
Effects of Dietary Coated Lysozyme on the Growth Performance, Antioxidant Activity, Immunity and Gut Health of Weaned Piglets
 , ,
, ,
Abstract
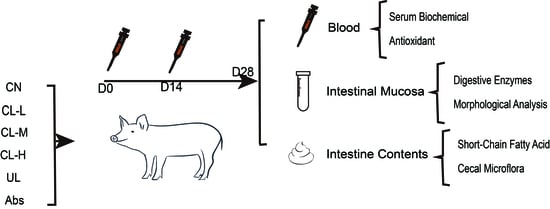
:
1. Introduction
2. Results
2.1. Growth Performance
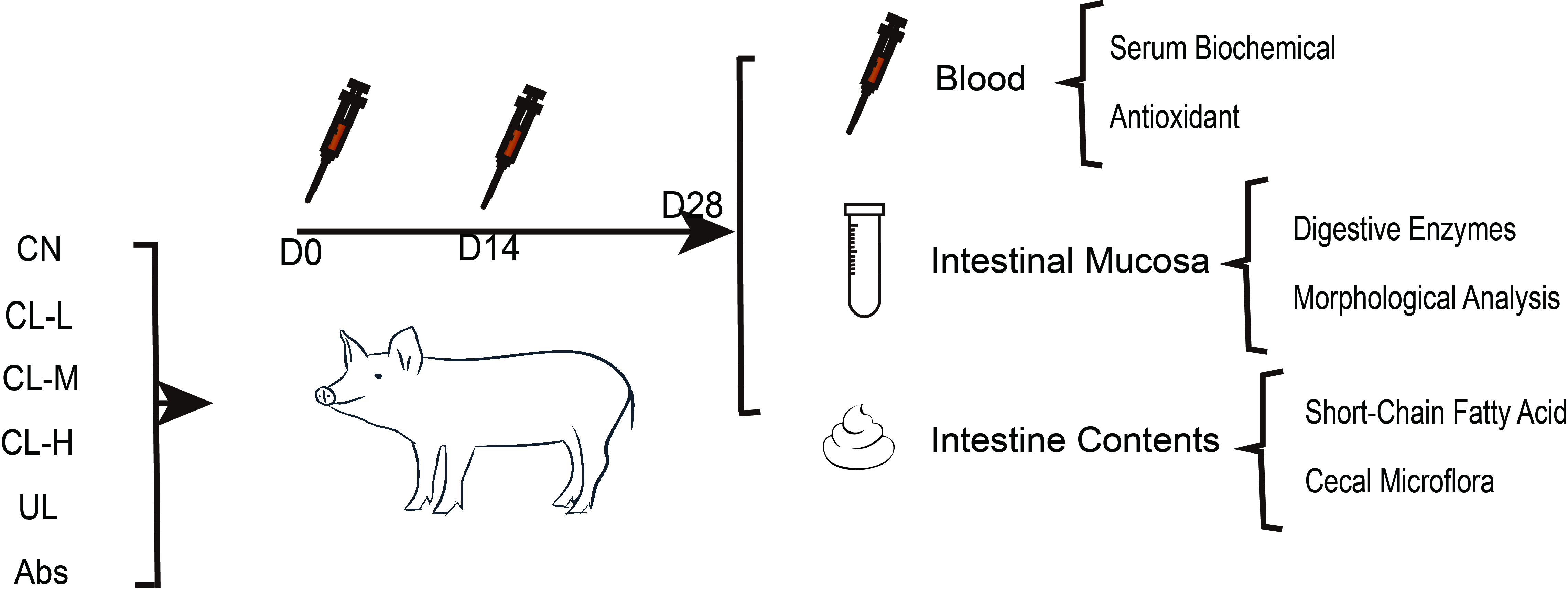
2.2. Serum Biochemical Index Analysis
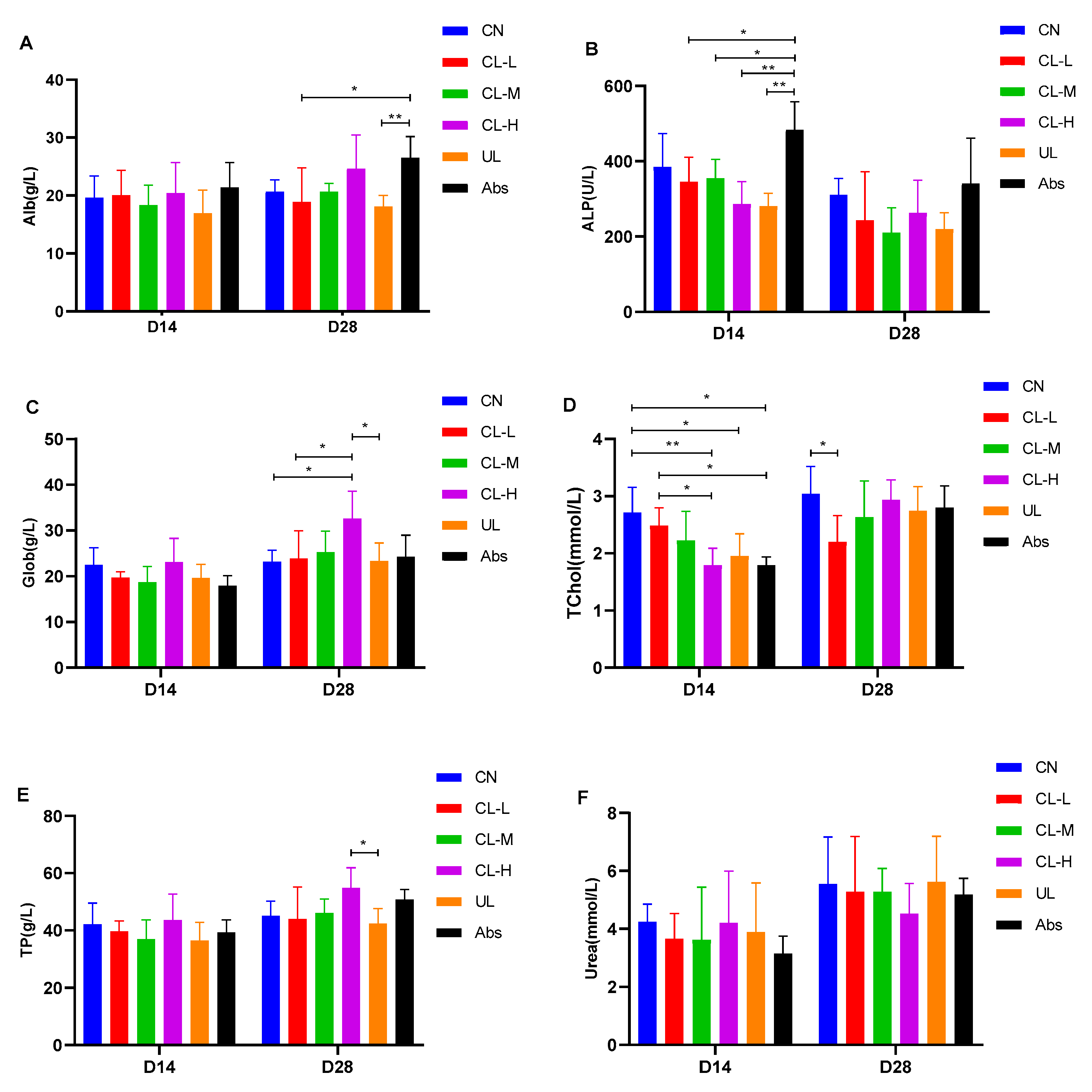
2.3. Immunoglobulin
2.4. Antioxidant Indexes
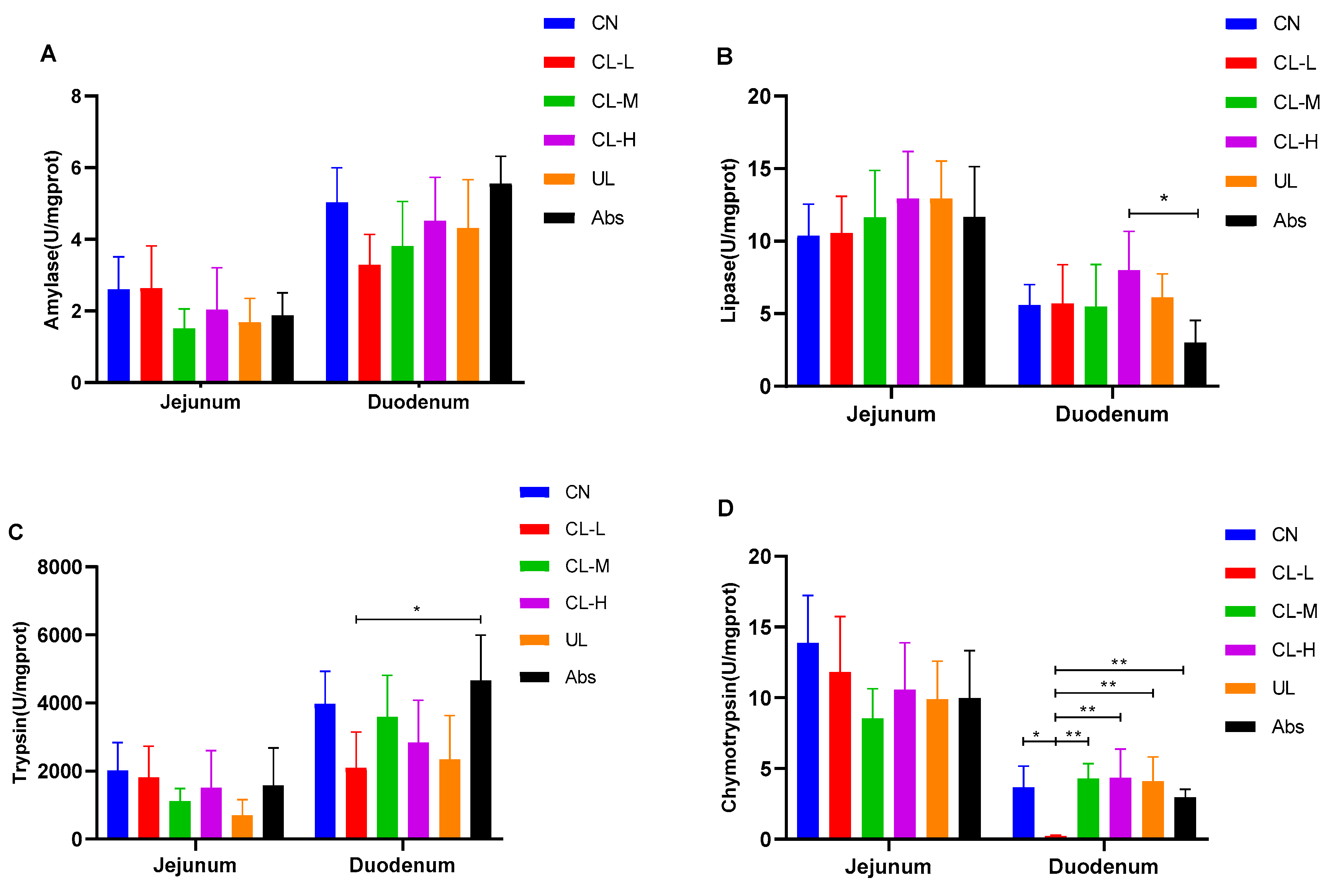
2.5. Digestive Enzymes
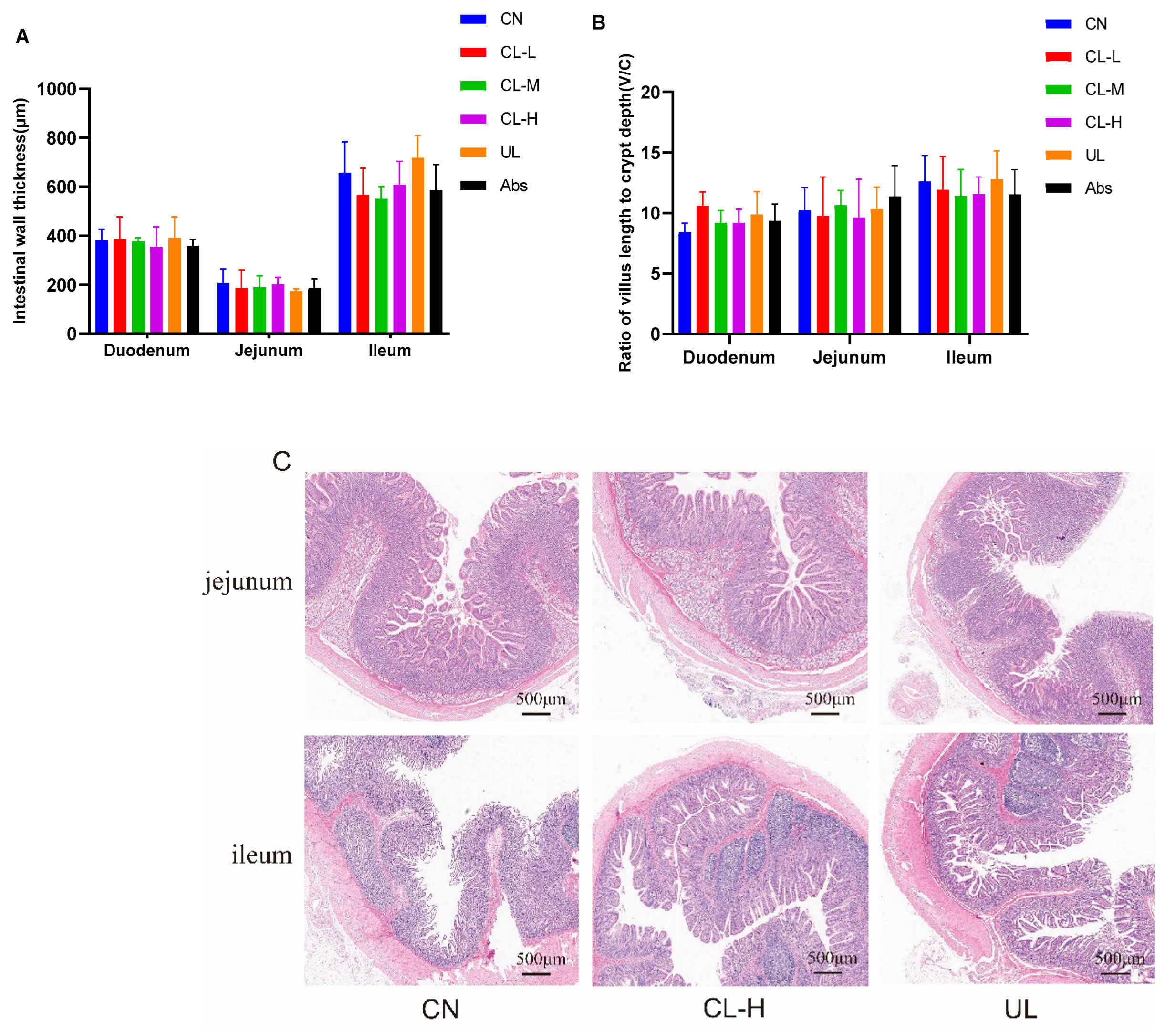
2.6. Morphological Analysis of the Small Intestine
2.7. Quantification of Short-Chain Fatty Acids
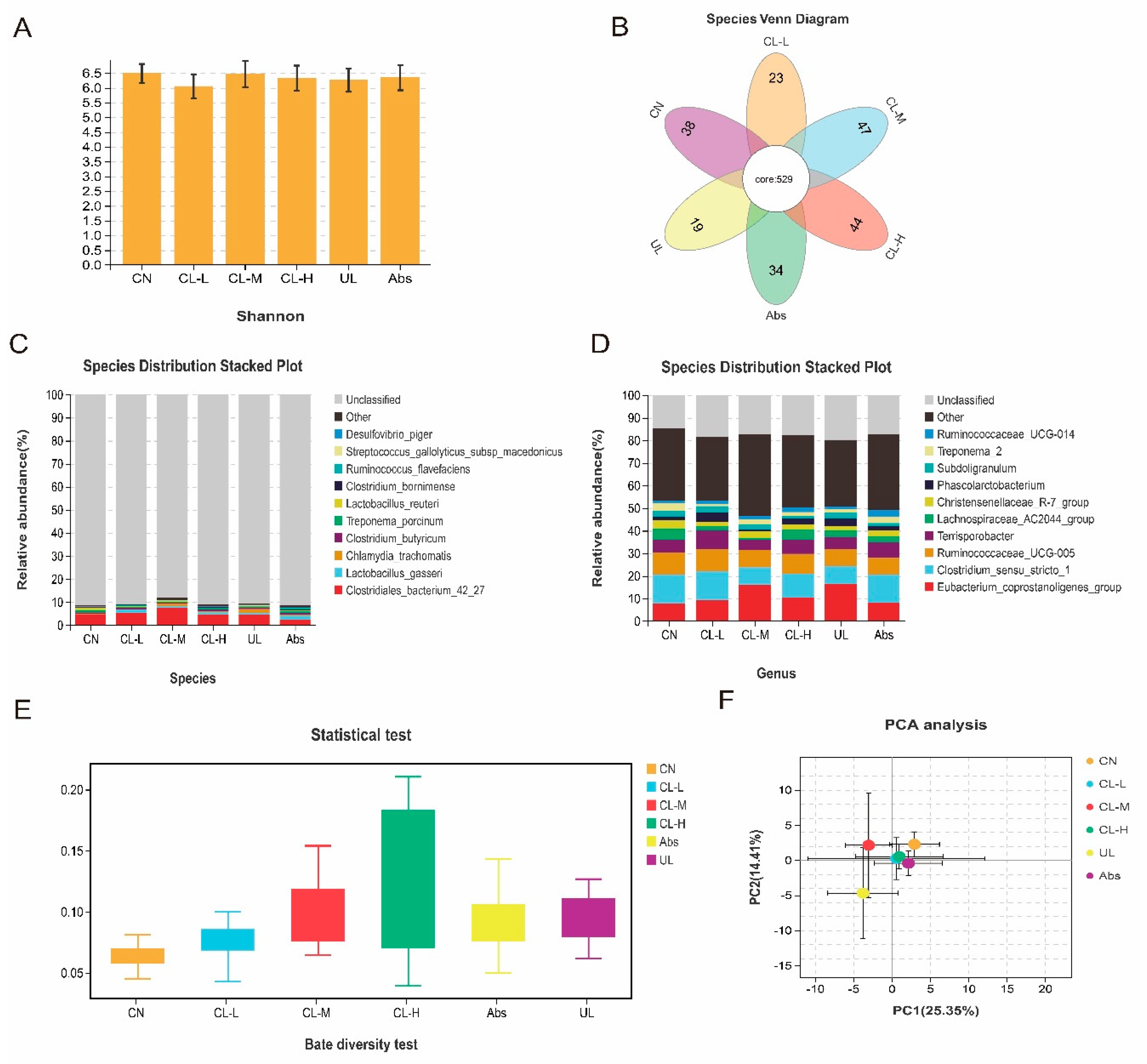
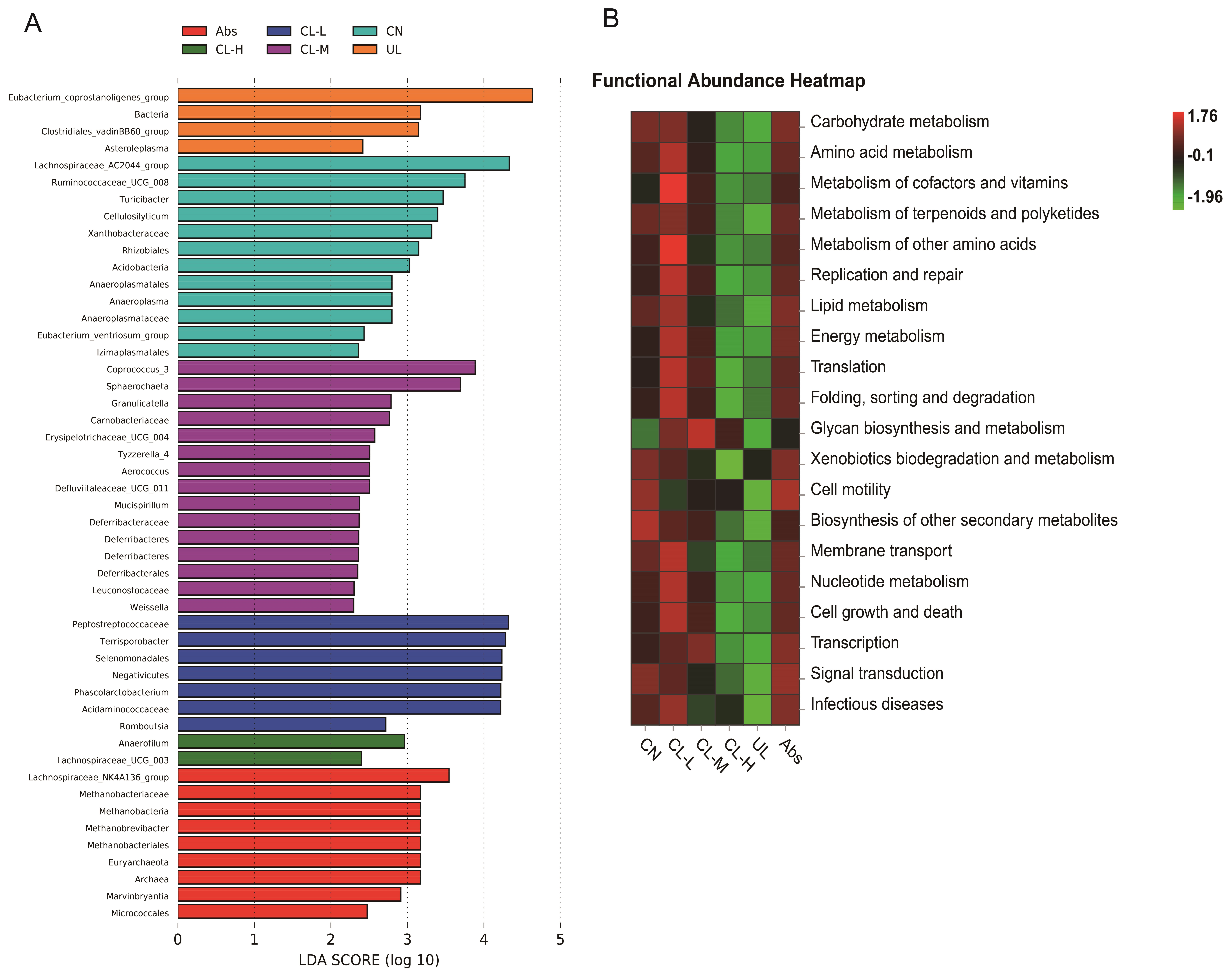
2.8. 16S rRNA Sequencing of the Cecal Microflora
3. Discussion
4. Materials and Methods
4.1. Animals, Treatment, and Designation
4.2. Experimental Design
4.3. Sample Collection
4.4. Growth Performance Evaluation
4.5. Determination of Serum Biochemical Index and Immunoglobulin Levels
4.6. Determination of Antioxidant Indexes
4.7. Digestive Enzymes
4.8. Morphological Analysis of the Small Intestine
4.9. Determination of Short-Chain Fatty Acid Levels
4.10. 16S rRNA Sequencing of the Cecal Microflora
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Heo, J.M.; Opapeju, F.O.; Pluske, J.R.; Kim, J.C.; Hampson, D.J.; Nyachoti, C.M. Gastrointestinal health and function in weaned pigs: A review of feeding strategies to control post-weaning diarrhoea without using in-feed antimicrobial compounds. J. Anim. Physiol. Anim. Nutr. 2013, 97, 207–237. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.H.; Xiao, K.; Luan, Z.S.; Song, J. Early weaning increases intestinal permeability, alters expression of cytokine and tight junction proteins, and activates mitogen-activated protein kinases in pigs. J. Anim. Sci. 2013, 91, 1094–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blavi, L.; Solà-Oriol, D.; Llonch, P.; López-Vergé, S.; Martín-Orúe, S.M.; Pérez, J.F. Management and Feeding Strategies in Early Life to Increase Piglet Performance and Welfare around Weaning: A Review. Animals 2021, 11, 302. [Google Scholar] [CrossRef]
- Brown, D.C.; Maxwell, C.V.; Erf, G.F.; Davis, M.E.; Singh, S.; Johnson, Z.B. The influence of different management systems and age on intestinal morphology, immune cell numbers and mucin production from goblet cells in post-weaning pigs. Vet. Immunol. Immunopathol. 2006, 111, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Guo, Y.; Wen, Z.; Jiang, X.; Ma, X.; Han, X. Weaning Stress Perturbs Gut Microbiome and Its Metabolic Profile in Piglets. Sci. Rep. 2018, 8, 18068. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.M.; Jiang, Z.Y.; Zheng, C.T.; Wang, L.; Yang, X.F. Effect of Lactobacillus plantarum on diarrhea and intestinal barrier function of young piglets challenged with enterotoxigenic Escherichia coli K88. J. Anim. Sci. 2014, 92, 1496–1503. [Google Scholar] [CrossRef] [Green Version]
- Meale, S.J.; Li, S.C.; Azevedo, P.; Derakhshani, H.; DeVries, T.J.; Plaizier, J.C.; Steele, M.A.; Khafipour, E. Weaning age influences the severity of gastrointestinal microbiome shifts in dairy calves. Sci. Rep. 2017, 7, 198. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Xiong, X.; Wang, X.; Tan, B.; Li, T.; Yin, Y. Effects of Weaning on Intestinal Upper Villus Epithelial Cells of Piglets. PLoS ONE 2016, 11, e0150216. [Google Scholar] [CrossRef] [Green Version]
- Cao, G.; Tao, F.; Hu, Y.; Li, Z.; Zhang, Y.; Deng, B.; Zhan, X. Positive effects of a Clostridium butyricum-based compound probiotic on growth performance, immune responses, intestinal morphology, hypothalamic neurotransmitters, and colonic microbiota in weaned piglets. Food Funct. 2019, 10, 2926–2934. [Google Scholar] [CrossRef]
- Bach Knudsen, K.E. Development of antibiotic resistance and options to replace antimicrobials in animal diets. Proc. Nutr. Soc. 2001, 60, 291–299. [Google Scholar]
- Cheng, G.; Hao, H.; Xie, S.; Wang, X.; Dai, M.; Huang, L.; Yuan, Z. Antibiotic alternatives: The substitution of antibiotics in animal husbandry? Front. Microbiol. 2014, 5, 217. [Google Scholar] [CrossRef] [PubMed]
- Thacker, P.A. Alternatives to antibiotics as growth promoters for use in swine production: A review. J. Anim. Sci. Biotechnol. 2013, 4, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verstegen, M.W.; Williams, B.A. Alternatives to the use of antibiotics as growth promoters for monogastric animals. Anim. Biotechnol. 2002, 13, 113–127. [Google Scholar] [CrossRef]
- Nyachoti, C.M.; Kiarie, E.; Bhandari, S.K.; Zhang, G.; Krause, D.O. Weaned pig responses to Escherichia coli K88 oral challenge when receiving a lysozyme supplement. J. Anim. Sci. 2012, 90, 252–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Guo, Y.; Wang, Z.; Yuan, J. Exogenous lysozyme influences Clostridium perfringens colonization and intestinal barrier function in broiler chickens. Avian Pathol. 2010, 39, 17–24. [Google Scholar] [CrossRef]
- Humphrey, B.D.; Huang, N.; Klasing, K.C. Rice expressing lactoferrin and lysozyme has antibiotic-like properties when fed to chicks. J. Nutr. 2002, 132, 1214–1218. [Google Scholar] [CrossRef] [Green Version]
- Masschalck, B.; Michiels, C.W. Antimicrobial properties of lysozyme in relation to foodborne vegetative bacteria. Crit. Rev. Microbiol. 2003, 29, 191–214. [Google Scholar] [CrossRef]
- Brugaletta, G.; De Cesare, A.; Laghi, L.; Manfreda, G.; Zampiga, M.; Oliveri, C.; Pérez-Calvo, E.; Litta, G.; Lolli, S.; Sirri, F. A multi-omics approach to elucidate the mechanisms of action of a dietary muramidase administered to broiler chickens. Sci. Rep. 2022, 12, 5559. [Google Scholar] [CrossRef]
- Ellison, R.T.; Giehl, T.J., III. Killing of gram-negative bacteria by lactoferrin and lysozyme. J. Clin. Investig. 1991, 88, 1080–1091. [Google Scholar] [CrossRef] [Green Version]
- Kawano, M.; Namba, Y.; Hanaoka, M. Regulatory factors of lymphocyte-lymphocyte interaction. I. Con A-induced mitogenic factor acts on the late G1 stage of T-cell proliferation. Microbiol. Immunol. 1981, 25, 505–515. [Google Scholar] [CrossRef]
- Ferraboschi, P.; Ciceri, S.; Grisenti, P. Applications of Lysozyme, an Innate Immune Defense Factor, as an Alternative Antibiotic. Antibiotics 2021, 10, 1534. [Google Scholar] [CrossRef] [PubMed]
- Oliver, W.T.; Wells, J.E. Lysozyme as an alternative to antibiotics improves growth performance and small intestinal morphology in nursery pigs. J. Anim. Sci. 2013, 91, 3129–3136. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Kong, J.; Zhang, G.; Zhang, X.; Seviour, R.; Kong, Y. Effects of dietary supplementation with lysozyme on the structure and function of the cecal microbiota in broiler chickens. PLoS ONE 2019, 14, e0216748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilcacundo, R.; Méndez, P.; Reyes, W.; Romero, H.; Pinto, A.; Carrillo, W. Antibacterial Activity of Hen Egg White Lysozyme Denatured by Thermal and Chemical Treatments. Sci. Pharm. 2018, 86, 48. [Google Scholar] [CrossRef] [Green Version]
- Diels, A.M.; De Taeye, J.; Michiels, C.W. Sensitisation of Escherichia coli to antibacterial peptides and enzymes by high-pressure homogenisation. Int. J. Food Microbiol. 2005, 105, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Kihm, D.J.; Leyer, G.J.; An, G.H.; Johnson, E.A. Sensitization of heat-treated Listeria monocytogenes to added lysozyme in milk. Appl. Environ. Microbiol. 1994, 60, 3854–3861. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Shi, J.; Shi, X.; Dong, Y.; Wu, X.; Li, Z.; Fang, Z.; Lin, Y.; Che, L.; Li, J.; et al. Effects of dietary supplementation with lysozyme during late gestation and lactation stage on the performance of sows and their offspring. J. Anim. Sci. 2018, 96, 4768–4779. [Google Scholar]
- May, K.D.; Wells, J.E.; Maxwell, C.V.; Oliver, W.T. Granulated lysozyme as an alternative to antibiotics improves growth performance and small intestinal morphology of 10-day-old pigs. J. Anim. Sci. 2012, 90, 1118–1125. [Google Scholar] [CrossRef]
- Yang, H.S.; Fu, D.Z.; Kong, X.F.; Wang, W.C.; Yang, X.J.; Nyachoti, C.M.; Yin, Y.L. Dietary supplementation with N-carbamylglutamate increases the expression of intestinal amino acid transporters in weaned Huanjiang mini-pig piglets. J. Anim. Sci. 2013, 91, 2740–2748. [Google Scholar] [CrossRef]
- Sancak, Y.; Peterson, T.R.; Shaul, Y.D.; Lindquist, R.A.; Thoreen, C.C.; Bar-Peled, L.; Sabatini, D.M. The Rag GTPases bind raptor and mediate amino acid signaling to mTORC1. Science 2008, 320, 1496–1501. [Google Scholar] [CrossRef] [Green Version]
- Che, L.Q.; Peng, X.; Hu, L.; Wu, C.; Xu, Q.; Fang, Z.F.; Lin, Y.; Xu, S.Y.; Li, J.; Feng, B.; et al. The addition of protein-bound amino acids in low-protein diets improves the metabolic and immunological characteristics in fifteen- to thirty-five-kg pigs. J. Anim. Sci. 2017, 95, 1277–1287. [Google Scholar] [PubMed]
- Figueroa, J.L.; Lewis, A.J.; Miller, P.S.; Fischer, R.L.; Diedrichsen, R.M. Growth, carcass traits, and plasma amino acid concentrations of gilts fed low-protein diets supplemented with amino acids including histidine, isoleucine, and valine. J. Anim. Sci. 2003, 81, 1529–1537. [Google Scholar] [CrossRef] [PubMed]
- Gómez, R.S.; Lewis, A.J.; Miller, P.S.; Chen, H.Y. Growth performance, diet apparent digestibility, and plasma metabolite concentrations of barrows fed corn-soybean meal diets or low-protein, amino acid-supplemented diets at different feeding level. J. Anim. Sci. 2002, 80, 644–653. [Google Scholar] [CrossRef]
- Tomas, F.M.; Campbell, R.G.; King, R.H.; Johnson, R.J.; Chandler, C.S.; Taverner, M.R. Growth hormone increases whole-body protein turnover in growing pigs. J. Anim. Sci. 1992, 70, 3138–3143. [Google Scholar] [CrossRef] [PubMed]
- Muri, J.; Kopf, M. Redox regulation of immunometabolism. Nat. Rev. Immunol. 2021, 21, 363–381. [Google Scholar] [CrossRef] [PubMed]
- Gong, G.; Qin, Y.; Huang, W.; Zhou, S.; Wu, X.; Yang, X.; Zhao, Y.; Li, D. Protective effects of diosgenin in the hyperlipidemic rat model and in human vascular endothelial cells against hydrogen peroxide-induced apoptosis. Chem. Biol. Interact. 2010, 184, 366–375. [Google Scholar] [CrossRef]
- Sauriasari, R.; Andrajati, R.; Azizahwati; Dharmeizar; Saputri, D.A.; Muris, R.U.; Manfaatun, A.; Amanda, O.F.; Setiawan, H.; Sakano, N.; et al. Marker of lipid peroxidation related to diabetic nephropathy in Indonesian type 2 diabetes mellitus patients. Diabetes Res. Clin. Pract. 2015, 108, 193–200. [Google Scholar] [CrossRef]
- Peixoto, E.B.; Pessoa, B.S.; Biswas, S.K.; Lopes de Faria, J.B. Antioxidant SOD mimetic prevents NADPH oxidase-induced oxidative stress and renal damage in the early stage of experimental diabetes and hypertension. Am. J. Nephrol. 2009, 29, 309–318. [Google Scholar] [CrossRef]
- Zhu, L.H.; Zhao, K.L.; Chen, X.L.; Xu, J.X. Impact of weaning and an antioxidant blend on intestinal barrier function and antioxidant status in pigs. J. Anim. Sci. 2012, 90, 2581–2589. [Google Scholar] [CrossRef]
- Xu, J.; Xu, C.; Chen, X.; Cai, X.; Yang, S.; Sheng, Y.; Wang, T. Regulation of an antioxidant blend on intestinal redox status and major microbiota in early weaned piglets. Nutrition 2014, 30, 584–589. [Google Scholar] [CrossRef]
- Samarghandian, S.; Azimi-Nezhad, M.; Farkhondeh, T.; Samini, F. Anti-oxidative effects of curcumin on immobilization-induced oxidative stress in rat brain, liver and kidney. Biomed. Pharmacother. 2017, 87, 223–229. [Google Scholar] [CrossRef]
- Switzar, L.; Giera, M.; Niessen, W.M. Protein digestion: An overview of the available techniques and recent developments. J. Proteome Res. 2013, 12, 1067–1077. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ni, X.; Wen, B.; Zhou, Y.; Liu, L.; Zeng, Y.; Zhao, W.; Khalique, A.; Wang, P.; Pan, K.; et al. Bacillus strains improve growth performance via enhancing digestive function and anti-disease ability in young and weaning rex rabbits. Appl. Microbiol. Biotechnol. 2020, 104, 4493–4504. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Cui, X.; Wang, Z.; Xiao, C.; Ji, Q.; Wei, Q.; Huang, Y.; Bao, G.; Liu, Y. Effects of Clostridium butyricum and a Bacteriophage Cocktail on Growth Performance, Serum Biochemistry, Digestive Enzyme Activities, Intestinal Morphology, Immune Responses, and the Intestinal Microbiota in Rabbits. Antibiotics 2021, 10, 1347. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.H.; Holtrop, G.; Lobley, G.E.; Calder, A.G.; Stewart, C.S.; Flint, H.J. Contribution of acetate to butyrate formation by human faecal bacteria. Br. J. Nutr. 2004, 91, 915–923. [Google Scholar] [CrossRef] [Green Version]
- Chambers, E.S.; Viardot, A.; Psichas, A.; Morrison, D.J.; Murphy, K.G.; Zac-Varghese, S.E.; MacDougall, K.; Preston, T.; Tedford, C.; Finlayson, G.S.; et al. Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adults. Gut 2015, 64, 1744–1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mountzouris, K.C.; Tsirtsikos, P.; Kalamara, E.; Nitsch, S.; Schatzmayr, G.; Fegeros, K. Evaluation of the efficacy of a probiotic containing Lactobacillus, Bifidobacterium, Enterococcus, and Pediococcus strains in promoting broiler performance and modulating cecal microflora composition and metabolic activities. Poult. Sci. 2007, 86, 309–317. [Google Scholar] [CrossRef]
- Shi, F.; Li, Y.; Han, R.; Fu, A.; Wang, R.; Nusbaum, O.; Qin, Q.; Chen, X.; Hou, L.; Zhu, Y. Valerian and valeric acid inhibit growth of breast cancer cells possibly by mediating epigenetic modifications. Sci. Rep. 2021, 11, 2519. [Google Scholar] [CrossRef]
- Jayaraj, R.L.; Beiram, R.; Azimullah, S.; Mf, N.M.; Ojha, S.K.; Adem, A.; Jalal, F.Y. Valeric Acid Protects Dopaminergic Neurons by Suppressing Oxidative Stress, Neuroinflammation and Modulating Autophagy Pathways. Int. J. Mol. Sci. 2020, 21, 7670. [Google Scholar] [CrossRef]
- Kelly, D.; Conway, S.; Aminov, R. Commensal gut bacteria: Mechanisms of immune modulation. Trends Immunol. 2005, 26, 326–333. [Google Scholar] [CrossRef]
- Sommer, F.; Bäckhed, F. The gut microbiota—Masters of host development and physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.; Su, W.; Rahat-Rozenbloom, S.; Wolever, T.M.; Comelli, E.M. Adiposity, gut microbiota and faecal short chain fatty acids are linked in adult humans. Nutr. Diabetes 2014, 4, e121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, C.; Zhang, Z.; Morales, M.; Wang, Y.; Khafipour, E.; Friel, J. Feeding practice influences gut microbiome composition in very low birth weight preterm infants and the association with oxidative stress: A prospective cohort study. Free Radic. Biol. Med. 2019, 142, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef]
- Liu, Y.; Lin, Q.; Huang, X.; Jiang, G.; Li, C.; Zhang, X.; Liu, S.; He, L.; Liu, Y.; Dai, Q.; et al. Effects of Dietary Ferulic Acid on the Intestinal Microbiota and the Associated Changes on the Growth Performance, Serum Cytokine Profile, and Intestinal Morphology in Ducks. Front. Microbiol. 2021, 12, 698213. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CN | CL-L | CL-M | CL-H | UL | Abs | SEM | p Value | |
|---|---|---|---|---|---|---|---|---|
| (d 0) Initial weight (kg) | 8.66 ± 0.95 | 8.71 ± 0.31 | 9.18 ± 0.24 | 8.91 ± 0.25 | 8.62 ± 0.34 | 9.08 ± 0.49 | 85.07 | 0.299 |
| (d 14) Middle weight (kg) | 12.74 ± 0.80 c | 13.11 ± 0.25 c | 14.26 ± 0.51 abc | 14.60 ± 0.31 ab | 13.78 ± 1.12 bc | 16.06 ± 0.19 a | 222.54 | 0.007 |
| (d 28) Final weight (kg) | 16.88 ± 1.30 b | 1.81 ± 0.19 b | 19.07 ± 0.77 b | 20.84 ± 0.67 a | 18.47 ± 1.60 b | 21.98 ± 0.47 a | 373.22 | 0.000 |
| ADG (g) | ||||||||
| Days 0 to 14 | 339.13 ± 49.85 bc | 313.87 ± 14.54 c | 362.67 ± 35.29 bc | 406.24 ± 4.50 ab | 368.33 ± 61.36 bc | 451.23 ± 21.60 a | 13.08 | 0.009 |
| Days 14 to 28 | 295.71 ± 40.92 c | 358.77 ± 22.72 abc | 343.62 ± 18.86 bc | 445.74 ± 68.57 a | 334.91 ± 81.18 bc | 422.80 ± 41.55 ab | 16.09 | 0.029 |
| Days 0 to 28 | 317.42 ± 24.74 b | 336.32 ± 8.75 b | 353.14 ± 25.86 b | 425.99 ± 32.48 a | 351.62 ± 55.86 b | 437.01 ± 26.31 a | 12.64 | 0.003 |
| ADFI (g) | 672.57 ± 4.92 a | 613.50 ± 11.54 b | 605.60 ± 10.66 b | 649.26 ± 22.38 a | 544.97 ± 12.35 c | 673.90 ± 21.50 a | 11.31 | 0.000 |
| F/G | 2.13 ± 0.15 a | 1.82 ± 0.05 b | 1.72 ± 0.11 b | 1.53 ± 0.06 c | 1.58 ± 0.28 b | 1.54 ± 0.05 c | 0.06 | 0.002 |
| CN | CL-L | CL-M | CL-H | UL | Abs | SEM | p Value | |
|---|---|---|---|---|---|---|---|---|
| CAT (U/mL) | 169.01 ± 56.16 b | 242.58 ± 51.59 a | 263.70 ± 50.18 a | 281.09 ± 44.31 a | 280.33 ± 54.93 a | 291.10 ± 50.60 a | 10.77 | 0.003 |
| GSH-Px (U/L) | 80.03 ± 11.93 d | 105.68 ± 23.53 c | 103.72 ± 13.89 c | 135.71 ± 18.13 ab | 145.50 ± 14.28 a | 118.44 ± 20.24 bc | 4.62 | 0.000 |
| MDA (nmol/mL) | 5.97 ± 1.20 a | 5.61 ± 1.12 ab | 4.20 ± 1.04 bc | 4.39 ± 1.38 bc | 3.25 ± 1.37 c | 5.39 ± 1.04 ab | 0.25 | 0.005 |
| SOD (U/mL) | 195.03 ± 45.01 d | 258.78 ± 39.32 ab | 300.52 ± 51.47 abc | 329.88 ± 52.14 ab | 343.96 ± 53.61 a | 270.14 ± 63.65 bc | 11.79 | 0.000 |
| T-AOC (μmol/mL) | 1.54 ± 0.31 c | 2.05 ± 0.23 b | 2.19 ± 0.35 ab | 2.49 ± 0.54 a | 2.58 ± 0.25 a | 2.07 ± 0.35 b | 0.08 | 0.000 |
| CN | CL-L | CL-M | CL-H | UL | Abs | SEM | p Value | |
|---|---|---|---|---|---|---|---|---|
| acetic acid | 22.54 ± 8.77 a | 14.64 ± 2.30 c | 13.42 ± 2.90 c | 15.74 ± 2.41 ab | 14.47 ± 2.75 c | 17.53 ± 4.72 ab | 1.04 | 0.113 |
| propionic acid | 8.71 ± 2.85 a | 6.34 ± 0.66 ab | 5.40 ± 1.34 b | 6.39 ± 0.76 ab | 5.87 ± 0.58 b | 7.80 ± 2.42 ab | 0.39 | 0.100 |
| isobutyric acid | 0.66 ± 0.15 a | 0.56 ± 0.09 a | 0.44 ± 0.12 a | 0.61 ± 0.14 a | 0.60 ± 0.16 a | 0.63 ± 0.29 a | 0.03 | 0.521 |
| butyric acid | 4.16 ± 1.05 a | 2.60 ± 0.61 bc | 1.92 ± 0.74 c | 2.69 ± 0.43 bc | 2.13 ± 1.07 bc | 3.37 ± 0.62 ab | 0.21 | 0.008 |
| isovaleric acid | 1.11 ± 0.25 a | 0.90 ± 0.16 a | 0.72 ± 0.18 a | 1.05 ± 0.20 a | 1.05 ± 0.35 a | 1.15 ± 0.41 a | 0.06 | 0.284 |
| valeric acid | 0.80 ± 0.16 ab | 0.68 ± 0.08 ab | 0.59 ± 0.10 b | 0.85 ± 0.09 a | 0.71 ± 0.14 ab | 0.87 ± 0.21 a | 0.03 | 0.075 |
| Diet Composition | Percentage (%) | Nutrient Levels | Content |
|---|---|---|---|
| Corn | 62.12 | Total (MJ/kg) | 14.55 |
| Bean meal | 24.88 | Crude protein | 17.4 |
| Wheat bran | 3 | Total phosphorus | 0.51 |
| Fatty powder | 2.0 | Lysine | 1.158 |
| Limestone | 1.2 | Methionine+cysteine | 0.612 |
| Calcium hydrogen phosphate | 0.8 | Threonine | 0.679 |
| Soybean oil | 0.07 | ||
| Fish meal | 1 | ||
| NaCl | 0.3 | ||
| Choline chloride | 0.1 | ||
| Potassium magnesium sulfate | 0.2 | ||
| Premix * | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Huang, P.; Cui, X.; Li, X.; Sun, J.; Ji, Q.; Wei, Q.; Huang, Y.; Li, Z.; Bao, G.; et al. Effects of Dietary Coated Lysozyme on the Growth Performance, Antioxidant Activity, Immunity and Gut Health of Weaned Piglets. Antibiotics 2022, 11, 1470. https://doi.org/10.3390/antibiotics11111470
Xu X, Huang P, Cui X, Li X, Sun J, Ji Q, Wei Q, Huang Y, Li Z, Bao G, et al. Effects of Dietary Coated Lysozyme on the Growth Performance, Antioxidant Activity, Immunity and Gut Health of Weaned Piglets. Antibiotics. 2022; 11(11):1470. https://doi.org/10.3390/antibiotics11111470
Chicago/Turabian StyleXu, Xiangfei, Pan Huang, Xuemei Cui, Xuefeng Li, Jiaying Sun, Quanan Ji, Qiang Wei, Yee Huang, Zhefeng Li, Guolian Bao, and et al. 2022. "Effects of Dietary Coated Lysozyme on the Growth Performance, Antioxidant Activity, Immunity and Gut Health of Weaned Piglets" Antibiotics 11, no. 11: 1470. https://doi.org/10.3390/antibiotics11111470