 | γ-terpinene a | 1062 | 1064 | 5.9 ± 1 | 94 |
| 7 | terpinolene | 1083 | 1085 | 0.2 ± 0.020 | 96 |
| 8 | β-linaloola | 1100 | 1101 | 0.2 ± 0.020 | 91 |
| 9 | (E)-p-menth-2,8-dien-1-ol | 1122 | 1123 | 0.13 ± 0.02 | 80 |
| 10 | limonene oxide, cis- | 1130 | 1131 | 1 ± 0.3 | 96 |
| 11 | limonene oxide, trans- | 1138 | 1138 | 0.7 ± 0.08 | 91 |
| 12 | α-terpineol | 1178 | 1179 | 0.3 ± 0.020 | 80 |
| 13 | cis-carveol | 1222 | 1222 | 0.3 ± 0.020 | 96 |
| 14 | citral a | 1240 | 1240 | 3.8 ± 0.9 | 96 |
| 15 | Δ-carvone | 1242 | 1242 | 0.15 ± 0.01 | 93 |
| 16 | nerol acetate | 1363 | 1364 | 0.8 ± 0.05 | 91 |
| 17 | geranyl aceate | 1384 | 1385 | 0.9 ± 0.06 | 91 |
| 18 | Caryophyllene a | 1415 | 1415 | 0.15 ± 0.01 | 99 |
| 19 | α-bergamotene | 1431 | 1430 | 0.21 ± 0.02 | 87 |
| 20 | β-bisabolene a | 1504 | 1506 | 0.56 ± 0.04 | 95 |
| 21 | caryophylleneoxyde | 1596 | 1592 | 0.57 ± 0.05 | 91 |
| % Characterized | / | / | 87.81 | / | |
| Others | / | / | 12.19 | / | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pellegrini, F.; Camero, M.; Catella, C.; Fracchiolla, G.; Sblano, S.; Patruno, G.; Trombetta, C.M.; Galgano, M.; Pratelli, A.; Tempesta, M.; et al. Virucidal Activity of Lemon Essential Oil against Feline Calicivirus Used as Surrogate for Norovirus. Antibiotics 2023, 12, 322. https://doi.org/10.3390/antibiotics12020322
Pellegrini F, Camero M, Catella C, Fracchiolla G, Sblano S, Patruno G, Trombetta CM, Galgano M, Pratelli A, Tempesta M, et al. Virucidal Activity of Lemon Essential Oil against Feline Calicivirus Used as Surrogate for Norovirus. Antibiotics. 2023; 12(2):322. https://doi.org/10.3390/antibiotics12020322
Chicago/Turabian StylePellegrini, Francesco, Michele Camero, Cristiana Catella, Giuseppe Fracchiolla, Sabina Sblano, Giovanni Patruno, Claudia Maria Trombetta, Michela Galgano, Annamaria Pratelli, Maria Tempesta, and et al. 2023. "Virucidal Activity of Lemon Essential Oil against Feline Calicivirus Used as Surrogate for Norovirus" Antibiotics 12, no. 2: 322. https://doi.org/10.3390/antibiotics12020322