
Rhodiola rosea Reduces Intercellular Signaling in Campylobacter jejuni


1
Department of Food Science and Technology, Biotechnical Faculty, University of Ljubljana, Jamnikarjeva ulica 101, 1000 Ljubljana, Slovenia
2
Department of Pharmacognosy, Institute of Pharmaceutical Sciences, University of Graz, Beethovenstraße 8, 8010 Graz, Austria
*
Author to whom correspondence should be addressed.
Antibiotics 2022, 11(9), 1220; https://doi.org/10.3390/antibiotics11091220
Submission received: 27 July 2022
/
Revised: 2 September 2022
/
Accepted: 5 September 2022
/
Published: 8 September 2022
(This article belongs to the Special Issue Antimicrobial Activity of Different Plant Extracts, Plant-Derived Compounds and Synthetic Derivatives of Natural Compounds on Pathogenic Microorganisms)
Abstract
:Campylobacter jejuni is a major foodborne pathogen and the leading cause of bacterial gastroenteritis, i.e., campylobacteriosis. Besides searching for novel antimicrobials, identification of new targets for their action is becoming increasingly important. Rhodiola rosea has long been used in traditional medicine. Ethanolic extracts from the roots and rhizomes of the plant contain a wide range of bioactive compounds with various pharmacological activities. In this study, cultivated plant materials have been used, i.e., “Mattmark” and “Rosavine”. Through optimized protocols, we obtained fractions of the initial ethanolic extracts rich in most important bioactive compounds from R. rosea, including salidroside, rosavins, proanthocyanidins (PACs), and flavonoids. The antimicrobial activity in relation to the chemical composition of the extracts and their fractions was studied with an emphasis on C. jejuni AI-2-mediated intercellular signaling. At concentration 15.625 mg/L, bioluminescence reduction rates varied from 27% to 72%, and the membrane remained intact. Fractions rich in PACs had the strongest antimicrobial effect against C. jejuni, with the lowest minimal inhibitory concentrations (MICs) (M F3 40%: 62.5 mg/L; R F3 40%: 250 mg/L) and the highest intercellular signaling reduction rates (M F3 40%: 72%; R F3 40%: 65%). On the other hand, fractions without PACs were less effective (MICs: M F5 PVP: 250 mg/L; R F5 PVP: 1000 mg/L and bioluminescence reduction rates: M F5 PVP: 27%; R F5 PVP: 43%). Additionally, fractions rich in flavonoids had strong antimicrobial activity (MICs: M F4 70%: 125 mg/L; R F4 70%: 250 mg/L and bioluminescence reduction rates: M F4 70%: 68%; R F4 70%: 50%). We conclude that PACs and flavonoids are crucial compound groups responsible for the antimicrobial activity of R. rosea roots and rhizomes in C. jejuni.
1. Introduction
Overuse of antibiotics in veterinary and human medicine is responsible for the development of antibiotic-resistant strains. Due to increased antimicrobial resistance, the search for novel antimicrobial agents and their potential targets is of utmost importance. Antibiotic resistance has also been recorded for Campylobacter jejuni, a major foodborne pathogen and the leading cause of bacterial gastroenteritis, also known as campylobacteriosis. This infection occurs very commonly in European countries. With over 120,000 confirmed human cases of illness in 2020, it has been the most commonly reported zoonosis, representing over 60% of all reported cases in 2020 [1].
There are different potential targets that can be affected by antimicrobials. Among them are bacterial intercellular communication systems, collectively termed quorum sensing (QS) [2]. QS is a bacterial cell-to-cell communication and it refers to the ability of bacteria to sense information from other cells in the population [3]. Bacteria communicate through small molecules also known as autoinducers (AIs) [4]. The autoinducer-2 (AI-2) signaling molecule regulates interspecies communication as it is present in both Gram-positive and Gram-negative bacteria [5,6]. Elvers and Park [7] described the role of the luxS gene in the production of the AI-2 signal and AI-2-mediated signaling in C. jejuni. The LuxS enzyme has a central role as a metabolic enzyme in the methyl cycle, responsible for the generation of S-adenosyl-L-methionine (SAM). The enzyme has its role in the two-step reaction of homocysteine formation, where AI-2 is formed as a by-product [8]. As LuxS-deficient C. jejuni shows critical differences in colonization and virulence [9], AI-2-mediated signaling can be considered as a potential target for the control of that pathogen.
Natural compounds with anti-QS potential can be found in medicinal plants and their extracts [10]. Various phytochemicals inhibit C. jejuni intercellular signaling. Examples of this include citrus ethanolic extracts [11], epigallocatechin gallate (EGCG) [12], resveratrol inclusion complexes [13], coriander essential oil, linalool [14], Euodia ruticarpa ethanolic extract [15], carvacrol [16], curcumin, allyl sulfide, garlic oil, and ginger oil [17]. Šimunović et al. [9] investigated the correlation of QS inhibition with changes in C. jejuni motility, adhesion to polystyrene surfaces, and adhesion to and invasion of INT407 cells. A positive correlation was reported between C. jejuni QS reduction and reduced motility, adhesion to polystyrene surfaces, and invasion. In their screening, among 20 natural extracts, essential oils, and pure compounds, Rhodiola rosea ethanolic extract showed the best overall antimicrobial activity against C. jejuni.
Rhodiola rosea L. (Sedum roseum (L.) Scop.) is an herbaceous plant belonging to the family Crassulaceae. Vernacular names by which we can recognize the plant include roseroot, golden root, and arctic root [18]. The plant grows up to 70 cm in height and has fleshy, succulent leaves. The flowers form a compact whorled inflorescence on top of the halms. The root system forms thick rhizomes [19].
R. rosea has long been used in traditional medicine. A key factor determining the quality of its formulations is the quantification of salidroside, rosavin, and rosarin [20]. In Northern Europe, Russia, and North America, R. rosea extracts are standardized to contain at least 3% rosavins and from 0.8% to 1% salidroside [21,22]. The majority of these extracts are derived from wild plants harvested in Russia and Mongolia, which threatens the long-term conservation of natural populations. In addition, fraudulent material containing non-R. rosea plant material is suspected to be on the market. In order to preserve the natural sources of R. rosea plant material (the collection of which is prohibited in many countries), and to ensure the quality and authenticity of the plant material, domestication and cultivation of the plant seems to be the most appropriate solution [23].
Extracts of the plant’s underground organs contain various compounds, including monoterpene alcohols and their glycosides, cyanogenic glycosides, aryl glycosides, phenylethanoids (salidroside, p-tyrosol), phenylpropanoids and their glycosides (rosavins), flavonoids, flavonolignans, proanthocyanidins (PACs), and gallic acid derivatives [24]. According to Ma et al. [25], salidroside, p-tyrosol, rosarin, rosavin, rosin, and rosiridin are responsible for the biological activity of R. rosea. Besides being used in pharmaceutical preparations, the plant is popular as a food additive and dietary supplement. The plant’s underground organs can be used as a crude drug (dried and powdered) or as an extract [26].
Due to its wide range of biologically active compounds, the plant has various pharmacological activities. Among them are antioxidative, anti-inflammatory, anti-fatigue, anti-depressive, anxiolytic, anti-tumor, antimicrobial, neuroprotective, cardioprotective, and normalizing endocrine and immune activities [24,27,28,29].
In the present study, ethanolic extracts were prepared from R. rosea cultivated plant material, i.e., “Mattmark” and “Rosavine”, of which “Mattmark” is the first synthetic R. rosea cultivar [23]. For the first time, biologically active compounds or compound groups from R. rosea were separated into different fractions, and antimicrobial activity, with an emphasis on AI-2-mediated intercellular signaling in relation to the chemical composition of the extracts and their fractions, was studied. Due to great antimicrobial activity having been previously reported for the ethanolic extract from R. rosea in C. jejuni, the aim of the study was to evaluate whether certain compounds or compound groups are crucial for this activity.
2. Material and Methods
2.1. Bacterial Strains and Growth Conditions
C. jejuni NCTC 11168, C. jejuni 11168∆luxS, and Vibrio harveyi MM30 are part of Chair of Biotechnology, Microbiology and Food Safety (Biotechnical Faculty, University of Ljubljana, Ljubljana, Slovenia) strain collection and were stored at −80 °C. C. jejuni strains were stored in a 20% glycerol (Kemika, Zagreb, Croatia) and an 80% Muller-Hinton broth (MHB) (Oxoid, Hampshire, UK) solution. V. harveyi MM30 was stored in a 20% glycerol and an 80% AI bioassay (AB) liquid medium solution (composed as described in [9]). A reference strain, C. jejuni NCTC 11168, was revitalized by cultivating on Karmali selective media (Oxoid, Hampshire, UK), followed by cultivating on Mueller–Hinton agar (MHA) (Oxoid, Hampshire, UK) at 42 °C in microaerophilic conditions (85% N2, 10% CO2 and 5% O2). A mutant strain, C. jejuni 11168∆luxS, which does not produce the AI-2 signal [2], was cultivated on MHA with the addition of 30 mg/L kanamycin (Merck, Darmstadt, Germany). A reporter strain, V. harveyi MM30, with eliminated AI-2 production [30], was used for AI-2 bioassays and was directly inoculated into 5 mL of AB liquid medium and incubated at 30 °C in aerobic conditions.
2.2. Plant Material
R. rosea roots and rhizomes from cultivated plant material were provided by Dr. Christoph Carlen and Mr. Claude-Alain Carron (Agroscope, Conthey, Switzerland). Two types of plant material were used: “Mattmark”, harvested in April 2017 in Conthey at an altitude of 460 m, and “Rosavine”, harvested in June 2012 in Bruson at an altitude of 1050 m. Voucher specimens were kept at the Institute of Pharmaceutical Sciences, Department of Pharmacognosy, University of Graz.
2.3. Extracts Preparation
Ethanolic extracts were prepared from the roots and rhizomes of the plant. Sliced dried plant material was ground and about 50 g was used for the extraction. The lipophilic compounds were removed by pre-extraction with hexane (Carl Roth, Karlsruhe, Germany). Ethanolic extraction with 96% ethanol (Carl Roth, Karlsruhe, Germany) took place in an ultrasonic bath (Elma, Singen, Germany) for 10 min at room temperature, followed by centrifugation at 4000 RPM for 10 min. Ethanolic extraction was repeated three times, the supernatants were collected, and the solvent was evaporated using a rotary evaporator (Heidolph, Schwabach, Germany), followed by freeze drying using a VirTis Sentury freeze dryer (SP Scientific, Buena, CA, USA) resulting in 28.52% (“Mattmark”) and 32.47% (“Rosavine”) extraction yields. The ethanolic extracts, designated as M/R 96% EtOH, were stored in dark glass flasks in a fridge at 4 °C.
2.4. Fractionation of the Extracts: Fractions Preparation
Five fractions, enriched with different compounds or compound groups, were obtained from each crude ethanolic extract. Fractionation was performed on DIAION HP-20 adsorbent resin (Sigma-Aldrich, Steinheim, Germany) or using Polyclar AT (polyvinylpyrrolidone, or PVP) (Serva, Heidelberg, Germany). For the elution of compounds from the column, we used micropure water (MW; Barnstead Easypure RF) and different concentrations of methanol (Carl Roth, Karlsruhe, Germany).
The separation protocol described by Sun et al. [31] for the purification of salidroside and p-tyrosol was modified to obtain fractions of the ethanolic extracts as follows. DIAION HP-20 adsorbent resin was pre-treated by soaking it in 70% methanol/MW solution at 4 °C overnight, followed by washing with MW in a Buchner funnel until no alcohol remained. Then, 100 mg of crude ethanolic extract was dissolved in 10 mL of MW and centrifuged at 4000 RPM for 10 min at room temperature. The dissolved extract was mixed with 15 g of pre-treated DIAION HP-20 adsorbent resin. After full adsorption of substances to the adsorbent, it was loaded into a glass column and the bed volume (BV) was determined as 24 mL. Subsequently, the column was eluted with two BV of MW and two BV of a 20%, 40%, and 70% methanol/MW solution. The eluents were collected and their composition was determined directly by UHPLC-PDA-ESI-MS analysis. The optimized elution protocol was scaled up by a factor of 12 in order to obtain enough material for further microbiological analysis, resulting in fractions of M/R F1 0%, M/R F2 20%, M/R F3 40%, and M/R F4 70%.
The fifth fraction of each crude ethanolic extract was prepared using PVP, which mainly binds compounds with phenolic OH groups, e.g., tannins, PACs, and many flavonoids. Therefore, 50 mg of crude ethanolic extract was dissolved in 15 mL of 50% methanol/MW solution, followed by centrifugation at 4000 RPM for 10 min at room temperature. The supernatant was added to 1.25 g of PVP, mixed, and again centrifuged at 4000 RPM for 10 min at room temperature. The supernatant was filtered through a filter paper (Carl Roth, Karlsruhe, Germany) and the solvent was evaporated using a rotary evaporator, followed by freeze drying. In this case, the optimized protocol was scaled up by a factor of 20 to obtain enough material for further microbiological analysis, resulting in fractions M/R F5 PVP.
2.5. Chemical Characterization (UHPLC-PDA-ESI-MS Analysis)
Analysis of crude extracts and their fractions was carried out on two Dionex UltiMate 3000 UHPLC systems (Thermo Fisher Scientific, Waltham, CA, USA). The first system was coupled to a linear ion-trap mass spectrometer (MS), LTQ XL, equipped with an electrospray ionization (ESI) ion source (Thermo Fisher Scientific, Waltham, California, USA). Both systems were equipped with a photodiode array detector (PDA) (Thermo Fisher Scientific, Waltham, California, USA). Separation was undertaken on a Zorbax SB-C18 rapid resolution HD column (Agilent, Santa Clara, California, USA), 100 × 2.1 mm, 1.8 µm particle size. The mobile phase consisted of MW + 0.1% formic acid (A) (Honeywell Fluka, Seelze, Germany) and acetonitrile (VWR International, Rosny-sous-Bois-cedex, France) + 0.1% formic acid (B). A gradient elution was performed, starting with 2% B, increasing to 22% B at 13.33 min, 70% B at 22.22 min, then dropping back to 2% B at 22.67 min and keeping this composition until the end (28 min). The column temperature was set to 40 °C. Flow rate was set to 0.450 mL/min. Injection volume was 2 μL. Samples of crude ethanolic extracts were prepared using 20% ethanol/MW (2:8) in a concentration of 5 mg/mL and centrifuged at 13,000 RPM for 10 min at room temperature before analysis. Fractions were prepared using MW and different concentrations of ethanol/MW or methanol/MW. Therefore, we analyzed the fractions directly, after centrifugation at 13,000 RPM for 10 min at room temperature. PDA detection was performed in the 190 nm to 500 nm wavelength range. The mass spectra were recorded in negative and positive ion mode in the m/z range of 50 to 2000 amu. Mass spectral conditions were set as follows: source voltage 5.0 kV (ESI positive), 3.5 kV (ESI negative); capillary temperature 350 °C; source temperature 300 °C; sheath gas flow 40 arb, auxiliary gas flow 10 arb.
2.6. Determination of Minimal Inhibitory Concentrations (MICs)
The MICs of preparations against C. jejuni NCTC 11168 and C. jejuni 11168∆luxS were determined by the broth microdilution method previously described [32] with some modifications. Stock solutions were prepared in dimethyl sulfoxide (DMSO) (Merck, Germany) and later diluted in MHB to a final concentration of 1000 mg/L and 2.5% of DMSO. The bacterial inoculum contained approximately 5 × 105 colony-forming units (CFU)/mL. Cell viability was determined using resazurin fluorescent dye solution, prepared as previously described [9]. The fluorescence intensity was measured using the microplate reader Varioskan LUX (Thermo Fischer Scientific, Waltham, CA, USA) at an excitation wavelength of 560 nm and an emission wavelength of 590 nm. The MIC represents the lowest concentration where fluorescence, expressed in relative fluorescence units (RFU), reached the value of the negative control. The MICs were determined in triplicate.
2.7. AI-2 Bioassay
AI-2 bioassays were performed to evaluate the influence of the preparations on C. jejuni AI-2-mediated intercellular signaling. The cell-free spent media (SMs) were prepared and AI-2 bioassays were performed as described in [9]. Briefly, C. jejuni NCTC 11168 (C. j. 11168-Tr.) and C. jejuni 11168∆luxS (C. j. 11168∆luxS-Tr.) cultures with approximately 5 × 105 CFU/mL were treated separately with sub-inhibitory concentrations of preparations (i.e., 0.25× MIC or lower). Stock solutions were prepared in DMSO and were further diluted in MHB to a final 1% of DMSO. After 24 h incubation, SMs were prepared by filter-sterilization through 0.2 µm syringe filters (Sartorius, Göttingen, Germany). The SMs were stored in a freezer at −20 °C.
For AI-2 bioassays, V. harveyi MM30 was used as a reporter strain because it does not produce the AI-2 signal itself [30,33]. The bacterial culture was prepared with approximately 5 × 103 CFU/mL. Bioluminescence was measured using a microplate reader (Varioskan LUX) at 30 min intervals over 22.5 h. An impact of preparations in sub-inhibitory concentrations on C. jejuni intercellular signaling was determined indirectly from the reported bioluminescence of V. harveyi MM30 expressed in relative light units (RLU) and compared with the non-treated control (C. j. 11168-C+). To confirm that LuxS-deficient C. jejuni does not produce an AI-2 signal, we added V. harveyi MM30 culture to 5% of C. jejuni 11168∆luxS (C. j. 11168∆luxS-C+) SM and measured the bioluminescence of the reporter strain. By measuring the bioluminescence of V. harveyi MM30 without the addition of C. jejuni SM, we proved that the strain does not produce the AI-2 signal itself. Bioluminescence reduction rates (%) were calculated using equation 1. They indicate how C. jejuni AI-2-mediated intercellular signaling is reduced due to the exposure of bacteria to the preparations in comparison with the non-treated control.
To examine whether compounds of the most active fractions (i.e., PACs from Fractions 3) bind to AI-2 signal molecule, additional AI-2 bioassays were performed. In this case, we added preparations in sub-inhibitory concentrations to the initial SMs, and after 1 h incubation we measured bioluminescence of the V. harveyi MM30 reporter strain. The signals were compared with the ones from the original SMs, and bioluminescence reduction rates were calculated again. All the experiments were carried out in triplicate.
2.8. Membrane Integrity
The influence of preparations on C. jejuni membrane integrity was determined using a LIVE/DEAD BacLight Bacterial Viability kit (L-7012; Molecular Probes, Eugene, CA, USA).
The membrane disruption assays were performed according to Kovač et al. [34]. A mixture of green fluorescent dye SYTO 9 and propidium iodide (PI) was prepared according to the manufacturer instructions. Stock solutions of the extracts and their fractions were prepared in DMSO at 100-fold higher concentrations and were further diluted in MHB to the final 1% of DMSO. The dye mixture was added to 100 μL of the treated or non-treated C. jejune cultures with approximately 5 × 105 CFU/mL (1:1, v/v). The kinetics of PI intracellular penetration was measured in RFU by a microplate reader (Varioskan LUX) in terms of the SYTO 9 fluorescence at an excitation wavelength of 481 nm and an emission wavelength of 510 nm. Kinetic measurements over the last 10 min of the assays were used to calculate the membrane disruption (%). The experiment was carried out in triplicate.
2.9. Statistical Analysis
Before statistical tests, normality was tested for all the data using Shapiro–Wilk and Kolmogorov–Smirnov tests. Based on the normality test, we determined statistical significance between treatments and control using the Mann–Whitney U test in the case of non-normal distribution of the data, and using one-way ANOVA in case the data were normally distributed. At a 95% confidence interval, the results were statistically significant at value p < 0.05. The analyses were performed using IBM SPSS Statistics software, version 23 (IBM Corp., Armonk, CA, USA).
3. Results
3.1. Chemical Characterization of the Extracts and Fractions
The composition of the ethanolic extracts was qualitatively analyzed by comparing the retention time, mass spectrometric fragmentation, and UV spectra from UHPLC-PDA-ESI-MS data analysis with data from the literature [35,36,37,38,39]. Eighteen compounds were identified in the “Mattmark” ethanolic extract (Table S1, Figure S1) and seventeen in the “Rosavine” ethanolic extract (Table S2, Figure S1). Aside from typical salidroside (phenylethanoid) and rosavins (cinnamyl alcohol glycosides), chromatograms recorded at UV 270 nm indicated significant amounts of PACs as a complex mixture, of which only a few compounds could be assigned to epigallocatechin-3-O-gallate (EGCG), and two dimers containing EGCG and epigallocatechin (EGC), respectively. Two flavonoid glycosides could be assigned to the herbacetin-7-O-glycosides rhodiosin and rhodionin. The main difference between the “Mattmark” and “Rosavine” plant material is in the quantity of certain compounds or compound groups. Thus, “Mattmark” (Figure S2) contains more PACs, rosavins, and flavonoids than “Rosavine”. On the other hand, “Rosavine” (Figure S2) has higher amounts of salidroside, based on peak area comparison in UV chromatograms.
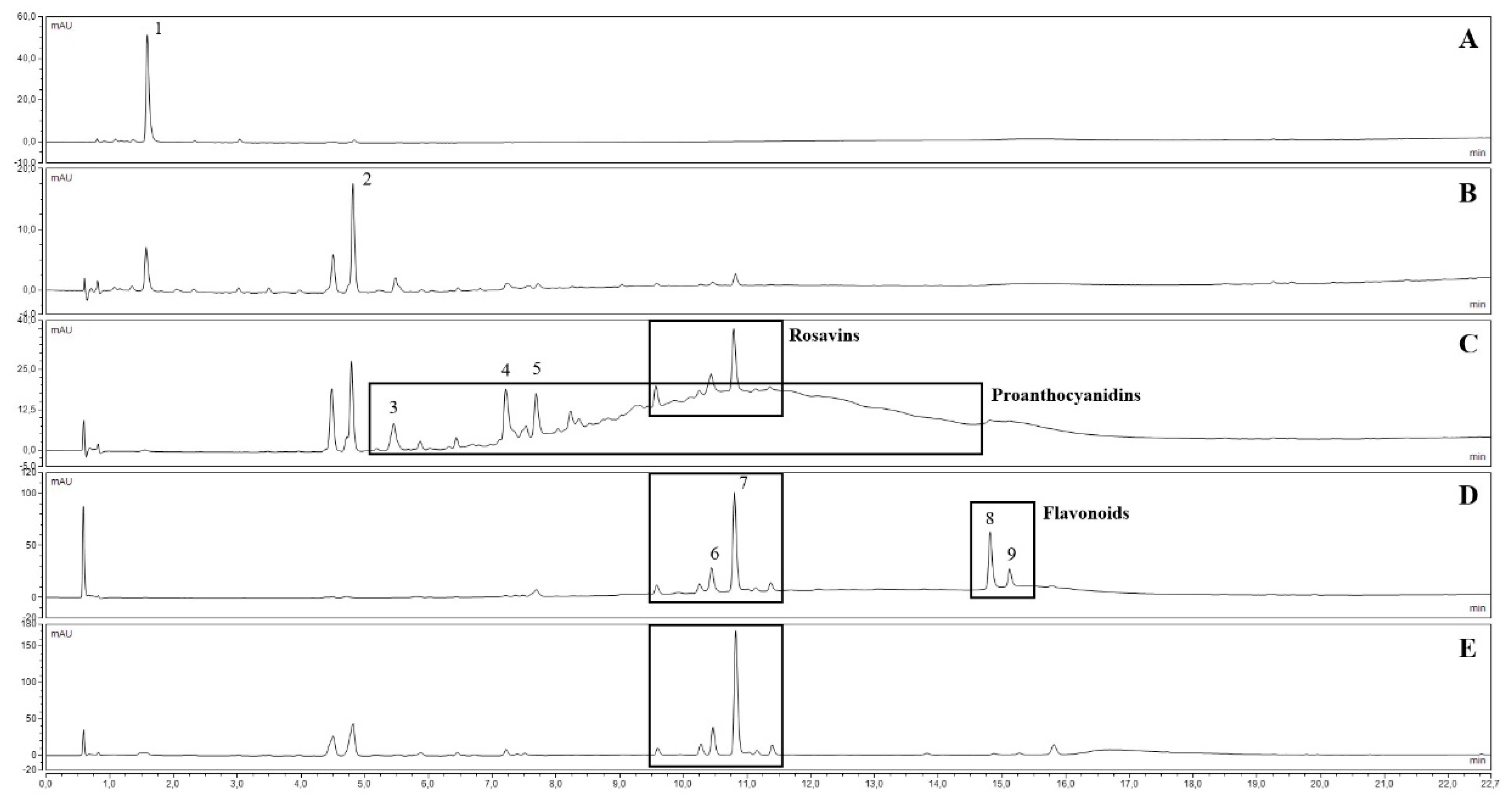
Five fractions were prepared from each crude ethanolic extract of the plant material. The main compounds or compound groups of each fraction are listed in Table 1. The composition of each fraction obtained from the “Mattmark” ethanolic extract is shown on Figure 1, and a comparison of peak areas of selected compounds is presented in Figure S3. By the adsorption of the crude ethanolic extracts to DIAION HP-20 adsorbent resin and the stepwise elution of compounds by increasing methanol concentrations, the separation of compound groups could be obtained, although salidroside could be found in Fractions F2 and Fractions F3, and rosavins could be found in Fractions F2 (trace amounts), Fractions F3, and Fractions F4. After adsorption of the crude ethanolic extracts to PVP, UHPLC analysis indicated that PACs and flavonoids, as well as gallic acid, were almost quantitatively removed from the extracts (Fractions F5).
3.2. Determination of MICs
The MICs were determined by the broth microdilution method for the reference strain C. jejuni NCTC 11168 and for the mutant strain C. jejuni 11168∆luxS (Table 2). Preparations with MICs under 1000 mg/L were tested further in sub-inhibitory concentrations, i.e., 0.25× MICs or lower, to avoid an impact on cell growth.
For C. jejuni NCTC 11168, the MIC of the “Mattmark” ethanolic extract was 125 mg/L, while the MICs of its fractions varied from 62.5 to 500 mg/L. The MIC of the “Rosavine” ethanolic extract was 500 mg/L, while the MICs of its fractions varied from over 1000 to 250 mg/L. In general, Fractions 3 and Fractions 4 showed better or the same antimicrobial activity than the ethanolic extracts, while Fractions 1, Fractions 2, and Fractions 5 were less or equally effective. The MICs of the “Mattmark” ethanolic extract and its fractions were lower compared with the MICs of the “Rosavine” ethanolic extract and its fractions.
3.3. Inhibition of Intercellular Signaling
AI-2 bioassays were performed to evaluate whether the ethanolic extracts and their fractions, in sub-inhibitory concentrations, affected C. jejuni AI-2-mediated intercellular signaling.
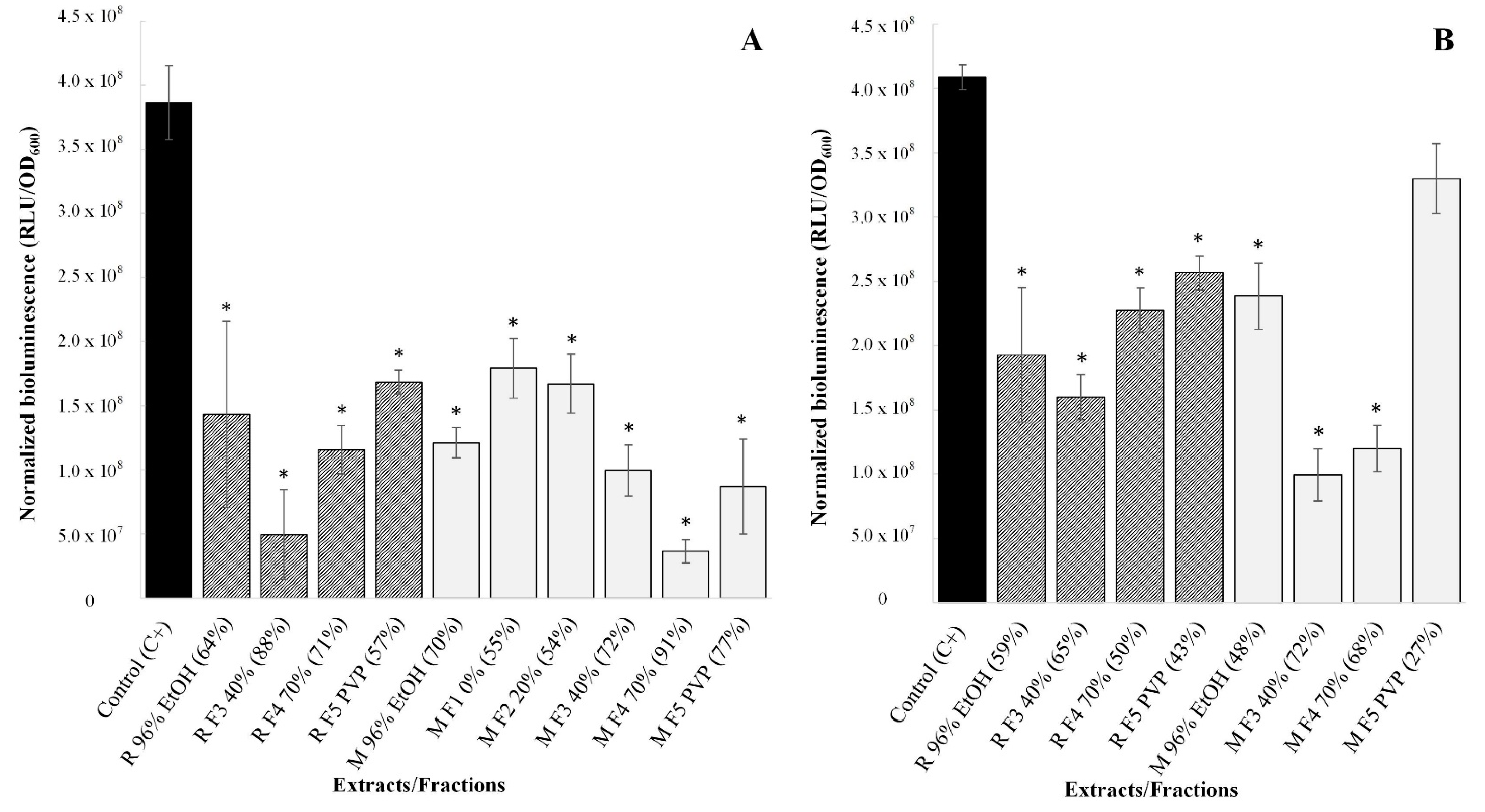
First, the impact of the ethanolic extracts and their fractions on AI-2 production in C. jejuni was tested in sub-inhibitory concentrations of 0.25× MICs. In this case, all the preparations significantly inhibited C. jejuni signaling (p < 0.05), with bioluminescence reduction rates from 54% to 91% (Figure 2A). The “Mattmark” ethanolic extract and its fractions were more effective in intercellular signaling reduction, although they were tested in lower concentrations than the “Rosavine” ethanolic extract and its fractions. Overall, R. rosea preparations showed a high impact on C. jejuni signaling but, due to the variation in tested concentrations, it was not possible to assign specific compounds or compound groups to the observed effect.
Consequently, the reduction in C. jejuni AI-2-mediated intercellular signaling by the preparations was tested at the same concentration, i.e., 15.625 mg/L, which corresponds to 0.25× MIC of “Mattmark” Fraction 3. By testing preparations at the same concentration, we aimed to find out if some extracts/fractions were more effective in reducing intercellular signaling. Both ethanolic extracts, as well as almost all their fractions, significantly affected C. jejuni signaling, even if present at such a low concentration (p < 0.05). Only “Mattmark” Fraction 5 did not have a significant impact (p > 0.05). Bioluminescence reduction rates varied from 27% to 72% (Figure 2B). Both Fractions 3 most effectively reduced C. jejuni signaling when tested at the same concentration. Additionally, Fractions 4 had a great impact. On the other hand, Fractions 5, which were devoid of PACs and flavonoids, were less effective (Figure 2B).
3.4. Disruption of Membrane Integrity
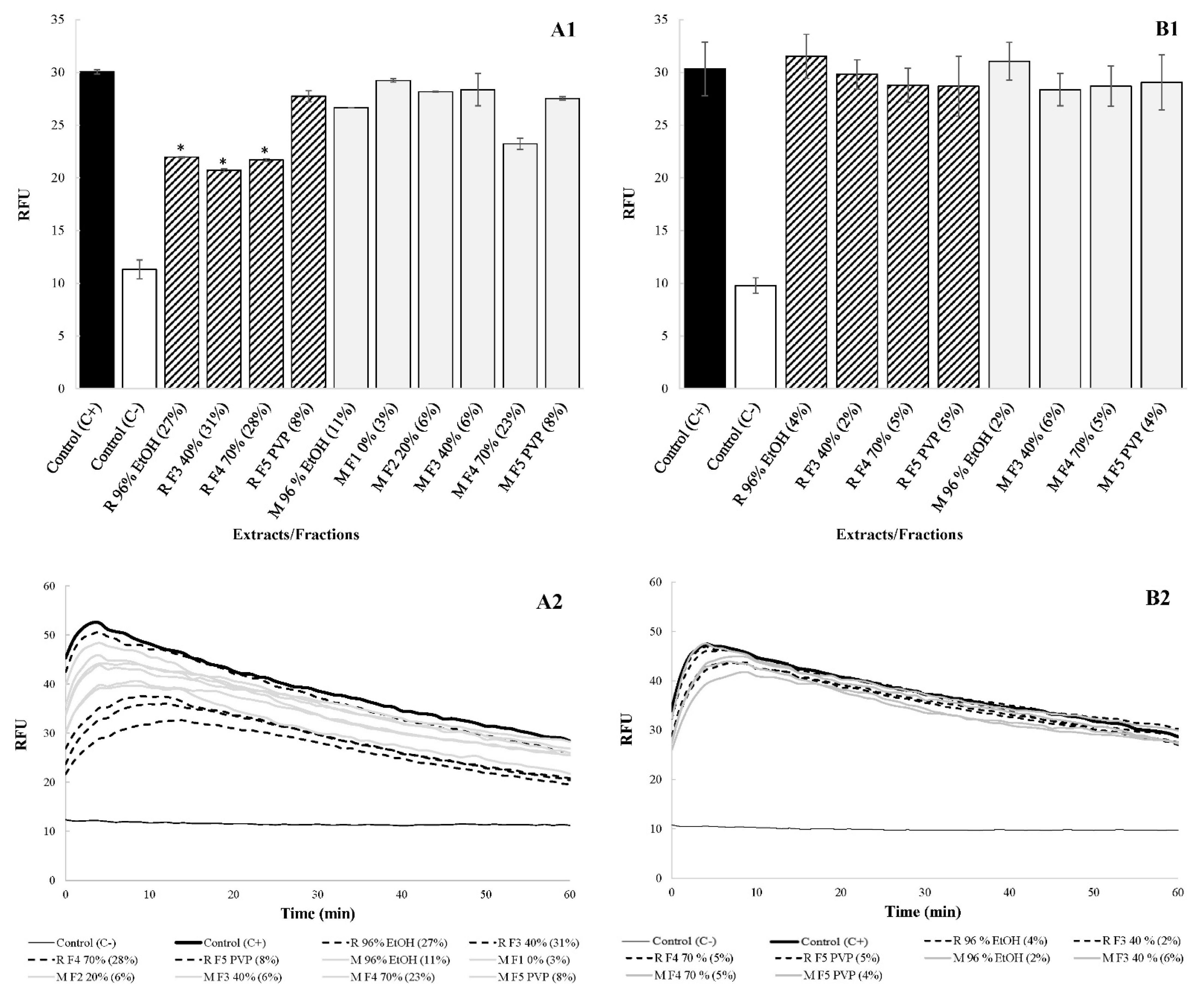
Membrane integrity assays were performed to determine if the disruption of C. jejuni membrane is also involved in the antimicrobial activity of R. rosea preparations. As in previous experiments, preparations were first tested at sub-inhibitory concentrations of 0.25× MICs, followed by testing at the same concentration, i.e., 15.625 mg/L.
At 0.25× MICs, the disruptive impact on membrane integrity varied from 3% to 31% (Figure 3(A1,A2)). In this case, the disruption of membrane integrity is likely to contribute to the antimicrobial activity of at least some of the preparations. On the other hand, when tested at the same concentration of 15.625 mg/L, the disruptive impact on membrane integrity was very low and varied from 2% to 6% (Figure 3(B1,B2)). We assume that such an effect does not contribute to the antimicrobial activity of the preparations.
4. Discussion
QS represents an important mechanism for modulating C. jejuni behavior within its population [40]. In the present study, ethanolic extracts of cultivated plant material, i.e., “Mattmark” and “Rosavine”, were prepared and fractionated by optimized protocols in order to obtain five fractions rich in salidroside, rosavins, PACs, and/or flavonoids. The fifth fractions, with almost no gallic acid, PACs, or flavonoids, were prepared from the initial ethanolic extracts to determine whether this group of compounds could play the most important role in the antimicrobial activity of R. rosea ethanolic extract.
According to the MICs, Fractions 1—rich in mainly simple gallic acid (M/R F1 0%)—and Fractions 5—without gallic acid, PACs, or flavonoids (M/R F5 PVP)—have lower antimicrobial activity, corresponding to higher MICs than the ethanolic extracts (R/M 96% EtOH). Moreover, Fractions 5 have lower antimicrobial activity than Fractions 3 (M/R F3 40%), which are rich in PACs. This indicates that PACs, including the monomeric EGCG, could be one of the crucial compound groups responsible for the antimicrobial activity of R. rosea ethanolic extract. Furthermore, flavonoids (herbacetin glycosides) might also contribute to anti-Campylobacter activity when comparing Fractions 4 and Fractions 5.
AI-2 bioassays were performed to determine the effect of the ethanolic extracts and their fractions on C. jejuni AI-2-mediated signaling. At concentrations of 0.25× MICs, both ethanolic extracts and their fractions showed a statistically significant impact (p < 0.05), with bioluminescence reduction rates ranging from 54 to 91%. Due to the variation in tested concentrations of the preparations, it was not possible to refer to the compounds or groups of compounds that play the most important role in this. Therefore, the preparations were tested at the same concentration, i.e., 15.625 mg/L, which also represents the 0.25× MIC of M F3 40%. All preparations, except R F5 PVP, significantly affected C. jejuni signaling (p < 0.05), even at such a low concentration. Here, it is important to note that M F3 40% showed a greater reduction in intercellular signaling than R F3 40%. Besides this, M F5 PVP showed much lower impact than R F5 PVP. It is also important that “Mattmark” plant material contained a higher amount of PACs and flavonoid than “Rosavine” plant material. In addition, Fractions 3 were enriched in PACs, including EGCG, while Fractions 5 had the PACs removed. These results suggest that PACs may be one of the crucial compounds in R. rosea with the potential to reduce C. jejuni signaling. High bioluminescence reduction rates (from 65% to 72%) were achieved in the case of treating C. jejuni with a sub-inhibitory concentration of Fractions 3, compared with lower impact (from 27% to 43%) when C. jejuni was treated with sub-inhibitory concentration of Fractions 5. Because M F3 40% contains more PACs than R F3 40%, a greater loss of activity was observed for “Mattmark” when we removed those compounds, as we did in Fractions 5. Similarly to MIC determination, the herbacetin glycosides rhodionin and rhodiosin could also be relevant for intercellular signaling reduction. This can be deduced from the significant reduction in bioluminescence by Fractions 4 (which were rich in these flavonoids) and the decrease in the respective effects in Fractions 5 (which were also devoid of flavonoid glycosides). The stronger intercellular signaling reduction by M F4 70% compared with R F4 70%, showing a lower concentration of flavonoids, supports the assumption that the herbacetin glycosides contribute to the observed intercellular signaling reduction of the crude extract. A high QS reduction rate (>90%) had previously been reported when C. jejuni was treated with sub-inhibitory concentrations of, e.g., C. limon [11] and R. rosea [9] ethanolic extracts. Strong anti-QS activity of EGCG from green tea against C. jejuni NCTC 11168 was previously reported by the authors of [12]. In their case, EGCG decreased the bioluminescence reported for V. harveyi BB152 of 96% when tested at a sub-inhibitory concentration of 0.75× MBC or 65.25 mg/L. Our study supports the conclusion of Šimunović et al. [9] that QS or AI-2-mediated intercellular signaling is a potential target in the control of C. jejuni and that various natural plant-based preparations act as true intercellular signaling modulators. As an upgrade to their research, we can see from our results that PACs, including EGCG and the herbacetin glycosides, are the most important compound groups of R. rosea crude ethanolic extract responsible for AI-2-mediated signaling reduction in C. jejuni.
This study also demonstrated that the antimicrobial activity of the extracts and their fractions was due to the reduction in C. jejuni signaling without affecting the membrane integrity. Even though preparations at 0.25× MICs did not affect cell growth, the membrane integrity was significantly disrupted (p > 0.05) by “Rosavine” ethanolic extract and its Fractions 3 and Fractions 4. Nevertheless, no significant effect (p < 0.05) on membrane integrity was observed compared with the non-treated control when the extracts and their fractions were tested at a concentration of 15.625 mg/L. The study also excluded that PACs affect C. jejuni signaling by binding to the AI-2 signaling molecule which was released to the growth medium. The AI-2 bioassays have shown that the reported bioluminescence of the reporter strain V. harveyi MM30 did not vary significantly (p < 0.05) between the initial SMs and the SMs to which we once again added Fractions 3, rich in PACs, at a concentration of 15.625 mg/L. For example, the bioluminescence reduction rates varied from 1% to 2% between the initial SMs obtained after treating C. jejuni with sub-inhibitory concentrations of Fractions 3 and the same SMs to which preparations were again added in sub-inhibitory concentrations (data not shown).
PACs are the most abundant plant-derived polyphenols belonging to one of the tannin groups (condensed tannins). They are among the most commonly consumed dietary polyphenols. Condensed tannins are able to form insoluble complexes with carbohydrates and proteins [41]. Studies have shown that mucosal immunity to pathogen infection can be enhanced by PACs [42]. In addition, our study supports the fact that PACs, in this case from R. rosea underground organs, represent natural compounds with the ability to reduce intercellular signaling in order to fight C. jejuni. This is in agreement with a recent publication of Hao et al. [43]: this confirmed EGCG with significant inhibitory effects on the development of biofilm, protease, elastase activity, swimming, and swarming motility, which were also positively related to the production of C4-AHL signaling molecules in Pseudomonas aeruginosa. In addition, herbacetin glycosides of R. rosea might also contribute to C. jejuni signaling reduction. Interestingly, a study by the authors of [44] showed that herbacetin has a high affinity to LuxR-type protein Shewanella baltica in a virtual screening.
5. Conclusions
In this study, we provided a protocol for the separation of bioactive compounds or compound groups from R. rosea roots and rhizomes. Our results suggest PACs and flavonoids are the most important compound groups from R. rosea, with great potential for C. jejuni AI-2-mediated signaling reduction. Nevertheless, it is still unclear whether AI-2 in C. jejuni represents a true QS signaling molecule or whether it is a metabolic by-product of a crucial central metabolic methyl cycle pathway [45]. To date, AI-2 receptors have still not been identified [46], and mechanisms by which bioactive compounds affect C. jejuni signaling should be further investigated.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/antibiotics11091220/s1: Table S1. Identified compounds in the ethanolic extract are from R. rosea “Mattmark”; Table S2. Identified compounds in the ethanolic extract are from R. rosea “Rosavine”; Figure S1. The base peak (A: “Mattmark”; B: “Rosavine”) and UV chromatograms (A1; B1: total scan; A2; B2: 276 nm; A3; B3: 360 nm) are from UHPLC-PDA-ESI-MS analysis of R. rosea ethanolic extracts; Figure S2. Comparison of peak areas at UV 270 nm of the crude ethanolic extracts of R. rosea roots “Mattmark” and “Rosavine”. Samples were analyzed at a concentration of 5 mg/mL; Figure S3. Comparison of peak areas at UV 270 nm of fractions F1 to F5 of “Mattmark” (M) and “Rosavine” (R). Samples were analyzed at a concentration of 5 mg/mL.
Author Contributions
Conceptualization, F.B. and S.S.M.; methodology, A.K.; F.B. and S.S.M.; laboratory experimental work, A.K.; validation, A.K. and F.B.; data analyses, A.K. and F.B.; writing: original manuscript, A.K.; writing: review and editing, F.B. and S.S.M.; funding acquisition, S.S.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Slovenian Research Agency (ARRS) grant P4-0116 and research project J4-2542.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are available in the Supplementary Materials.
Acknowledgments
Christoph Carlen and Claude-Alain Carron (Agroscope, Switzerland) for providing plant material; Erasmus+, European program for education, training, youth, and sport, for financial support of research stay in Graz.
Conflicts of Interest
The authors declare no conflict of interest.
References
- EFSA and ECDC. The European Union one health 2020 zoonoses report. EFSA J. 2021, 19, 6971. [Google Scholar]
- Plummer, P.J. LuxS and quorum-sensing in Campylobacter. Front. Cell. Infect. Microbiol. 2012, 2, 9. [Google Scholar] [CrossRef]
- Vasil, M.L. DNA microarrays in analysis of quorum sensing: Strengths and limitations. J. Bacteriol. 2003, 185, 2061–2065. [Google Scholar] [CrossRef]
- Deep, A.; Chaudhary, U.; Gupta, V. Quorum sensing and bacterial pathogenicity: From molecules to disease. J. Lab. Physicians. 2011, 3, 4–11. [Google Scholar] [CrossRef]
- Federle, M.J.; Bassler, B.L. Interspecies communication in bacteria. J. Clin. Investig. 2003, 112, 1291–1299. [Google Scholar] [CrossRef]
- Ng, W.L.; Bassler, B.L. Bacterial quorum-sensing network architectures. Annu. Rev. Genet. 2009, 43, 197–222. [Google Scholar] [CrossRef]
- Elvers, K.T.; Park, S.F. Quorum sensing in Campylobacter jejuni: Detection of a luxS encoded signaling molecule. Microbiology 2002, 148, 1475–1481. [Google Scholar] [CrossRef]
- Winzer, K.; Hardie, K.R.; Burgess, N.; Doherty, N.; Kirke, D.; Holden, M.T.G.; Linforth, R.; Cornell, K.A.; Taylor, A.J.; Hill, P.J.; et al. LuxS: Its role in central metabolism and the in vitro synthesis of 4-hydroxy-5-methyl-3(2H)-furanone. Microbiology 2002, 148, 909–922. [Google Scholar] [CrossRef]
- Šimunović, K.; Ramić, D.; Xu, C.; Smole Možina, S. Modulation of Campylobacter jejuni motility, adhesion to polystyrene surfaces, and invasion of INT407 cells by quorum-sensing inhibition. Microorganisms 2020, 8, 104. [Google Scholar] [CrossRef]
- Asfour, H.Z. Anti-quorum sensing natural compounds. J. Microsc. Ultrastruct. 2018, 6, 1–10. [Google Scholar] [CrossRef]
- Castillo, S.; Heredia, N.; Arechiga-Carvajal, E.; García, S. Citrus extracts as inhibitors of quorum sensing, biofilm formation and motility of Campylobacter jejuni. Food Biotechnol. 2014, 28, 106–122. [Google Scholar] [CrossRef]
- Castillo, S.; Heredia, N.; García, S. 2(5H)-Furanone, epigallocatechin gallate, and a citric-based disinfectant disturb quorum-sensing activity and reduce motility and biofilm formation of Campylobacter jejuni. Folia Microbiol. 2014, 60, 89–95. [Google Scholar] [CrossRef]
- Duarte, A.; Alves, A.C.; Ferreira, S.; Silva, F.; Domingues, F.C. Resveratrol inclusion complexes: Antibacterial and anti-biofilm activity against Campylobacter spp. and Arcobacter butzleri. Food Res. Int. 2015, 77, 244–250. [Google Scholar] [CrossRef]
- Duarte, A.; Luís, Â.; Oleastro, M.; Domingues, F.C. Antioxidant properties of coriander essential oil and linalool and their potential to control Campylobacter spp. Food Control 2016, 61, 115–122. [Google Scholar] [CrossRef]
- Bezek, K.; Kurinčič, M.; Knauder, E.; Klančnik, A.; Raspor, P.; Bucar, F.; Smole Možina, S. Attenuation of adhesion, biofilm formation and quorum sensing of Campylobacter jejuni by Euodia ruticarpa. Phytother. Res. 2016, 30, 1527–1532. [Google Scholar] [CrossRef]
- Wagle, B.R.; Donoghue, A.M.; Shrestha, S.; Upadhyaya, I.; Arsi, K.; Gupta, A.; Liyanage, R.; Rath, N.C.; Donoghue, D.J.; Upadhyay, A. Carvacrol attenuates Campylobacter jejuni colonization factors and proteome critical for persistence in the chicken gut. Poult. Sci. 2020, 99, 4566–4577. [Google Scholar] [CrossRef]
- Wagle, B.R.; Donoghue, A.M.; Jesudhasan, P.R. Select phytochemicals reduce Campylobacter jejuni in postharvest poultry and modulate the virulence attributes of C. jejuni. Front. Microbiol. 2021, 12, 725087. [Google Scholar] [CrossRef]
- Ganzera, M.; Yayla, Y.; Khan, I.A. Analysis of the marker compounds of Rhodiola rosea L. (golden root) by reversed phase high performance liquid chromatography. Chem. Pharm. Bull. 2001, 49, 465–467. [Google Scholar] [CrossRef]
- Bykov, V.A.; Zapesochnaya, G.G.; Kurkin, V.A. Traditional and biotechnological aspects of obtaining medicinal preparations from Rhodiola Rosea, L. (A review). Pharm. Chem. J. 1999, 33, 29–40. [Google Scholar] [CrossRef]
- Rodin, I.A.; Stavrianidi, A.N.; Braun, A.V.; Shpigun, O.A.; Popik, M.V. Simultaneous determination of salidroside, rosavin, and rosarin in extracts from Rhodiola rosea by high performance liquid chromatography with tandem mass spectrometry detection. J. Anal. Chem. 2012, 67, 1026–1030. [Google Scholar] [CrossRef]
- Khanum, F.; Singh Bawa, A.; Singh, B. Rhodiola rosea: A versatile adaptogen. Compr. Rev. Food Sci. Food Saf. 2005, 4, 55–62. [Google Scholar] [CrossRef]
- Perfumi, M.; Mattioli, L. Adaptogenic and central nervous system effects of single doses of 3% rosavin and 1% salidroside Rhodiola rosea L. extract in mice. Phytother. Res. 2007, 21, 37–43. [Google Scholar] [CrossRef]
- Vouillamoz, J.F.; Carron, C.A.; Malnoë, P.; Baroffio, C.A.; Carlen, C. Rhodiola rosea Mattmark, the first synthetic cultivar is launched in Switzerland. Acta Hortic. 2012, 955, 185–189. [Google Scholar] [CrossRef]
- Panossian, A.; Wikman, G.; Sarris, J. Rosenroot (Rhodiola rosea): Traditional use, chemical composition, pharmacology and clinical efficacy. Phytomedicine 2010, 17, 481–493. [Google Scholar] [CrossRef]
- Ma, C.Y.; Tang, J.; Wang, H.; Gu, X.; Tao, G.J. Simultaneous determination of six active compounds in Rhodiola, L. by RP-LC. Chromatographia 2008, 67, 383–388. [Google Scholar] [CrossRef]
- Kosakowska, O.; Bączek, K.; Przybył, J.L.; Pióro-Jabrucka, W.; Czupa, W.; Synowiec, A.; Gniewosz, M.; Costa, R.; Mondello, L.; Węglarz, Z. Antioxidant and antibacterial activity of roseroot (Rhodiola rosea L.) dry extracts. Molecules. 2018, 23, 1767. [Google Scholar] [CrossRef]
- Brown, R.P.; Gerbarg, P.L.; Ramazanov, Z. Rhodiola rosea: A phytomedicinal overview. Reliab. Herb. Med. Inf. 2002, 56, 40–52. [Google Scholar]
- Chiang, H.M.; Chen, H.C.; Wu, C.S.; Wu, P.Y.; Wen, K.C. Rhodiola plants: Chemistry and biological activity. J. Food Drug Anal. 2015, 23, 359–369. [Google Scholar] [CrossRef]
- Kelly, G.S. Rhodiola rosea: A possible plant adaptogen. Altern. Med. Rev. 2001, 6, 293–302. [Google Scholar]
- Surette, M.G.; Miller, M.B.; Bassler, B.L. Quorum sensing in Escherichia coli, Salmonella typhimurium, and Vibrio harveyi: A new family of genes responsible for autoinducer production. Proc. Natl. Acad. Sci. USA 1999, 96, 1639–1644. [Google Scholar] [CrossRef]
- Sun, L.; Zhou, R.; Sui, J.; Liu, Y. Simultaneous preparation of Salidroside and p-Tyrosol from Rhodiola crenulata by DIAION HP-20 macroporous resin chromatography combined with silica gel chromatography. Molecules 2018, 23, 1602. [Google Scholar] [CrossRef]
- Klančnik, A.; Piskernik, S.; Jeršek, B.; Smole Možina, S. Evaluation of diffusion and dilution methods to determine the antibacterial activity of plant extracts. J. Microbiol. Methods 2010, 81, 121–126. [Google Scholar] [CrossRef]
- Bassler, B.L.; Greenberg, E.P.; Stevens, A.M. Cross-species induction of luminescence on the quorum-sensing bacterium Vibrio harveyi. J. Bacteriol. 1997, 179, 4043–4045. [Google Scholar] [CrossRef] [Green Version]
- Kovač, J.; Šimunović, K.; Wu, Z.; Klančnik, A.; Bucar, F.; Zhang, Q.; Smole Možina, S. Antibiotic resistance modulation and modes of action of (-)-α-pinene in Campylobacter jejuni. PLoS ONE 2015, 10, e0122871. [Google Scholar]
- Alperth, F.; Turek, I.; Weiss, S.; Vogt, D.; Bucar, F. Qualitative and quantitative analysis of different Rhodiola rosea rhizome extracts by UHPLC-DAD-ESI-MSn. Sci. Pharm. 2019, 87, 8. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Chirikova, N.K.; Vasilieva, A.G.; Fedorov, I.A. LC-MS profile, gastrointestinal and gut microbiota stability and antioxidant activity of Rhodiola rosea herb metabolites: A comparative study with subterranean organs. Antioxidants 2020, 9, 526. [Google Scholar] [CrossRef]
- Petsalo, A.; Jalonen, J.; Tolonen, A. Identification of flavonoids of Rhodiola rosea by liquid chromatography-tandem mass spectrometry. J. Chromatogr. A 2006, 1112, 224–231. [Google Scholar] [CrossRef]
- Zapesochnaya, G.G.; Kurkin, V.A. The flavonoids of the rhizomes of Rhodiola rosea. Plenum Publ. Corp. 1983, 19, 21–29. [Google Scholar]
- Zomborszki, Z.P.; Kúsz, N.; Csupor, D.; Peschel, W. Rhodiosin and herbacetin in Rhodiola rosea preparations: Additional markers for quality control? Pharm. Biol. 2019, 57, 295–305. [Google Scholar] [CrossRef]
- Teren, M.; Turonova Michova, H.; Vondrakova, L.; Demnerova, K. Molecules autoinducer 2 and cjA and their impact on gene expression in Campylobacter jejuni. J. Mol. Microbiol. Biotechnol. 2019, 28, 207–215. [Google Scholar] [CrossRef]
- Balalaie, A.; Rezvani, M.B.; Mohammadi Basir, M. Dual function of proanthocyanidins as both MMP inhibitor and crosslinker in dentin biomodification: A literature review. Dent. Mater. J. 2018, 37, 173–182. [Google Scholar] [CrossRef]
- Andersen-Civil, A.I.S.; Arora, P.; Williams, A.R. Regulation of enteric infection and immunity by dietary proanthocyanidins. Front. Immunol. 2021, 12, 637603. [Google Scholar] [CrossRef]
- Hao, S.; Yang, D.; Zhao, L.; Shi, F.; Ye, G.; Fu, H.; Lin, J.; Guo, H.; He, R.; Li, J.; et al. EGCG-mediated potential inhibition of biofilm development and quorum sensing in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2021, 22, 4946. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Chen, J.; Koseki, S.; Yang, Q.; Yu, H.; Fu, L. Screening and preservation application of quorum sensing inhibitors of Pseudomonas fluorescens and Shewanella baltica in seafood products. LWT 2021, 149, 111749. [Google Scholar] [CrossRef]
- Ramić, D.; Ogrizek, J.; Bucar, F.; Jeršek, B.; Jeršek, M.; Smole Možina, S. Campylobacter jejuni biofilm control with Lavandin essential oils and by-products. Antibiotics 2022, 11, 854. [Google Scholar] [CrossRef]
- Püning, C.; Su, Y.; Lu, X.; Gölz, G. Molecular mechanisms of Campylobacter biofilm formation and quorum sensing. In Fighting Campylobacter Infections. Current Topics in Microbiology and Immunology; Backert, S., Ed.; Springer: Cham, Switzerland, 2021; Volume 431, pp. 293–319. [Google Scholar]
Figure 1.
UV chromatograms at 270 nm of “Mattmark” fractions (A): Fraction 1 (M F1 0%); (B): Fraction 2 (M F2 20%); (C): Fraction 3 (M F3 40%); (D): Fraction 4 (M F4 70%); (E): Fraction 5 (M F5 PVP)). Compounds: 1 = gallic acid, 2 = salidroside, 3 = EGC-EGCG dimer, 4 = EGCG-EGCG dimer, 5 = EGCG, 6 = rosarin, 7 = rosavin, 8 = rhodiosin, 9 = rhodionin; for details see Table S1.
Figure 1.
UV chromatograms at 270 nm of “Mattmark” fractions (A): Fraction 1 (M F1 0%); (B): Fraction 2 (M F2 20%); (C): Fraction 3 (M F3 40%); (D): Fraction 4 (M F4 70%); (E): Fraction 5 (M F5 PVP)). Compounds: 1 = gallic acid, 2 = salidroside, 3 = EGC-EGCG dimer, 4 = EGCG-EGCG dimer, 5 = EGCG, 6 = rosarin, 7 = rosavin, 8 = rhodiosin, 9 = rhodionin; for details see Table S1.

Figure 2.
Bioluminescence reported after the addition of V. harveyi MM30 to 5% of SM obtained from non-treated control (C+) or to 5% of SMs obtained after treating C. jejuni NCTC 11168 with different preparations in sub-inhibitory concentrations ((A): 0.25× MIC; (B): 15.625 mg/L). Average bioluminescence in RLU with deducted background and normalized to OD600 are presented ± standard deviations. In addition, the bioluminescence reduction rate is presented in brackets. * represents statistically significant values.
Figure 2.
Bioluminescence reported after the addition of V. harveyi MM30 to 5% of SM obtained from non-treated control (C+) or to 5% of SMs obtained after treating C. jejuni NCTC 11168 with different preparations in sub-inhibitory concentrations ((A): 0.25× MIC; (B): 15.625 mg/L). Average bioluminescence in RLU with deducted background and normalized to OD600 are presented ± standard deviations. In addition, the bioluminescence reduction rate is presented in brackets. * represents statistically significant values.

Figure 3.
Influence of the extracts and their fractions on C. jejuni NCTC 11168 membrane integrity ((A1/A2): preparations tested at 0.25× MICs; (B1/B2): preparations tested in the same concentration, i.e., 15.625 mg/L). An impact on membrane integrity in RFU is presented ± standard deviations. In addition, the decrease in membrane integrity is presented in brackets. * represents statistically significant values.
Figure 3.
Influence of the extracts and their fractions on C. jejuni NCTC 11168 membrane integrity ((A1/A2): preparations tested at 0.25× MICs; (B1/B2): preparations tested in the same concentration, i.e., 15.625 mg/L). An impact on membrane integrity in RFU is presented ± standard deviations. In addition, the decrease in membrane integrity is presented in brackets. * represents statistically significant values.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Fractions obtained from “Mattmark” and “Rosavine” ethanolic extracts and their main compounds.
Table 1.
Fractions obtained from “Mattmark” and “Rosavine” ethanolic extracts and their main compounds.
| No. | Label | Solvent | Main Compounds/Compound Groups |
|---|---|---|---|
| 1 | M/R F1 0% | MW, 0% methanol | Rich in gallic acid (Figure 1A) |
| 2 | M/R F2 20% | 20% methanol | Rich in salidroside (Figure 1B) |
| 3 | M/R F3 40% | 40% methanol | Rich in salidroside, rosavins, and PACs (Figure 1C) |
| 4 | M/R F4 70% | 70% methanol | Rich in rosavins and flavonoids (Figure 1D) |
| 5 | M/R F5 PVP | 50% methanol | Rich in salidroside and rosavins (contains almost no PACs or flavonoids) (Figure 1E) |
Table 2.
Determined MICs of ethanolic extracts and their fractions for C. jejuni NCTC 11168; C. jejuni 11168∆luxS.
Table 2.
Determined MICs of ethanolic extracts and their fractions for C. jejuni NCTC 11168; C. jejuni 11168∆luxS.
| Extracts/Fractions | C. jejuni NCTC 11168 | C. jejuni 11168∆luxS |
|---|---|---|
| MIC (mg/L) | ||
| “Mattmark” Plant Material | ||
| M 96% EtOH | 125 | 125 |
| M F1 0% | 250 | 250 |
| M F2 20% | 125 | 125 |
| M F3 40% | 62.5 | 62.5 |
| M F4 70% | 125 | 125 |
| M F5 PVP | 250 | 250 |
| “Rosavine” Plant Material | ||
| R 96% EtOH | 500 | 250 |
| R F1 0% | >1000 | 1000 |
| R F2 20% | >1000 | 500 |
| R F3 40% | 250 | 62.5 |
| R F4 70% | 250 | 125 |
| R F5 PVP | 1000 | 500 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kunčič, A.; Bucar, F.; Smole Možina, S. Rhodiola rosea Reduces Intercellular Signaling in Campylobacter jejuni. Antibiotics 2022, 11, 1220. https://doi.org/10.3390/antibiotics11091220
AMA Style
Kunčič A, Bucar F, Smole Možina S. Rhodiola rosea Reduces Intercellular Signaling in Campylobacter jejuni. Antibiotics. 2022; 11(9):1220. https://doi.org/10.3390/antibiotics11091220
Chicago/Turabian StyleKunčič, Ajda, Franz Bucar, and Sonja Smole Možina. 2022. "Rhodiola rosea Reduces Intercellular Signaling in Campylobacter jejuni" Antibiotics 11, no. 9: 1220. https://doi.org/10.3390/antibiotics11091220
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.