3.2. Epidemiological and Clinical Characteristics
The number of sequences included in the database increased year after year. Indeed, nearly 50% of the overall sequences and more than 50% of the clinical-related sequences were included in the last three years. This increase may be related to the higher feasibility of performing genetic analysis owing to a decrease in cost and availability of new technologies, but an increasing concern with regard to the diffusion of these infections may also be hypothesized.
Europe is the area where about 50% of the sequences originated. When these data are compared to areas with a similar level of health services (Americas and Australia), the frequency observed was much lower, suggesting that the high frequency of European sequences is related to a higher prevalence of GBS infections on the continent. The known high frequency of LA-GBS, particularly in dairy cows [
8,
18], supports the importance of promoting the registration of LA-GBS sequences in the database to favor the investigation of the measures to reduce the risk of transmission between humans and animals.
Blood represents 25% of overall biological sources of GBS included in the database, while 25% was related to the urinary and intestinal tract (including urine and feces). Milk represents 10% of the whole sequences and only 2.9% of the clinical ones. These data suggest that the role of direct infections through milk in humans is of minor importance compared to the role in dairy cows, where milk is considered the major source of infection [
19,
20]. Moreover, it can be hypothesized that the pathogenetic characteristics of humans or LA-GBS are different, even if an acquisition of the adaptation gene to an alternative species is common [
12,
21].
3.3. Antimicrobial Resistance Pattern
The database considers 19 different AMR-genes, but only eight were recovered from the sequences with a proportion >100. Tetracycline resistance in GBS is ubiquitously high (usually >80%), and most GBS strains are characterized by the presence of resistance elements,
tetO and
tetM [
3]. Indeed,
tetM is the gene with the higher frequency both in the whole and in the clinical-related database. This gene is related to GBS tetracycline resistance that includes the genes for efflux proteins like TetK and TetL or to ribosomal protection proteins like TetM and TetO. Efflux proteins belong to the major facilitator superfamily (MFS) and all
tet efflux genes encode membrane-associated proteins that export tetracycline from the cell, reducing the intracellular concentration of the antibiotic. In GBS, these TetK and TetL efflux proteins are encoded by
tetK and
tetL genes, respectively, and are usually located on large plasmids or plasmids that can integrate into the bacterial chromosome [
22]. These proteins are responsible for detaching tetracycline molecules from the ribosome, thus, the aminoacyl-tRNA molecules can bind again to the ribosomal A-site allowing protein synthesis to continue [
23,
24]. Since its discovery, tetracycline has been extensively overused, and thus resistance to this antibiotic is now widely observed, as confirmed by the results of this study.
The gene with the second higher frequency is
ermB, which is related to macrolide resistance. Indeed, the methyl-transferases encoded by the
erm gene family, composed of more than 40
erm variants, represent the most common macrolide resistance mechanism in pathogenic bacteria via ribosomal methylation.
ermB is considered the most widespread
erm family gene in Streptococci and GBS [
3], as confirmed by this study.
ant(6)la has the highest frequency in clinical-related sequences after the previous two genes, and its frequency was significantly higher than expected. This gene is related to resistance to aminoglycosides [
25]. The presence of this gene in isolates from human diseases has peculiar importance from a One Health approach because, recently, it was also recovered from streptococci of animal origin [
26], supporting the risk of a bi-directional transfer of resistance gene.
The very low frequency of genes related to penicillin-resistance GBS, such as
pbp1a, pbp2a, pbp2b, and
pbp2x, confirmed that overall penicillin resistance is still relatively rare [
3]. Analogously, a very low frequency of genes related to vancomycin resistance was observed. The resistance is related to the
vanG gene responsible for altering the vancomycin target site to
d-Ala-
d-Ser. It was suggested that the lack of vancomycin-sensitivity testing might bias the low frequency of vancomycin resistance isolated due to universal susceptibility to penicillin in GBS strains [
3]. However, the analysis of the sequences with very low presence of
van family genes confirms the low frequency of the vancomycin resistance observed in the field. The same results were observed for gentamycin resistance, associated with
aacA-aphD gene [
27], which was identified in less than four sequences. These data support the evidence of a low frequency for clinical isolates (<0.5%) [
1,
27].
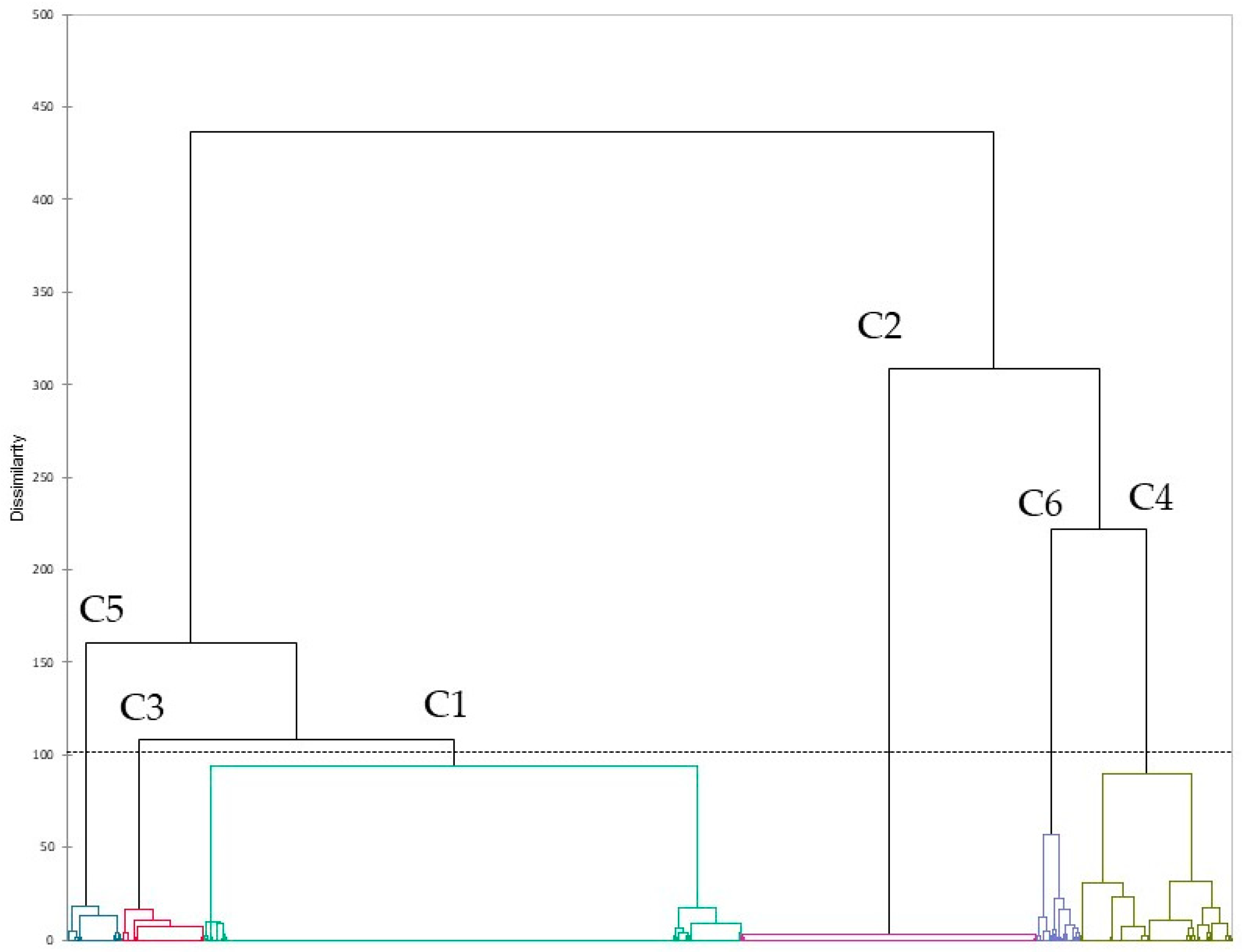
The pathogenetic and the AMR characteristics of pathogens are the results of the combination of their virulence and AMR genes. A cluster analysis was conducted to investigate the pattern of AMR genes in GBS, allowing to identify six different clusters. One cluster (C2) included isolates without AMR genes, while another (C6) included sequences positives for the 10 most frequent AMR genes, even with different proportions. All the other four clusters included sequences with a high proportion of tetracycline gene resistance associated with macrolide (C3), aminoglycosides (C4), and
msrD gene. This latter one is involved in resistance to azithromycin by acting as a ribosomal protection protein and displacing the macrolides from the ribosome. It is common in other Streptococci, supporting the evidence of AMR gene transmission among different streptococcal species [
28].
Sequences included in C5 and C6 have a higher frequency in 2021–2022, and C6 sequences were not found until 2016, whereas C1 and C2 (with lower proportion of AMR genes) had a lower-than-expected frequency in recent years, even not significantly for C2 in clinical isolates. In this latter case, a significant increase in C4 was observed in recent years, suggesting a potential role of this specific resistance mechanism in the development of clinical cases. Moreover, this cluster also has a significantly higher frequency in Asiatic and European isolates, even if the association with clinical cases was confirmed only for asiatic isolates. In this latter continent, a significant increase in C6 was also observed. C2 showed a significantly lower-than-expected frequency in Europe, supporting the previous hypothesis of a role of the acquisition of new resistance genes in the development of GBS disease. It should be noted that Asian sequences provided a contradicting signal. Specifically, both C2 (no AMR) and C6 (high AMR) had a frequency greater than expected. This result suggests that the spread and development of GBS infections in an area with an apparent low prevalence may be due to strains without any significant AMR, but their treatment could lead to an increase in AMR very rapidly and the spread of the resistant strains.
C6 was typically observed in isolates from the uro-vaginal and rectal tract, including urine and feces, as expected. On the other hand, milk was the major source of C2, which supports the very low frequency of clinical isolates from milk, and, therefore, a probable low frequency of antimicrobial treatment.


 ,
, 

{kind=link}