
An In Vitro and In Silico Investigation about Monteverdia ilicifolia Activity against Helicobacter pylori

 , , , , and
, , , , and
Abstract
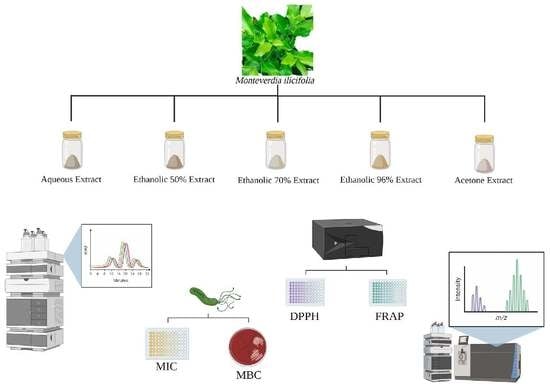
:
1. Introduction
2. Results and Discussion
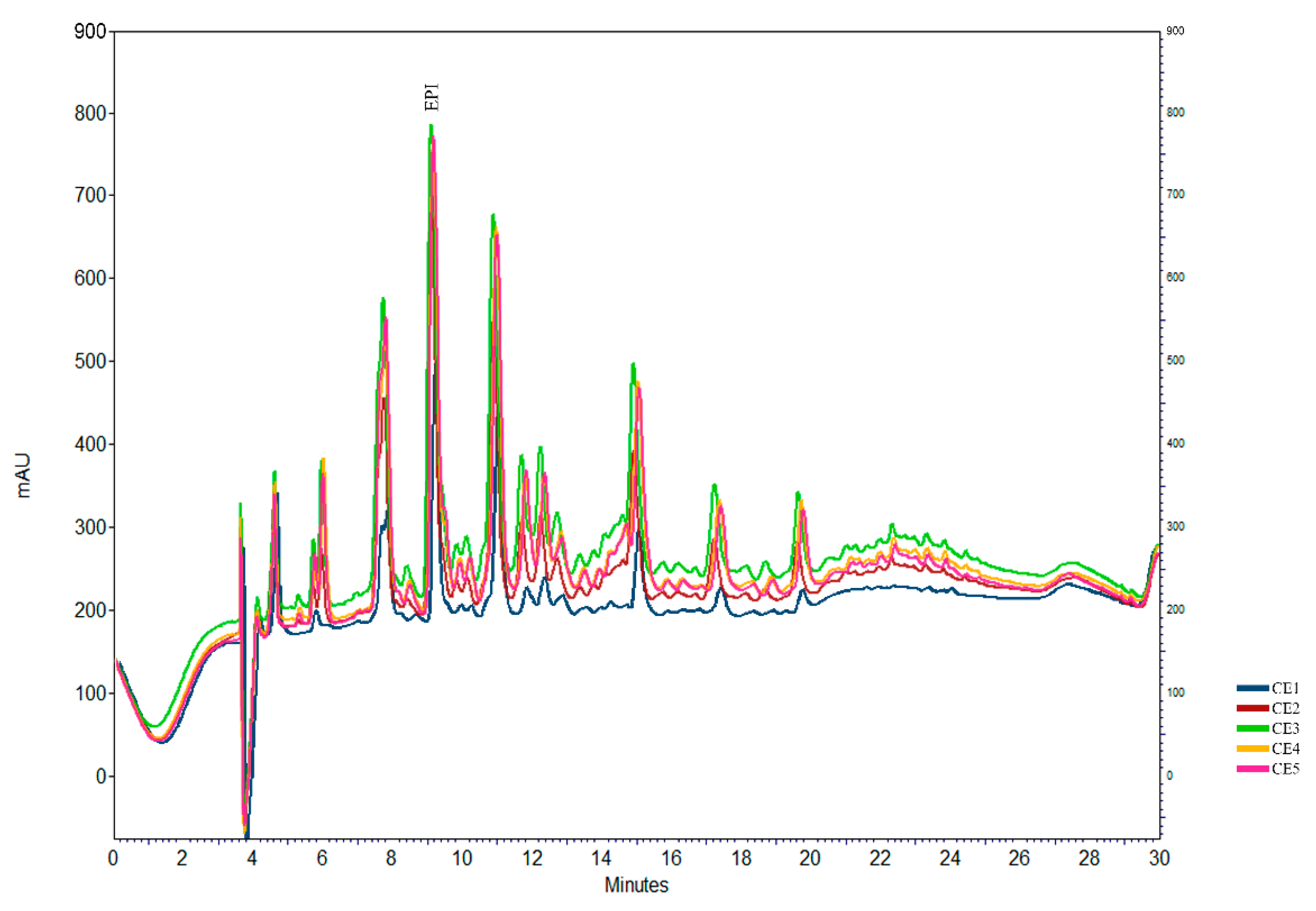
2.1. Epicatechin Determination Using HPLC
2.2. Antibacterial Activities of Extracts and Semi-Purified Fractions against H. pylori
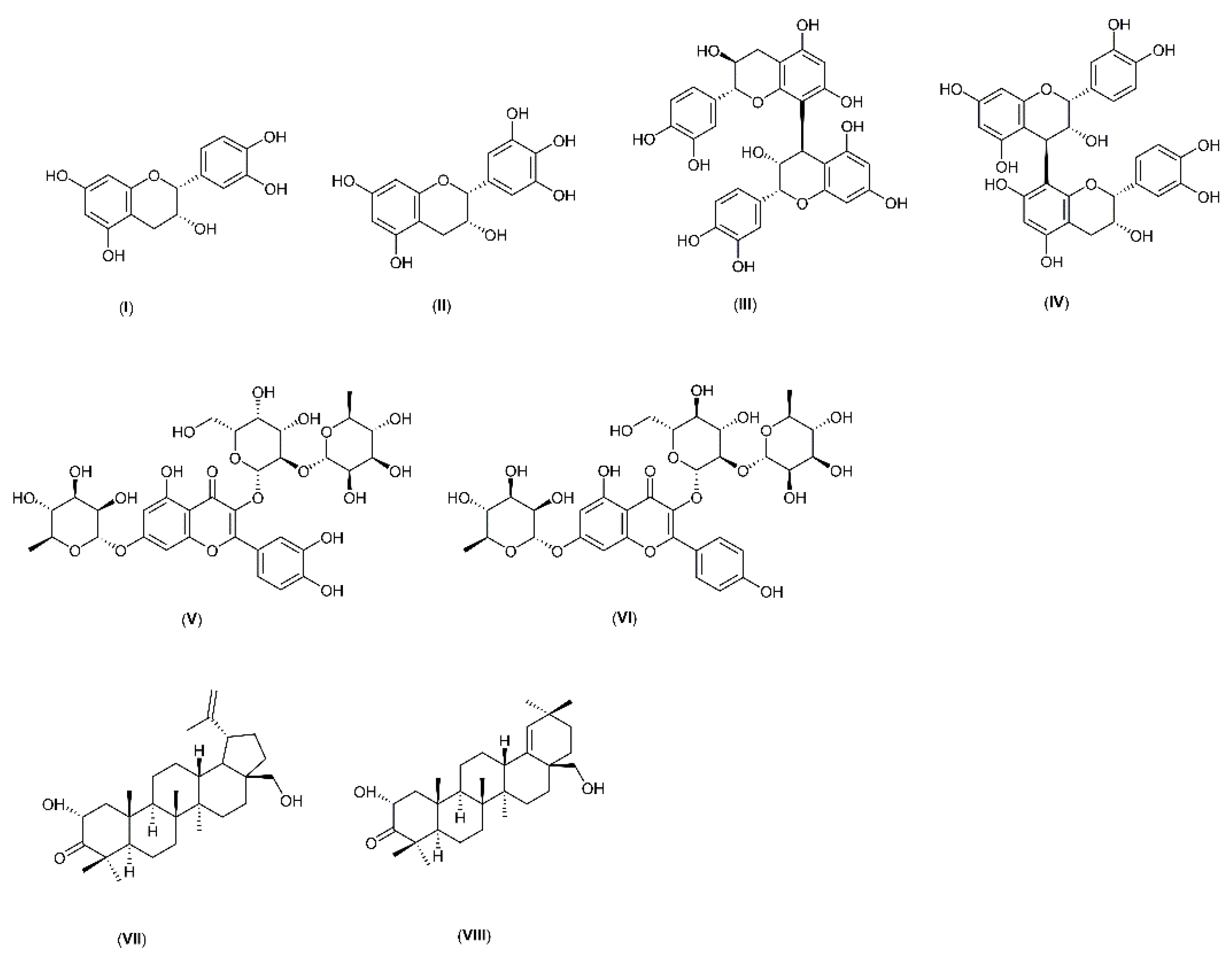
2.3. UPLC-MS Profiles of M. ilicifolia Extracts
2.4. Antioxidant Capacity of Extracts and Semi-Purified Fractions
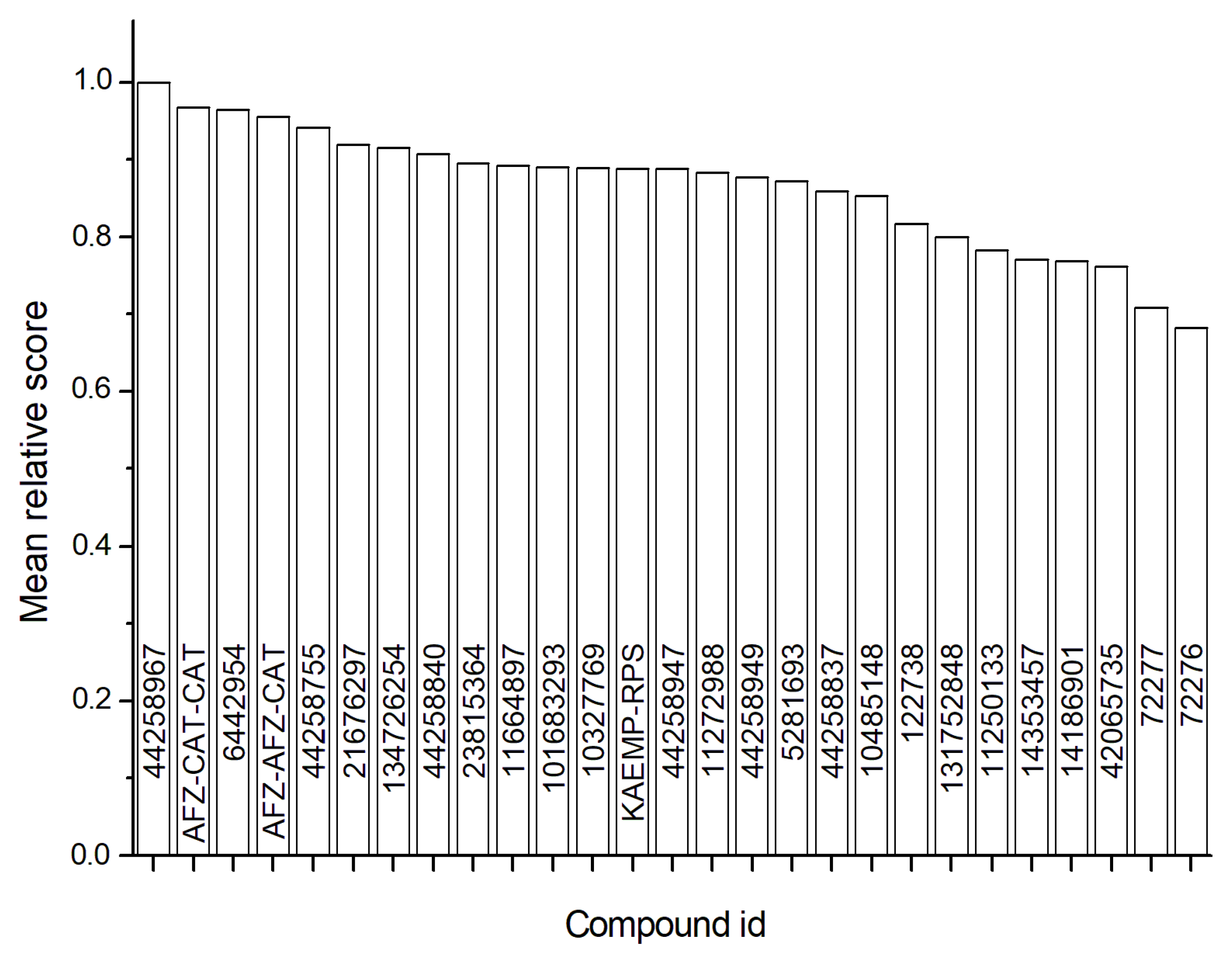
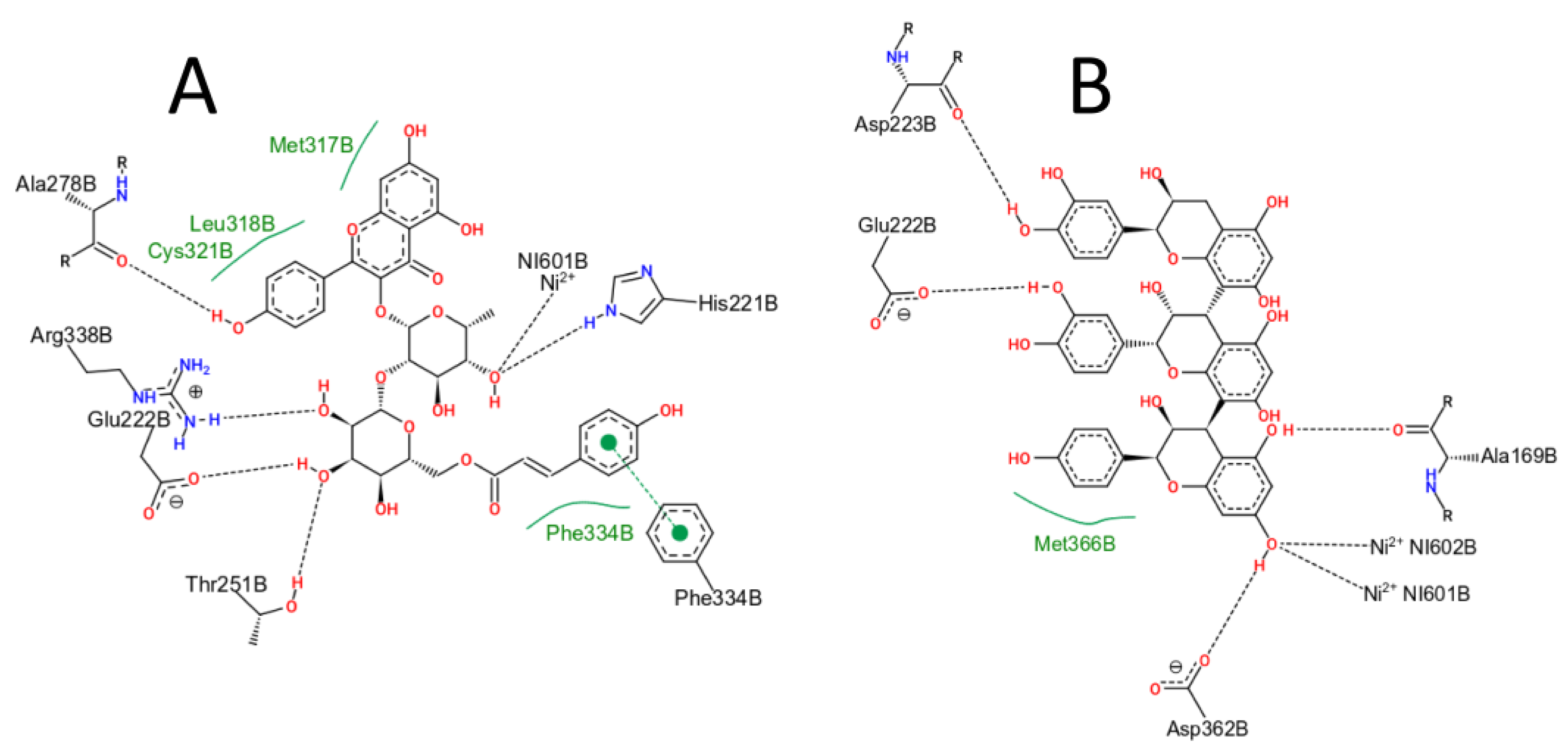
2.5. Virtual Screening
3. Materials and Methods
3.1. Plant Material
3.2. Extracts and Fractions Preparation
3.3. Epicatechin Determination Using HPLC Method
3.4. Bacteria Strain
3.4.1. Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
3.4.2. Urease Inhibition Assay
3.5. UHPLC-MS Conditions
3.6. Antioxidant Capacity Assay
3.6.1. Determination of DPPH (2,2-diphenyl-1-picrylhydrazyl) Radical Levels
3.6.2. Ferric Reducing Antioxidant Power (FRAP) Assay
3.7. Virtual Screening
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CE1 | Crude extract aqueous |
| EAF1 | Ethyl-acetate fraction from crude extract aqueous |
| nBF1 | n-Butanolic fraction from crude extract aqueous |
| AQF1 | Aqueous fraction from crude extract aqueous |
| CE2 | Crude extract ethanol: water 50:50 (v/v) |
| EAF2 | Ethyl-acetate fraction from crude extract ethanol: water 50:50 (v/v) |
| nBF2 | n-Butanolic fraction from crude extract ethanol: water 50:50 (v/v) |
| AQF2 | Aqueous fraction from crude extract ethanol: water 50:50 (v/v) |
| CE3 | Crude extract ethanol: water 70:30 (v/v) |
| EAF3 | Ethyl-acetate fraction from crude extract ethanol: water 70:30 (v/v) |
| nBF3 | n-Butanolic fraction from crude extract ethanol: water 70:30 (v/v) |
| AQF3 | Aqueous fraction from crude extract ethanol: water 70:30 (v/v) |
| CE4 | Crude extract ethanol: water 96:4 (v/v) |
| EAF4 | Ethyl-acetate fraction from crude extract ethanol: water 96:4 (v/v) |
| nBF4 | n-Butanolic fraction from crude extract ethanol: water 96:4 (v/v) |
| AQF4 | Aqueous fraction from crude extract ethanol: water 96:4 (v/v) |
| CE5 | Crude extract acetone: water 70:30 (v/v) |
| EAF5 | Ethyl-acetate fraction from crude extract acetone: water 70:30 (v/v) |
| nBF5 | n-Butanolic fraction from crude extract acetone: water 70:30 (v/v) |
| AQF5 | Aqueous fraction from crude extract acetone: water 70:30 (v/v) |
References
- Wroblewski, L.E.; Peek, R.M.; Wilson, K.T. Helicobacter pylori and gastric cancer: Factors that modulate disease risk. Clin. Microbiol. Rev. 2010, 23, 713–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kao, C.-Y.; Sheu, B.-S.; Wu, J.-J. Helicobacter pylori infection: An overview of bacterial virulence factors and pathogenesis. Biomed. J. 2016, 39, 14–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooi, J.K.; Lai, W.Y.; Ng, W.K.; Suen, M.M.; Underwood, F.E.; Tanyingoh, D.; Malfertheiner, P.; Graham, D.Y.; Wong, V.W.; Wu, J.C. Global Prevalence of Helicobacter pylori Infection: Systematic Review and Meta-analysis. Gastroenterology 2017, 153, 420–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, L.G.V.; Marinho, J.R.; Genta, R.; Ribeiro, L.T.; Passos, M.d.C.F.; Zaterka, S.; Assumpção, P.P.; Barbosa, A.J.A.; Barbuti, R.; Braga, L.L.J.A.d.g. IVth Brazilian Consensus Conference on Helicobacter pylori Infection. Arq. Gastroenterol. 2018, 55, 97–121. [Google Scholar] [CrossRef] [PubMed]
- Hunt, R.; Xiao, S.; Megraud, F.; Leon-Barua, R.; Bazzoli, F.; Van der Merwe, S.; Vaz Coelho, L.; Fock, M.; Fedail, S.; Cohen, H. Helicobacter pylori in developing countries. World gastroenterology organisation global guideline. J. Gastrointest. Liver Dis. 2011, 20, 299–304. [Google Scholar]
- Kusters, J.G.; van Vliet, A.H.; Kuipers, E.J. Pathogenesis of Helicobacter pylori infection. Clin. Microbiol. Rev. 2006, 19, 449–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorge, R.; Leite, J.; Oliveira, A.; Tagliati, C. Evaluation of antinociceptive, anti-inflammatory and antiulcerogenic activities of Maytenus ilicifolia. J. Ethnopharmacol. 2004, 94, 93–100. [Google Scholar] [CrossRef]
- Biral, L.; Simmons, M.P.; Smidt, E.C.; Tembrock, L.R.; Bolson, M.; Archer, R.H.; Lombardi, J.A. Systematics of New World Maytenus (Celastraceae) and a New Delimitation of the Genus. Syst. Bot. 2017, 42, 680–693. [Google Scholar] [CrossRef]
- Mazza, M.; dos Santos, J.; Mazza, C.d.S. Fenologia reprodutiva de Maytenus ilicifolia (Celastraceae) na Floresta Nacional de Irati, Paraná, Brasil. Braz. J. Bot. 2011, 34, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Souza-Formigoni, M.L.O.; Oliveira, M.G.M.; Monteiro, M.G.; da Silveira-Filho, N.G.; Braz, S.; Carlini, E. Antiulcerogenic effects of two Maytenus species in laboratory animals. J. Ethnopharmacol. 1991, 34, 21–27. [Google Scholar] [CrossRef]
- Pessuto, M.B.; Costa, I.C.d.; Souza, A.B.d.; Nicoli, F.M.; Petereit, F.; Luftmann, H.; Mello, J.C.P. Atividade antioxidante de extratos e taninos condensados das folhas de Maytenus ilicifolia Mart. ex Reiss. Química Nova 2009, 32, 412–416. [Google Scholar] [CrossRef]
- Gullo, F.P.; Sardi, J.C.; Santos, V.A.; Sangalli-Leite, F.; Pitangui, N.S.; Rossi, S.A.; de Paula e Silva, A.C.; Soares, L.A.; Silva, J.F.; Oliveira, H.C. Antifungal activity of maytenin and pristimerin. Evid.-Based Complement. Altern. Med. 2012, 2012, 340787. [Google Scholar] [CrossRef] [Green Version]
- Araújo Júnior, R.F.d.; Oliveira, A.L.C.d.S.L.; Pessoa, J.B.; Garcia, V.B.; Guerra, G.C.B.; Soares, L.A.L.; de Souza, T.P.; Petrovick, P.R.; de Araújo, A.A. Maytenus ilicifolia dry extract protects normal cells, induces apoptosis and regulates Bcl-2 in human cancer cells. Exp. Biol. Med. 2013, 238, 1251–1258. [Google Scholar] [CrossRef]
- Dos Santos, V.A.; Leite, K.M.; da Costa Siqueira, M.; Regasini, L.O.; Martinez, I.; Nogueira, C.T.; Galuppo, M.K.; Stolf, B.S.; Pereira, A.M.S.; Cicarelli, R. Antiprotozoal activity of quinonemethide triterpenes from Maytenus ilicifolia (Celastraceae). Molecules 2013, 18, 1053–1062. [Google Scholar] [CrossRef]
- Wonfor, R.; Natoli, M.; Parveen, I.; Beckman, M.; Nash, R.; Nash, D. Anti-inflammatory properties of an extract of M. ilicifolia in the human intestinal epithelial Caco-2 cell line. J. Ethnopharmacol. 2017, 209, 283–287. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, N.; Kashiwada, Y.J.J.o.n.m. Characteristic metabolites of Hypericum plants: Their chemical structures and biological activities. J. Nat. Med. 2021, 75, 423–433. [Google Scholar] [CrossRef]
- Taguri, T.; Tanaka, T.; Kouno, I. Antimicrobial activity of 10 different plant polyphenols against bacteria causing food-borne disease. Biol. Pharm. Bull. 2004, 27, 1965–1969. [Google Scholar] [CrossRef] [Green Version]
- Daglia, M. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef]
- Anvisa. Farmacopeia Brasileira, 5th ed.; Agência Nacional de Vigilância Sanitária: Brasília, Brazil; Volume 2, pp. 922–927.
- Wang, Y.C.; Huang, T.L. Anti-Helicobacter pylori activity of Plumbago zeylanica L. Pathog. Dis. 2005, 43, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Njume, C.; Afolayan, A.J.; Ndip, R.N. Preliminary phytochemical screening and in vitro anti-Helicobacter pylori activity of acetone and aqueous extracts of the stem bark of Sclerocarya birrea (Anacardiaceae). Arch. Med. Res. 2011, 42, 252–257. [Google Scholar] [CrossRef]
- Santos, S.d.C.; Mello, J.C.P.d. Taninos. In Farmacognosia: Da planta ao medicamento; Simões, C.M.O., Schenkel, E.P., Gosmann, G., Mello, J.C.P.d., Mentz, L.A., Petrovick, P.R., Eds.; Editora da UFRGS; Editora da UFSC: Porto Alegre, Florianópolis, 2007; pp. 615–656. [Google Scholar]
- Park, D.; Shin, K.; Choi, Y.; Guo, H.; Cha, Y.; Kim, S.-H.; Han, N.S.; Joo, S.S.; Choi, J.K.; Lee, Y.B. Antimicrobial activities of ethanol and butanol fractions of white rose petal extract. Regul. Toxicol. Pharmacol. 2016, 76, 57–62. [Google Scholar] [CrossRef]
- Bae, E.-A.; Han, M.J.; Kim, N.-J.; Kim, D.-H. Anti-Helicobacter pylori Activity of Hearbal Medicines. Biol. Pharm. Bull. 1998, 21, 990–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Z.-P.; Shi, D.-H.; Li, H.-Q.; Zhang, L.-N.; Xu, C.; Zhu, H.-L. Polyphenols based on isoflavones as inhibitors of Helicobacter pylori urease. Bioorganic Med. Chem. 2007, 15, 3703–3710. [Google Scholar] [CrossRef] [PubMed]
- Ohsaki, A.; Imai, Y.; Naruse, M.; Ayabe, S.; Komiyama, K.; Takashima, J. Four new triterpenoids from Maytenus ilicifolia. J. Nat. Prod. 2004, 67, 469–471. [Google Scholar] [CrossRef]
- Leite, J.P.V.; Rastrelli, L.; Romussi, G.; Oliveira, A.B.; Vilegas, J.H.; Vilegas, W.; Pizza, C. Isolation and HPLC quantitative analysis of flavonoid glycosides from Brazilian beverages (Maytenus ilicifolia and M. aquifolium). J. Agric. Food Chem. 2001, 49, 3796–3801. [Google Scholar] [CrossRef] [PubMed]
- Leite, J.P.V.; Braga, F.C.; Romussi, G.; Persoli, R.M.; Tabach, R.; Carlini, E.A.; Oliveira, A.B. Constituents from Maytenus ilicifolia leaves and bioguided fractionation for gastroprotective activity. J. Braz. Chem. Soc. 2010, 21, 248–254. [Google Scholar] [CrossRef] [Green Version]
- Lopes, G.C.; Blainski, A.; Santos, P.V.P.d.; Diciaula, M.C.; Mello, J.C.P.d. Development and validation of an HPLC method for the determination of epicatechin in Maytenus ilicifolia (Schrad.) Planch., Celastraceae. Rev. Bras. De Farmacogn. 2010, 20, 781–788. [Google Scholar] [CrossRef] [Green Version]
- Sannomiya, M.; Vilegas, W.; Rastrelli, L.; Pizza, C. A flavonoid glycoside from Maytenus aquifolium. Phytochemistry 1998, 49, 237–239. [Google Scholar] [CrossRef]
- Vilegas, W.; Sanommiya, M.; Rastrelli, L.; Pizza, C. Isolation and structure elucidation of two new flavonoid glycosides from the infusion of Maytenus aquifolium leaves. Evaluation of the antiulcer activity of the infusion. J. Agric. Food Chem. 1999, 47, 403–406. [Google Scholar] [CrossRef]
- Silvan, J.M.; Gutiérrez-Docio, A.; Moreno-Fernandez, S.; Alarcón-Cavero, T.; Prodanov, M.; Martinez-Rodriguez, A.J.J.F. Procyanidin-rich extract from grape seeds as a putative tool against Helicobacter pylori. Foods 2020, 9, 1370. [Google Scholar] [CrossRef]
- Pastene, E.; Parada, V.; Avello, M.; Ruiz, A.; Garcia, A. Catechin-based procyanidins from Peumus boldus Mol. aqueous extract inhibit Helicobacter pylori urease and adherence to adenocarcinoma gastric cells. Phytother. Res. 2014, 28, 1637–1645. [Google Scholar] [CrossRef] [PubMed]
- Mabe, K.; Yamada, M.; Oguni, I.; Takahashi, T. In vitro and in vivo activities of tea catechins against Helicobacter pylori. Antimicrob. Agents Chemother. 1999, 43, 1788–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-Gómez, R.; López-Solís, R.; Obreque-Slier, E.; Toledo-Araya, H. Comparative antibacterial effect of gallic acid and catechin against Helicobacter pylori. LWT Food Sci. Technol. 2013, 54, 331–335. [Google Scholar] [CrossRef]
- Yanagawa, Y.; Yamamoto, Y.; Hara, Y.; Shimamura, T. A combination effect of epigallocatechin gallate, a major compound of green tea catechins, with antibiotics on Helicobacter pylori growth in vitro. Curr. Microbiol. 2003, 47, 244–249. [Google Scholar] [CrossRef]
- Shabbir, M.; Khan, M.R.; Saeed, N. Assessment of phytochemicals, antioxidant, anti-lipid peroxidation and anti-hemolytic activity of extract and various fractions of Maytenus royleanus leaves. BMC Complement. Altern. Med. 2013, 13, 143. [Google Scholar] [CrossRef] [Green Version]
- Soobrattee, M.A.; Bahorun, T.; Neergheen, V.S.; Googoolye, K.; Aruoma, O.I. Assessment of the content of phenolics and antioxidant actions of the Rubiaceae, Ebenaceae, Celastraceae, Erythroxylaceae and Sterculaceae families of Mauritian endemic plants. Toxicol. Vitr. 2008, 22, 45–56. [Google Scholar] [CrossRef]
- Resende, F.O.; Rodrigues-Filho, E.; Luftmann, H.; Petereit, F.; Mello, J.C.J.J.o.t.B.C.S. Phenylpropanoid substituted flavan-3-ols from Trichilia catigua and their in vitro antioxidative activity. J. Braz. Chem. Soc. 2011, 22, 2087–2093. [Google Scholar] [CrossRef] [Green Version]
- Van Acker, S.A.; Tromp, M.N.; Griffioen, D.H.; Van Bennekom, W.P.; Van Der Vijgh, W.J.; Bast, A. Structural aspects of antioxidant activity of flavonoids. Free Radic. Biol. Med. 1996, 20, 331–342. [Google Scholar] [CrossRef]
- Stierand, K.; Maaß, P.C.; Rarey, M.J.B. Molecular complexes at a glance: Automated generation of two-dimensional complex diagrams. Bioinformatics 2006, 22, 1710–1716. [Google Scholar] [CrossRef] [Green Version]
- de Paula, M.N.; Kelm, M.; Symma, N.; Isolani, R.G.; da Silva, F.P.; Sendker, J.; Hensel, A.; de Mello, J.C.P.J.R.B.d.F. Anti-adhesive Activity of Maytenus ilicifolia Against Helicobacter pylori. Braz. J. Pharmacogn. 2021, 31, 726–731. [Google Scholar] [CrossRef]
- Chierrito, D.; Villas-Boas, C.B.; Tonin, F.S.; Fernandez-Llimos, F.; Sanches, A.C.; de Mello, J.C.J.C.n. Using cell cultures for the investigation of treatments for attention deficit hyperactivity disorder: A systematic review. Curr. Neuropharmacol. 2019, 17, 916–925. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.S.; Minozzo, B.R.; Santos, H.; Santa Ardisson, J.; Rodrigues, R.P.; Romão, W.; de Souza Borges, W.; Gonçalves, R.d.C.R.; Beltrame, F.L.; Kitagawa, R.R. Plectranthus barbatus Andrews as anti-Helicobacter pylori agent with activity against adenocarcinoma gastric cells. Ind. Crops Prod. 2020, 146, 112207. [Google Scholar] [CrossRef]
- Sereia, A.L.; de Oliveira, M.T.; Baranoski, A.; Marques, L.L.M.; Ribeiro, F.M.; Isolani, R.G.; de Medeiros, D.C.; Chierrito, D.; Lazarin-Bidóia, D.; Zielinski, A.A.F. In vitro evaluation of the protective effects of plant extracts against amyloid-beta peptide-induced toxicity in human neuroblastoma SH-SY5Y cells. PLoS ONE 2019, 14, e0212089. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comp. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rappé, A.K.; Casewit, C.J.; Colwell, K.; Goddard III, W.A.; Skiff, W.M.J.J.o.t.A.c.s. UFF, a full periodic table force field for molecular mechanics and molecular dynamics simulations. J. Am. Chem. Soc. 1992, 114, 10024–10035. [Google Scholar] [CrossRef]
- Jones, G.; Willett, P.; Glen, R.C.; Leach, A.R.; Taylor, R.J.J.o.m.b. Development and validation of a genetic algorithm for flexible docking. J. Mol. Biol. 1997, 267, 727–748. [Google Scholar] [CrossRef] [Green Version]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R.J.J.o.c. Open Babel: An open chemical toolbox. J. Cheminform. 2011, 3, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V.J.S.r. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MIC50 (µg/mL ± SD) | CE1 | CE2 | CE3 | CE4 | CE5 |
|---|---|---|---|---|---|
| CE | 1024 ± 2.1 | 1024 ± 5.1 | 512 ± 5.6 | 512 ± 2.2 | 64 ± 9.8 |
| EAF | >1024 ± 17.3 | 64 ± 5.4 | 256 ± 1.3 | 256 ± 3.5 | 64 ± 16.1 |
| nBF | >1024 ± 4.7 | 256 ± 1.3 | 1024 ± 2.7 | 1024 ± 3.2 | 128 ± 4.0 |
| AQF | >1024 ± 4.5 | >1024 ± 0.7 | 512 ± 1.9 | 512 ± 5.1 | 256 ± 8.7 |
| Identification | Retention Time (Min) | [M-H]- (m/z) | Main Fragments |
|---|---|---|---|
| (epi)gallocatechin | 8.83 | 305 | 109, 125, 139, 165, 219, 237, 261 |
| procyanidin B2 | 10.63 | 577 | 125, 151, 245, 289, 407, 425, 451 |
| (epi)catechin | 12.29 | 289 | 109, 179, 203, 245 |
| (epi)afzelechin-(epi)catechin | 13.70 | 561 | 125, 289, 435 |
| (epi)afzelechin-(epi)catechin-(epi)catechin | 14.77 | 849 | 125, 289, 407, 559, 679 |
| (epi)afzelechin-(epi)afzelechin-(epi)catechin | 15.31 | 833 | 125, 239, 407, 543 |
| kaempferol-galactoside-rhamnoside-rhamnoside | 15.51 | 739 | 284 |
| (epi)catechin-(epi)catechin | 17.17 | 577 | 125, 161, 289, 407 |
| (epi)afzelechin-(epi)catechin | 21.31 | 561 | 289, 435 |
| kaempferol-rhamnopentoside | 21.71 | 563 | 284 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Paula, M.N.; Ribeiro, T.D.V.R.; Isolani, R.; de Medeiros Araújo, D.C.; Borges, A.S.; Philippsen, G.S.; de Cássia Ribeiro Gonçalves, R.; Kitagawa, R.R.; Seixas, F.A.V.; de Mello, J.C.P. An In Vitro and In Silico Investigation about Monteverdia ilicifolia Activity against Helicobacter pylori. Antibiotics 2023, 12, 46. https://doi.org/10.3390/antibiotics12010046
de Paula MN, Ribeiro TDVR, Isolani R, de Medeiros Araújo DC, Borges AS, Philippsen GS, de Cássia Ribeiro Gonçalves R, Kitagawa RR, Seixas FAV, de Mello JCP. An In Vitro and In Silico Investigation about Monteverdia ilicifolia Activity against Helicobacter pylori. Antibiotics. 2023; 12(1):46. https://doi.org/10.3390/antibiotics12010046
Chicago/Turabian Stylede Paula, Mariana Nascimento, Taísa Dalla Valle Rörig Ribeiro, Raquel Isolani, Daniela Cristina de Medeiros Araújo, Augusto Santos Borges, Gisele Strieder Philippsen, Rita de Cássia Ribeiro Gonçalves, Rodrigo Rezende Kitagawa, Flavio Augusto Vicente Seixas, and João Carlos Palazzo de Mello. 2023. "An In Vitro and In Silico Investigation about Monteverdia ilicifolia Activity against Helicobacter pylori" Antibiotics 12, no. 1: 46. https://doi.org/10.3390/antibiotics12010046