
Matcha Green Tea Exhibits Bactericidal Activity against Streptococcus pneumoniae and Inhibits Functional Pneumolysin

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
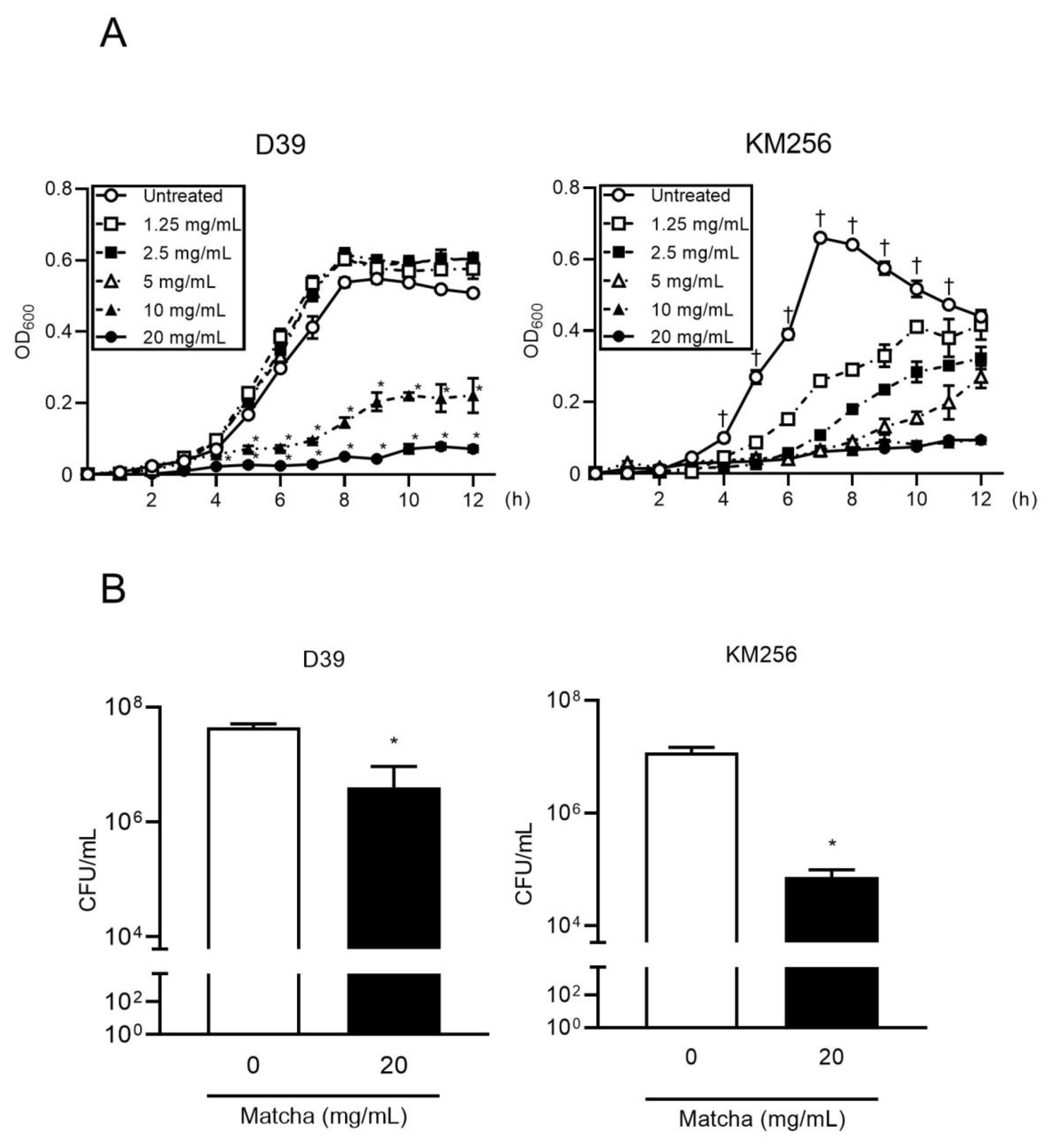
2.1. Matcha Supernatants Exhibit Antibacterial Activity against Streptococcus pneumoniae
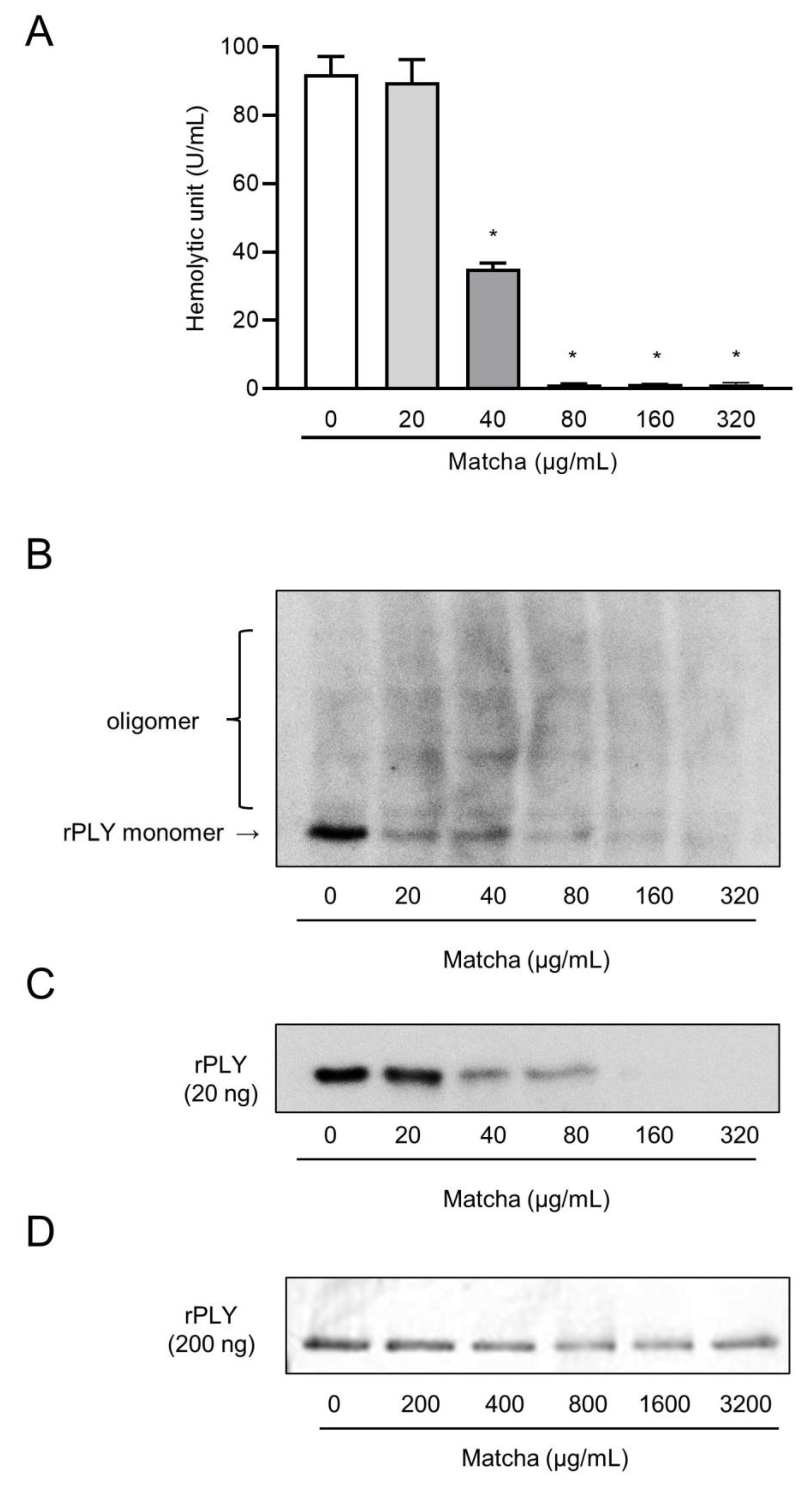
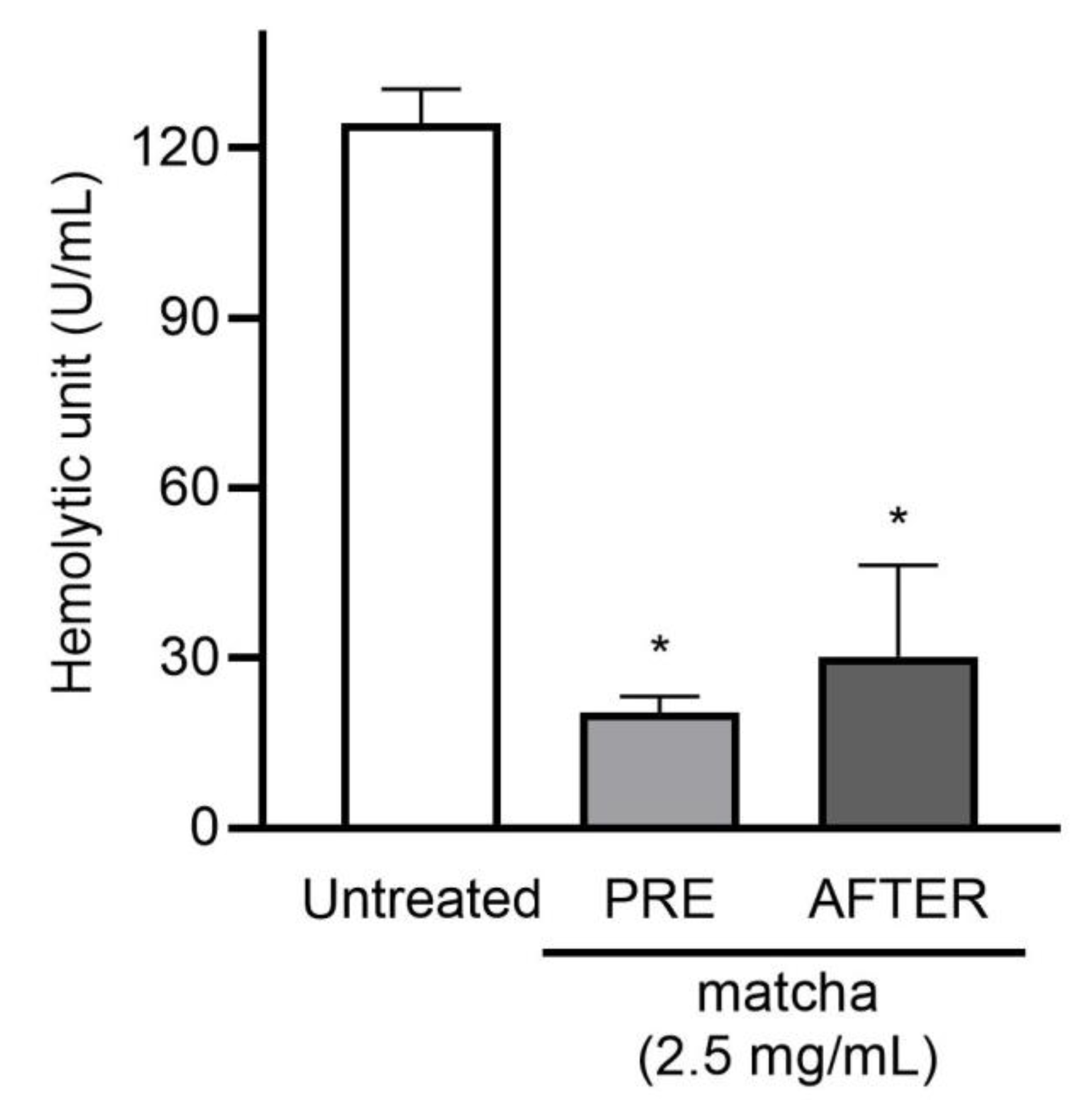
2.2. Matcha Supernatants Decrease Heolytic Activity of Recombinant PLY
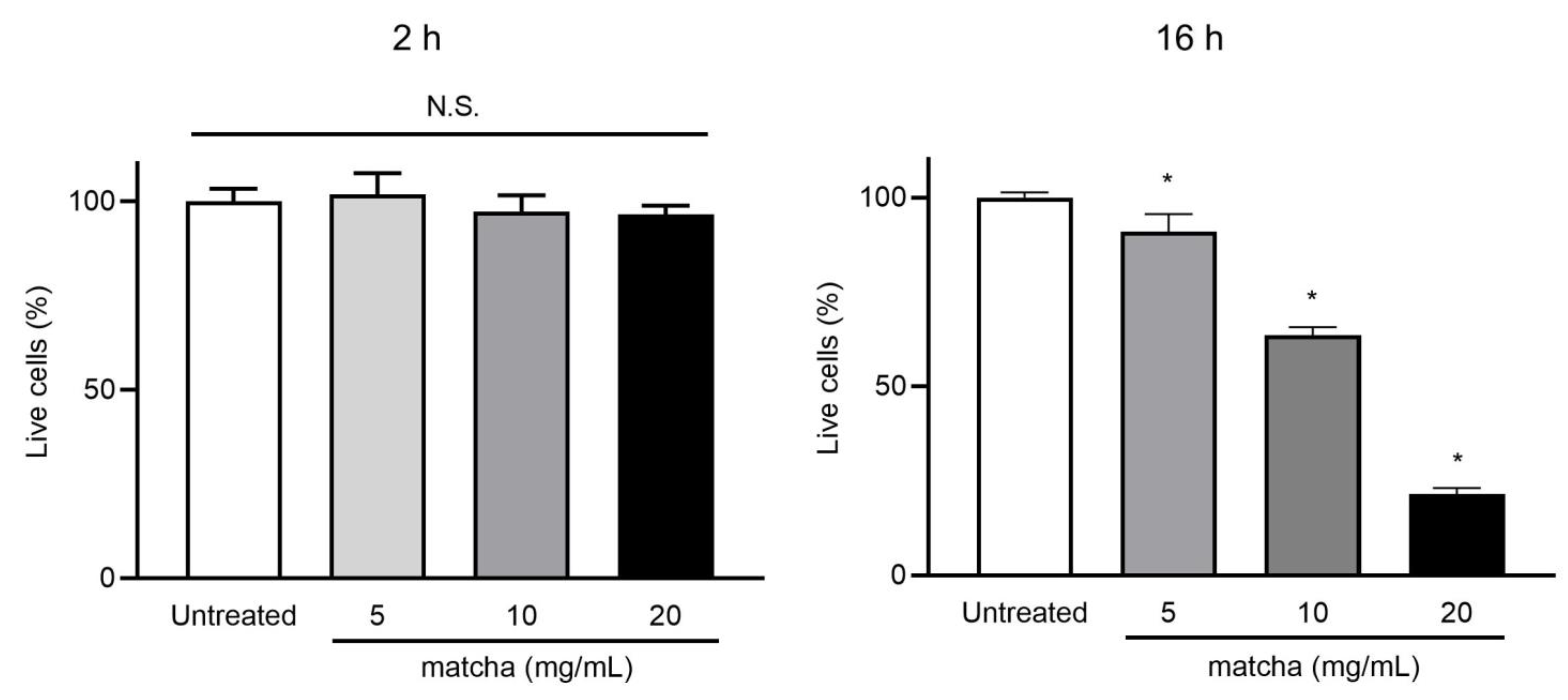
2.3. Matcha Supernatants Do Not Exhibit Cytotoxicity toward A549 Cells up to 5 mg/mL
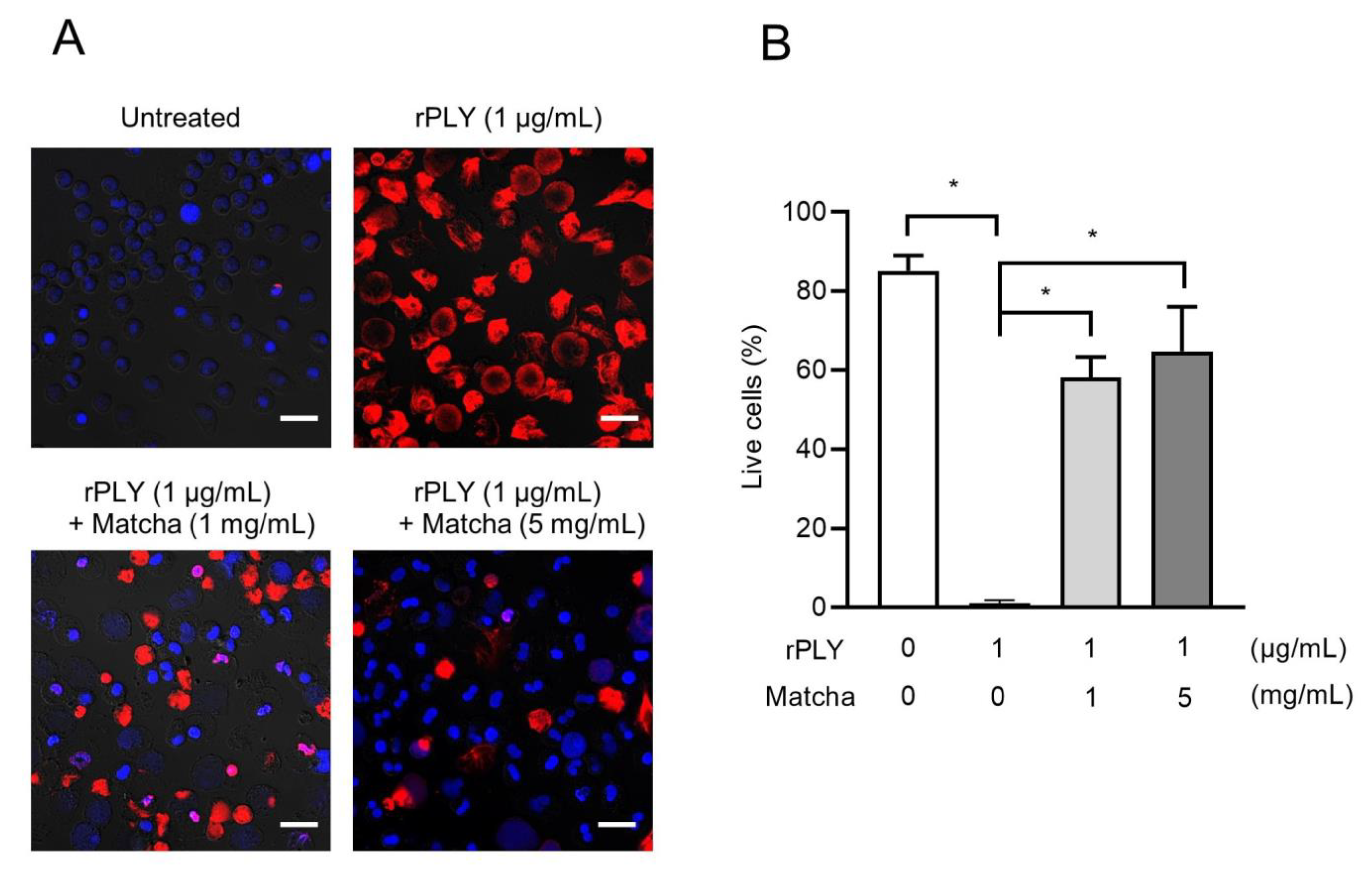
2.4. Matcha Supernatants Inhibit rPLY-Induced Neutrophil Death
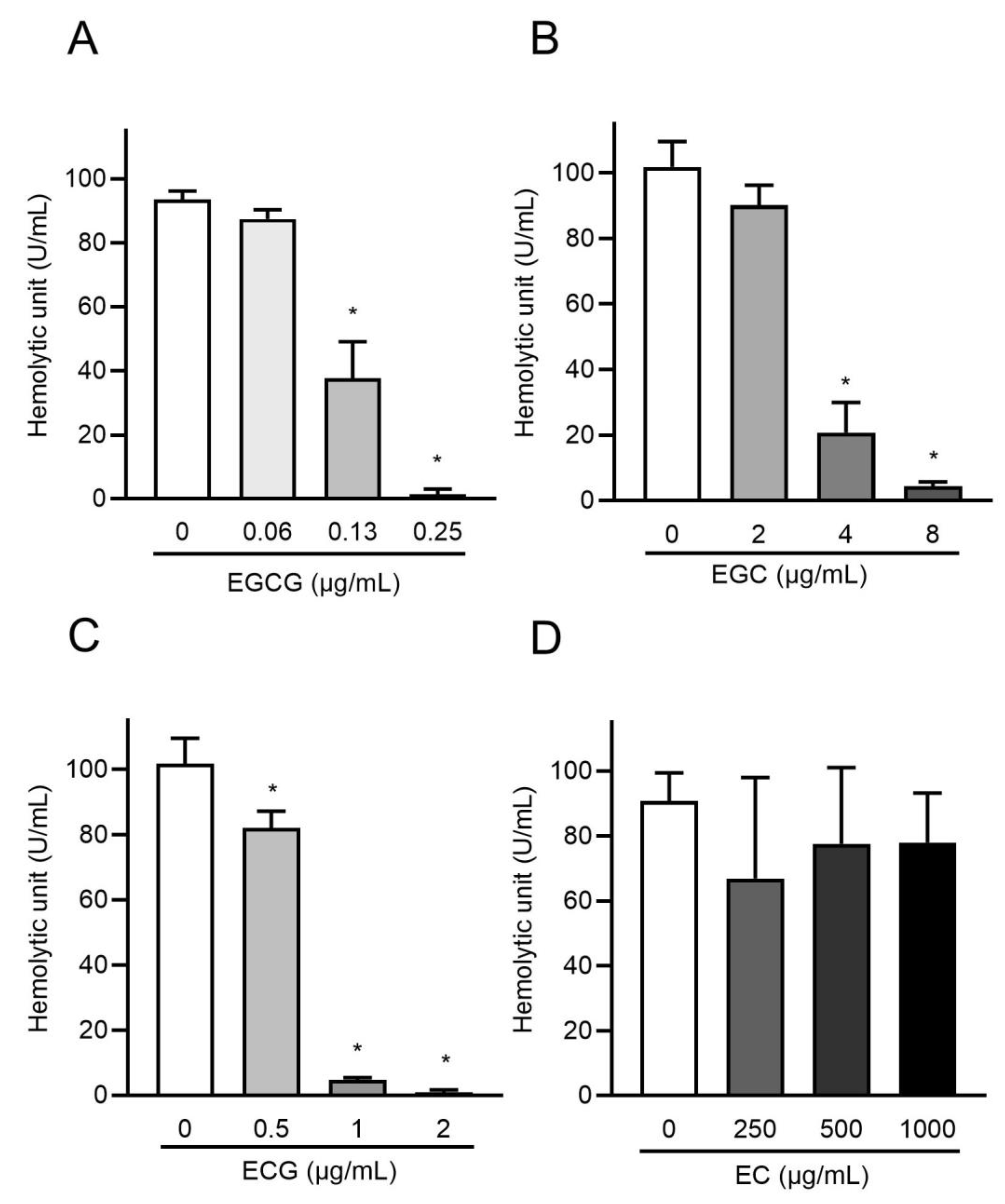
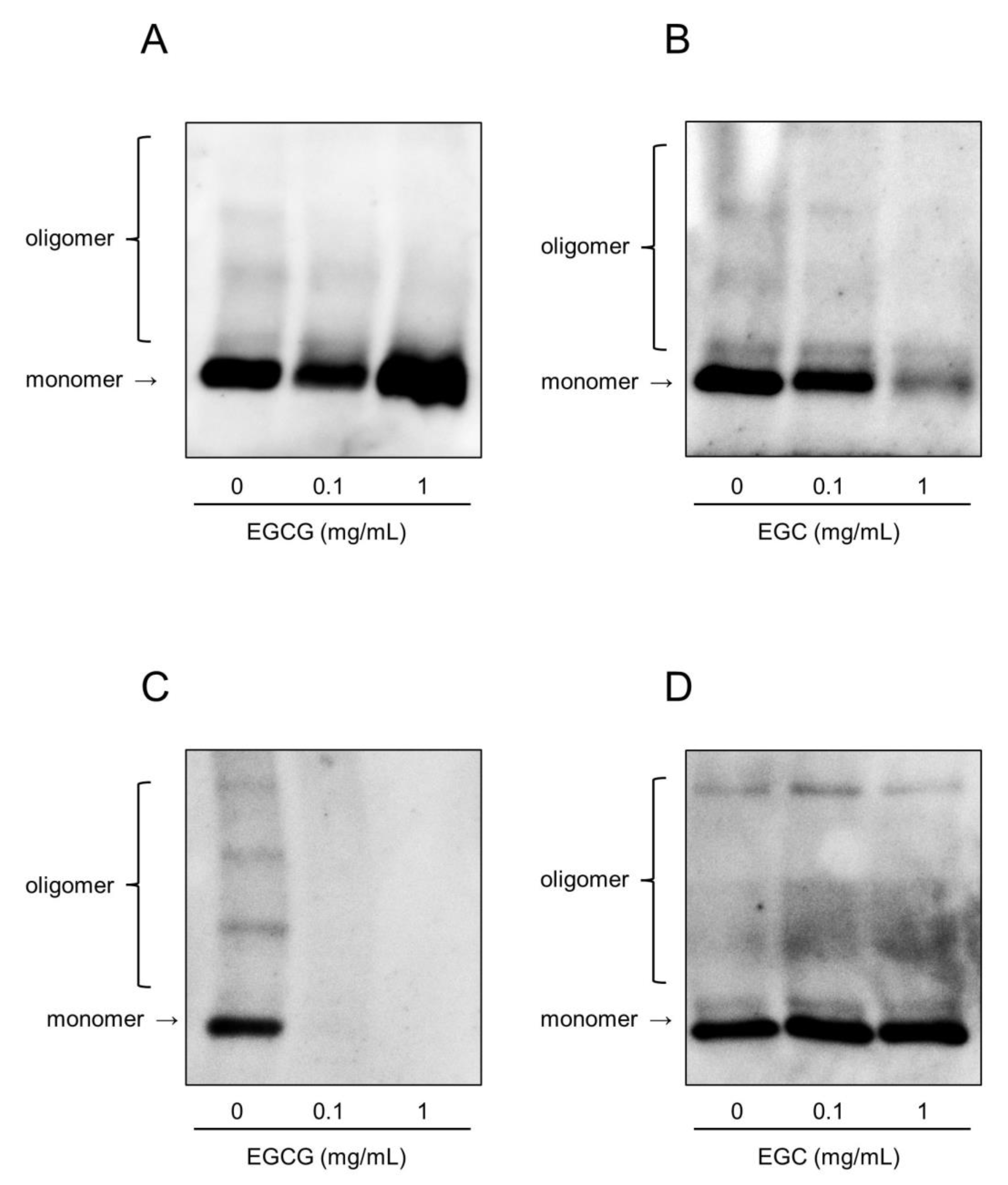
2.5. Catechins Suppress the Hemolytic Activity of rPLY by Inhibiting Its Oligomerization
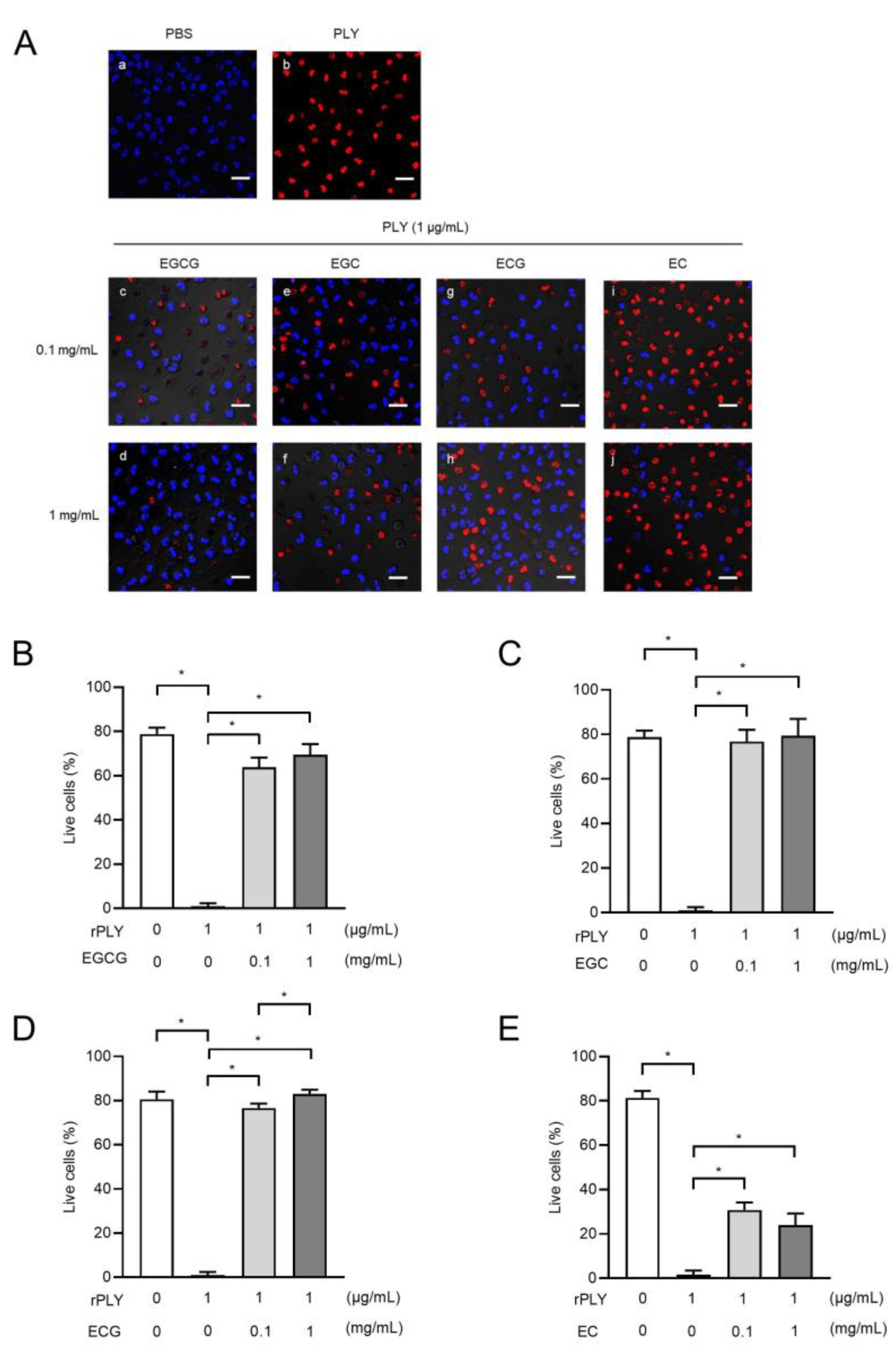
2.6. Catechins Suppress rPLY-Induced Loss of Neutrophil Viability
2.7. Catechins Suppress the Release of Neutrophil Elastase in Response to rPLY Stimulation
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Reagents
4.2. Effect of Matcha Supernatant on the Growth of S. pneumoniae
4.3. Hemolytic Assay
4.4. Human Neutrophil Isolation
4.5. Cytotoxic Assay
4.6. Western Blotting Assay
4.7. Protein Staining
4.8. Neutrophil Elastase Activity Assay
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Supplementary Materials and Methods
Appendix A.1.1. Bacterial Strains and Reagents
Appendix A.1.2. Hemolytic Assay
Appendix A.1.3. Cell Line
Appendix A.1.4. Cytotoxicity Assay
Appendix B


References
- Bogaert, D.; De Groot, R.; Hermans, P.W. Streptococcus pneumoniae colonization: The key to pneumococcal disease. Lancet Infect. Dis. 2004, 4, 144–154. [Google Scholar] [CrossRef]
- Appelbaum, P.C. Resistance among Streptococcus pneumoniae: Implications for drug selection. Clin. Infect. Dis. 2002, 34, 1613–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagai, K.; Kimura, O.; Domon, H.; Maekawa, T.; Yonezawa, D.; Terao, Y. Antimicrobial susceptibility of Streptococcus pneumoniae, Haemophilus influenzae, and Moraxella catarrhalis clinical isolates from children with acute otitis media in Japan from 2014 to 2017. J. Infect. Chemother. 2019, 25, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Alhamdi, Y.; Neill, D.R.; Abrams, S.T.; Malak, H.A.; Yahya, R.; Barrett-Jolley, R.; Wang, G.; Kadioglu, A.; Toh, C.H. Circulating pneumolysin is a potent inducer of cardiac injury during pneumococcal infection. PLoS Pathog. 2015, 11, e1004836. [Google Scholar] [CrossRef] [Green Version]
- Tabata, A.; Nagamune, H. Diversity of β-hemolysins produced by the human opportunistic streptococci. Microbiol. Immunol. 2021, 65, 512–529. [Google Scholar] [CrossRef] [PubMed]
- Berry, A.M.; Ogunniyi, A.D.; Miller, D.C.; Paton, J.C. Comparative virulence of Streptococcus pneumoniae strains with insertion-duplication, point, and deletion mutations in the pneumolysin gene. Infect. Immun. 1999, 67, 981–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, J.E.; Faraj, B.H.; Gingras, A.R.; Lonnen, R.; Sheikh, M.A.; El-Mezgueldi, M.; Moody, P.C.; Andrew, P.W.; Wallis, R. The crystal structure of pneumolysin at 2.0-A resolution reveals the molecular packing of the pre-pore complex. Sci. Rep. 2015, 5, 13293. [Google Scholar] [CrossRef] [Green Version]
- Palmer, M. The family of thiol-activated, cholesterol-binding cytolysins. Toxicon 2001, 39, 1681–1689. [Google Scholar] [CrossRef]
- Nishimoto, A.T.; Rosch, J.W.; Tuomanen, E.I. Pneumolysin: Pathogenesis and therapeutic target. Front. Microbiol. 2020, 11, 1543. [Google Scholar] [CrossRef] [PubMed]
- Alouf, J.E. Cholesterol-binding cytolytic protein toxins. Int. J. Med. Microbiol. 2000, 290, 351–356. [Google Scholar] [CrossRef]
- Domon, H.; Oda, M.; Maekawa, T.; Nagai, K.; Takeda, W.; Terao, Y. Streptococcus pneumoniae disrupts pulmonary immune defence via elastase release following pneumolysin-dependent neutrophil lysis. Sci. Rep. 2016, 6, 38013. [Google Scholar] [CrossRef] [PubMed]
- Holmlund, E.; Quiambao, B.; Ollgren, J.; Nohynek, H.; Käyhty, H. Development of natural antibodies to pneumococcal surface protein A, pneumococcal surface adhesin A and pneumolysin in Filipino pregnant women and their infants in relation to pneumococcal carriage. Vaccine 2006, 24, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Domon, H.; Maekawa, T.; Yonezawa, D.; Nagai, K.; Oda, M.; Yanagihara, K.; Terao, Y. Mechanism of macrolide-induced inhibition of pneumolysin release involves impairment of autolysin release in macrolide-resistant Streptococcus pneumoniae. Antimicrob. Agents Chemother. 2018, 62, e00161-18. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.; Steel, H.C.; Cockeran, R.; Smith, A.M.; von Gottberg, A.; de Gouveia, L.; Brink, A.; Klugman, K.P.; Mitchell, T.J.; Feldman, C. Clarithromycin alone and in combination with ceftriaxone inhibits the production of pneumolysin by both macrolide-susceptible and macrolide-resistant strains of Streptococcus pneumoniae. J. Antimicrob. Chemother. 2007, 59, 224–229. [Google Scholar] [CrossRef] [Green Version]
- Domon, H.; Isono, T.; Hiyoshi, T.; Tamura, H.; Sasagawa, K.; Maekawa, T.; Hirayama, S.; Yanagihara, K.; Terao, Y. Clarithromycin Inhibits pneumolysin Production via Downregulation of ply Gene Transcription despite Autolysis Activation. Microbiol. Spectr. 2021, 9, e0031821. [Google Scholar] [CrossRef] [PubMed]
- Rosch, J.W.; Boyd, A.R.; Hinojosa, E.; Pestina, T.; Hu, Y.; Persons, D.A.; Orihuela, C.J.; Tuomanen, E.I. Statins protect against fulminant pneumococcal infection and cytolysin toxicity in a mouse model of sickle cell disease. J. Clin. Investig. 2010, 120, 627–635. [Google Scholar] [CrossRef] [Green Version]
- Li, H.E.; Zhao, X.R.; Deng, X.M.; Wang, J.F.; Song, M.; Niu, X.D.; Peng, L.P. Insights into structure and activity of natural compound inhibitors of pneumolysin. Sci Rep-Uk. Sci. Rep. 2017, 7, 42015. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Zhou, Y.; Wang, G.; Shi, D.; Zha, Y.; Yi, P.; Wang, J. Morin moderates the biotoxicity of pneumococcal pneumolysin by weakening the oligomers’ formation. Chem. Pharm. Bull. 2017, 65, 538–544. [Google Scholar] [CrossRef] [Green Version]
- Maatsola, S.; Kurkinen, S.; Engström, M.T.; Nyholm, T.K.M.; Pentikäinen, O.; Salminen, J.P.; Haataja, S. Inhibition of pneumolysin cytotoxicity by hydrolysable tannins. Antibiotics 2020, 9, 930. [Google Scholar] [CrossRef]
- Kochman, J.; Jakubczyk, K.; Antoniewicz, J.; Mruk, H.; Janda, K. Health benefits and chemical composition of Matcha green tea: A review. Molecules 2020, 26, 85. [Google Scholar] [CrossRef]
- Serafini, M.; Del Rio, D.; Yao, D.N.D.; Bettuzzi, S.; Peluso, I. Health Benefits of Tea, 2nd ed.; CRC Press: Boca Raton, FL, USA; Taylor & Francis: Abingdon, UK, 2011. [Google Scholar]
- Watanabe, I.; Kuriyama, S.; Kakizaki, M.; Sone, T.; Ohmori-Matsuda, K.; Nakaya, N.; Hozawa, A.; Tsuji, I. Green tea and death from pneumonia in Japan: The Ohsaki cohort study. Am. J. Clin. Nutr. 2009, 90, 672–679. [Google Scholar] [CrossRef] [Green Version]
- Umeda, M.; Tominaga, T.; Kozuma, K.; Kitazawa, H.; Furushima, D.; Hibi, M.; Yamada, H. Preventive effects of tea and tea catechins against influenza and acute upper respiratory tract infections: A systematic review and meta-analysis. Eur. J. Nutr. 2021, 60, 4189–4202. [Google Scholar] [CrossRef]
- Horie, H.; Ema, K.; Sumikawa, O. Chemical components of Matcha and powdered green tea. J. Cook Sci. Jpn. 2017, 50, 182–188. [Google Scholar]
- Dietz, C.; Dekker, M.; Piqueras-Fiszman, B. An intervention study on the effect of matcha tea, in drink and snack bar formats, on mood and cognitive performance. Food Res. Int. 2017, 99, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Reygaert, W.C. Green tea catechins: Their use in treating and preventing infectious diseases. BioMed Res. Int. 2018, 2018, 9105261. [Google Scholar] [CrossRef]
- Iwasaki, M.; Inoue, M.; Sasazuki, S.; Sawada, N.; Yamaji, T.; Shimazu, T.; Willett, W.C.; Tsugane, S.; Japan Public Health Center-Based Prospective Study Group. Green tea drinking and subsequent risk of breast cancer in a population-based cohort of Japanese women. Breast Cancer Res. 2010, 12, R88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, D.J.; Anderton, C.R. Determination of catechins in matcha green tea by micellar electrokinetic chromatography. J. Chromatogr. A 2003, 1011, 173–180. [Google Scholar] [CrossRef]
- Cabrera, C.; Artacho, R.; Gimenez, R. Beneficial effects of green tea—A review. J. Am. Coll. Nutr. 2006, 25, 79–99. [Google Scholar] [CrossRef] [PubMed]
- Weil, A.; Daley, R. The Healthy Kitchen: Recipes for a Better Body, Life, and Spirit; Knopf: New York, NY, USA, 2003. [Google Scholar]
- Song, M.; Teng, Z.; Li, M.; Niu, X.; Wang, J.; Deng, X. Epigallocatechin gallate inhibits Streptococcus pneumoniae virulence by simultaneously targeting pneumolysin and sortase A. J. Cell Mol. Med. 2017, 21, 2586–2598. [Google Scholar] [CrossRef] [PubMed]
- Chang, E.H.; Huang, J.; Lin, Z.; Brown, A.C. Catechin-mediated restructuring of a bacterial toxin inhibits activity. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 191–198. [Google Scholar] [CrossRef]
- Karim, A.J.; Dalai, D.R. Green tea: A review on its natural anti-oxidant therapy and cariostatic benefits. J. Issues ISSN 2014, 2350, 1588. [Google Scholar]
- Musial, C.; Kuban-Jankowska, A.; Gorska-Ponikowska, M. Beneficial properties of green tea catechins. Int. J. Mol. Sci. 2020, 21, 1744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domon, H.; Terao, Y. The role of neutrophils and neutrophil elastase in pneumococcal pneumonia. Front. Cell Infect. Microbiol. 2021, 11, 615959. [Google Scholar] [CrossRef] [PubMed]
- Reygaert, W.C. The antimicrobial possibilities of green tea. Front. Microbiol. 2014, 5, 434. [Google Scholar] [CrossRef]
- Xu, J.; Xu, Z.; Zheng, W. A review of the antiviral role of green tea catechins. Molecules 2017, 22, 1337. [Google Scholar] [CrossRef] [Green Version]
- Umashankar, N.; Pemmanda, B.; Gopkumar, P.; Hemalatha, A.J.; Sundar, P.K.; Prashanth, H.V. Effectiveness of topical green tea against multidrug-resistant Staphylococcus aureus in cases of primary pyoderma: An open controlled trial. Indian J. Dermatol. Venereol. Leprol. 2018, 84, 163–168. [Google Scholar] [CrossRef]
- Reygaert, W.; Jusufi, I. Green tea as an effective antimicrobial for urinary tract infections caused by Escherichia coli. Front. Microbiol. 2013, 4, 162. [Google Scholar] [CrossRef] [Green Version]
- Sirk, T.W.; Brown, E.F.; Friedman, M.; Sum, A.K. Molecular binding of catechins to biomembranes: Relationship to biological activity. J. Agric. Food Chem. 2009, 57, 6720–6728. [Google Scholar] [CrossRef]
- Wu, M.; Brown, A.C. Applications of catechins in the treatment of bacterial infections. Pathogens 2021, 10, 546. [Google Scholar] [CrossRef] [PubMed]
- Rubins, J.B.; Charboneau, D.; Paton, J.C.; Mitchell, T.J.; Andrew, P.W.; Janoff, E.N. Dual function of pneumolysin in the early pathogenesis of murine pneumococcal pneumonia. J. Clin. Investig. 1995, 95, 142–150. [Google Scholar] [CrossRef]
- Suárez-Alvarez, B.; García-Suárez, M.d.M.; Méndez, F.J.; de los Toyos, J.R. Characterisation of mouse monoclonal antibodies for pneumolysin: Fine epitope mapping and V gene usage. Immunol. Lett. 2003, 88, 227–239. [Google Scholar] [CrossRef]
- Lee, J.; Suh, E.; Byambabaatar, S.; Lee, S.; Kim, H.; Jin, K.S.; Ree, M. Structural characteristics of pneumolysin and its domains in a biomimetic solution. ACS Omega 2018, 3, 9453–9461. [Google Scholar] [CrossRef] [PubMed]
- Jakubczyk, K.; Kochman, J.; Kwiatkowska, A.; Kałduńska, J.; Dec, K.; Kawczuga, D.; Janda, K. Antioxidant properties and nutritional composition of Matcha green tea. Foods 2020, 9, 483. [Google Scholar] [CrossRef]
- Domon, H.; Nagai, K.; Maekawa, T.; Oda, M.; Yonezawa, D.; Takeda, W.; Hiyoshi, T.; Tamura, H.; Yamaguchi, M.; Kawabata, S.; et al. Neutrophil elastase subverts the immune response by cleaving toll-like receptors and cytokines in pneumococcal pneumonia. Front. Immunol. 2018, 9, 732. [Google Scholar] [CrossRef] [Green Version]
- Cockeran, R.; Theron, A.J.; Steel, H.C.; Matlola, N.M.; Mitchell, T.J.; Feldman, C.; Anderson, R. Proinflammatory interactions of pneumolysin with human neutrophils. J. Infect. Dis. 2001, 183, 604–611. [Google Scholar] [CrossRef] [Green Version]
- Yanagihara, K.; Fukuda, Y.; Seki, M.; Izumikawa, K.; Miyazaki, Y.; Hirakata, Y.; Tsukamoto, K.; Yamada, Y.; Kamhira, S.; Kohno, S. Effects of specific neutrophil elastase inhibitor, sivelestat sodium hydrate, in murine model of severe pneumococcal pneumonia. Exp. Lung Res. 2007, 33, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Xiaokaiti, Y.; Wu, H.; Chen, Y.; Yang, H.; Duan, J.; Li, X.; Pan, Y.; Tie, L.; Zhang, L.; Li, X. EGCG reverses human neutrophil elastase-induced migration in A549 cells by directly binding to HNE and by regulating alpha1-AT. Sci. Rep. 2015, 5, 11494. [Google Scholar] [CrossRef] [PubMed]
- Donà, M.; Dell’Aica, I.; Calabrese, F.; Benelli, R.; Morini, M.; Albini, A.; Garbisa, S. Neutrophil restraint by green tea: Inhibition of inflammation, associated angiogenesis, and pulmonary fibrosis. J. Immunol. 2003, 170, 4335–4341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, T.J.; Mendez, F.; Paton, J.C.; Andrew, P.W.; Boulnois, G.J. Comparison of pneumolysin genes and proteins from Streptococcus pneumoniae types 1 and 2. Nucleic Acids Res. 1990, 18, 4010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domon, H.; Hiyoshi, T.; Maekawa, T.; Yonezawa, D.; Tamura, H.; Kawabata, S.; Yanagihara, K.; Kimura, O.; Kunitomo, E.; Terao, Y. Antibacterial activity of hinokitiol against both antibiotic-resistant and -susceptible pathogenic bacteria that predominate in the oral cavity and upper airways. Microbiol. Immunol. 2019, 63, 213–222. [Google Scholar] [CrossRef]
- Cheung, A.L.; Chien, Y.T.; Bayer, A.S. Hyperproduction of alpha-hemolysin in a sigB mutant is associated with elevated SarA expression in Staphylococcus aureus. Infect. Immun. 1999, 67, 1331–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiyoshi, T.; Domon, H.; Maekawa, T.; Nagai, K.; Tamura, H.; Takahashi, N.; Yonezawa, D.; Miyoshi, T.; Yoshida, A.; Tabeta, K.; et al. Aggregatibacter actinomycetemcomitans induces detachment and death of human gingival epithelial cells and fibroblasts via elastase release following leukotoxin-dependent neutrophil lysis. Microbiol. Immunol. 2019, 63, 100–110. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Unit | Matcha Green Tea Powder * | Sencha Green Tea Infusion, Dried ** |
|---|---|---|---|
| per 1.5 g | per 0.3 g | ||
| Energy | kcal | 3.0 | 0.6 |
| Protein | g | 0.43 | 0.02 |
| Fat | g | 0.09 | 0.002 |
| Ash | g | 0.10 | 0.03 |
| Carbohydrates | g | 0.66 | 0.13 |
| Dietary fiber | g | 0.47 | 0.01 |
| Caffeine | mg | 43.5 | 14.7 |
| Vitamin C | mg | 2.85 | 0.63 |
| Vitamin K | mg | 0.046 | 0.0001 |
| Carotenoids | mg | 2.24 | 0.01 |
| Lutein | mg | 1.16 | 0.003 |
| Theanine | mg | 26.4 | 4.6 |
| Chlorophylls | mg | 14.3 | 0.006 |
| Catechin (C) | mg | 0.6 | 1.7 |
| Epicatechin (EC) | mg | 7.1 | 6.3 |
| Gallocatechin (GC) | mg | 1.7 | 9.3 |
| Epigallocatechin (EGC) | mg | 30.0 | 26.7 |
| Epigallocatechin gallate (EGCG) | mg | 88.5 | 20.4 |
| Gallocatechin gallate (GCG) | mg | 1.0 | 4.5 |
| Epicatehin gallate (ECG) | mg | 15.0 | 3.3 |
| Catechin gallate (CG) | mg | 0.1 | 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sasagawa, K.; Domon, H.; Sakagami, R.; Hirayama, S.; Maekawa, T.; Isono, T.; Hiyoshi, T.; Tamura, H.; Takizawa, F.; Fukushima, Y.; et al. Matcha Green Tea Exhibits Bactericidal Activity against Streptococcus pneumoniae and Inhibits Functional Pneumolysin. Antibiotics 2021, 10, 1550. https://doi.org/10.3390/antibiotics10121550
Sasagawa K, Domon H, Sakagami R, Hirayama S, Maekawa T, Isono T, Hiyoshi T, Tamura H, Takizawa F, Fukushima Y, et al. Matcha Green Tea Exhibits Bactericidal Activity against Streptococcus pneumoniae and Inhibits Functional Pneumolysin. Antibiotics. 2021; 10(12):1550. https://doi.org/10.3390/antibiotics10121550
Chicago/Turabian StyleSasagawa, Karin, Hisanori Domon, Rina Sakagami, Satoru Hirayama, Tomoki Maekawa, Toshihito Isono, Takumi Hiyoshi, Hikaru Tamura, Fumio Takizawa, Yoichi Fukushima, and et al. 2021. "Matcha Green Tea Exhibits Bactericidal Activity against Streptococcus pneumoniae and Inhibits Functional Pneumolysin" Antibiotics 10, no. 12: 1550. https://doi.org/10.3390/antibiotics10121550