
Conjugation Dynamics of Self-Transmissible and Mobilisable Plasmids into E. coli O157:H7 on Arabidopsis thaliana Rosettes

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Transconjugant Frequencies after Filter Mating
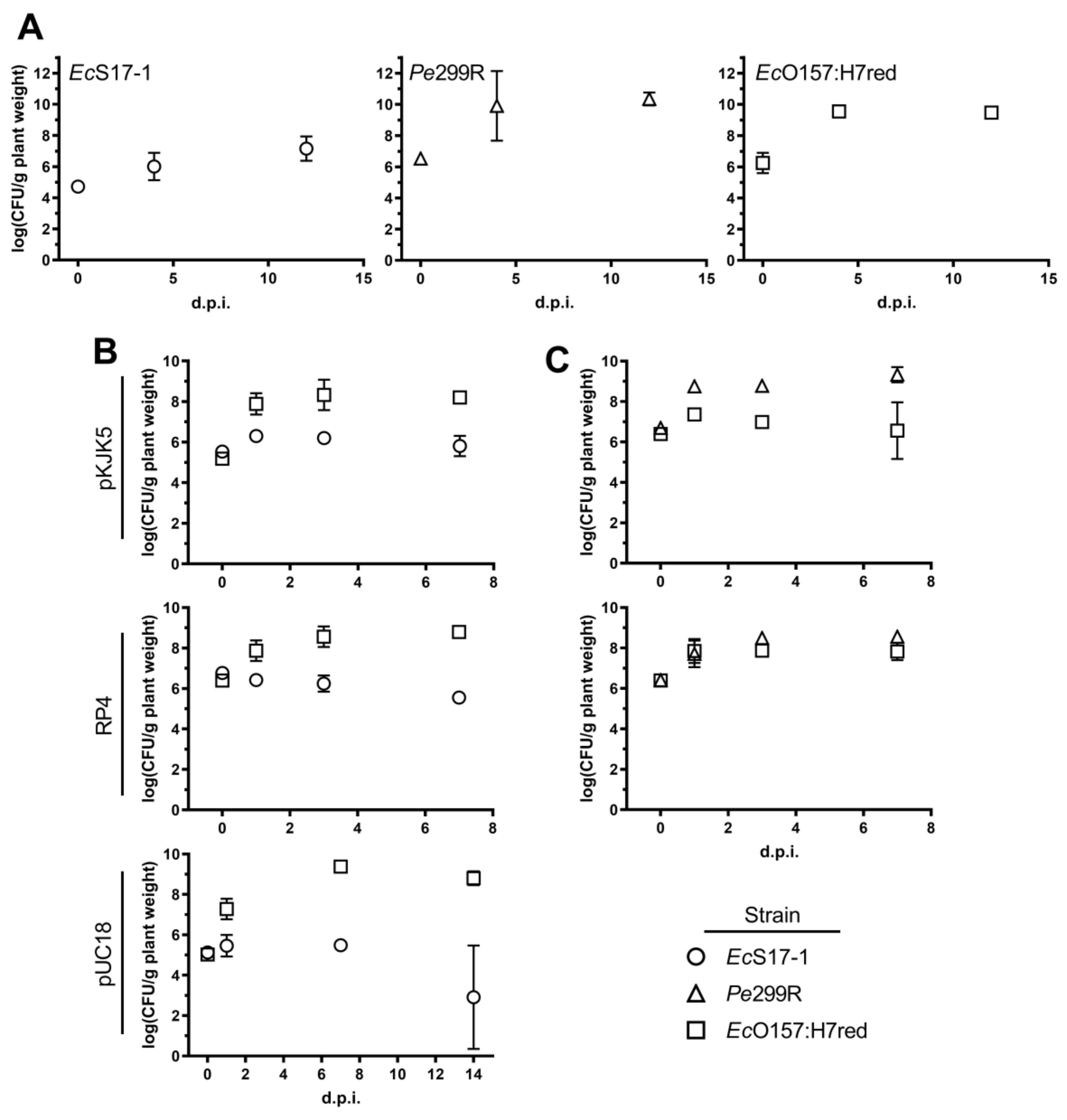
2.2. Growth Dynamics of Individual or Co-Inoculated Bacterial Strains in Planta
2.3. Conjugation Dynamics of Self-Transmissible Plasmids in Planta
2.4. Effect of Non-Self-Transmissible but Mobilisable Plasmids on Transconjugant Frequencies in Planta
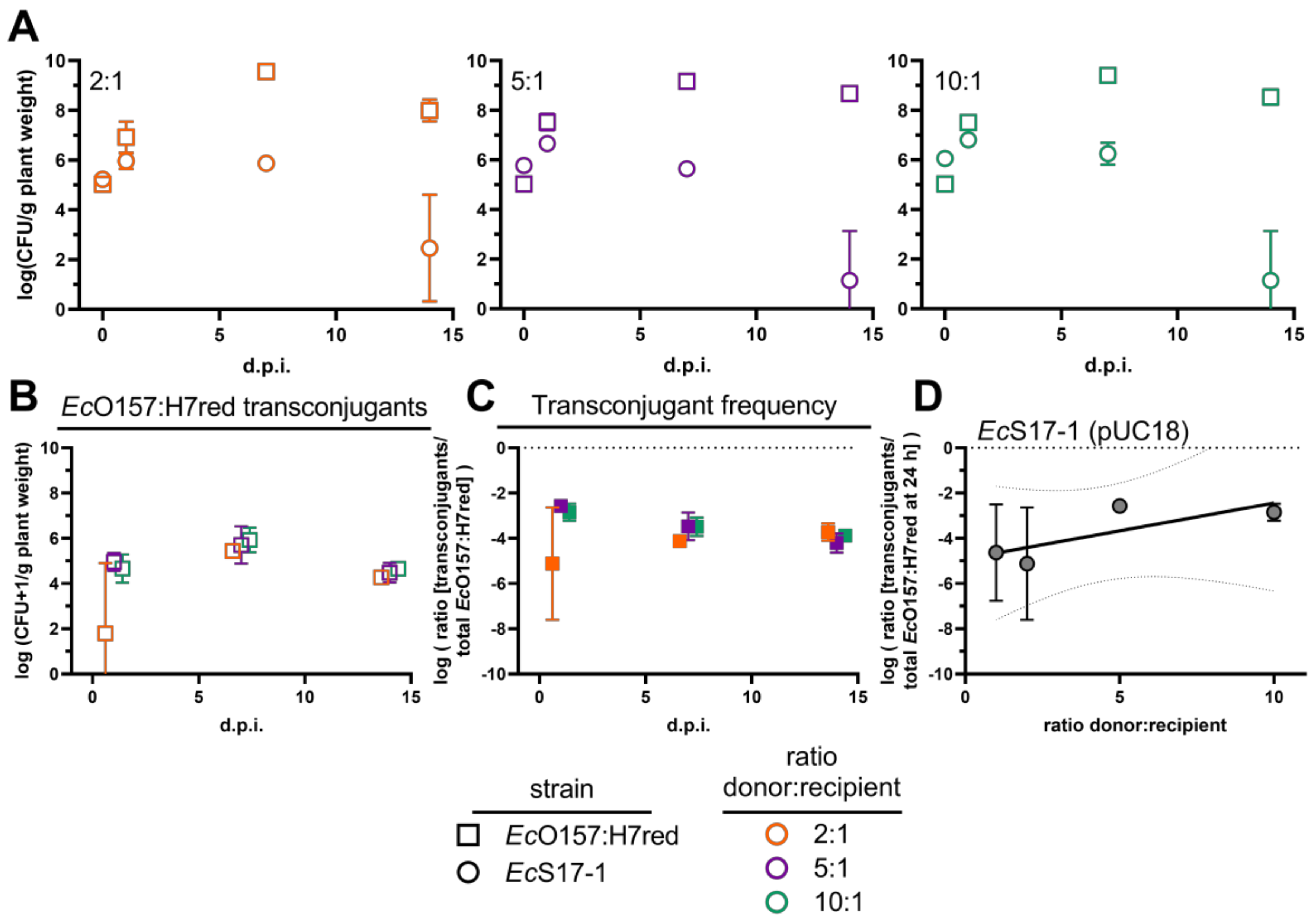
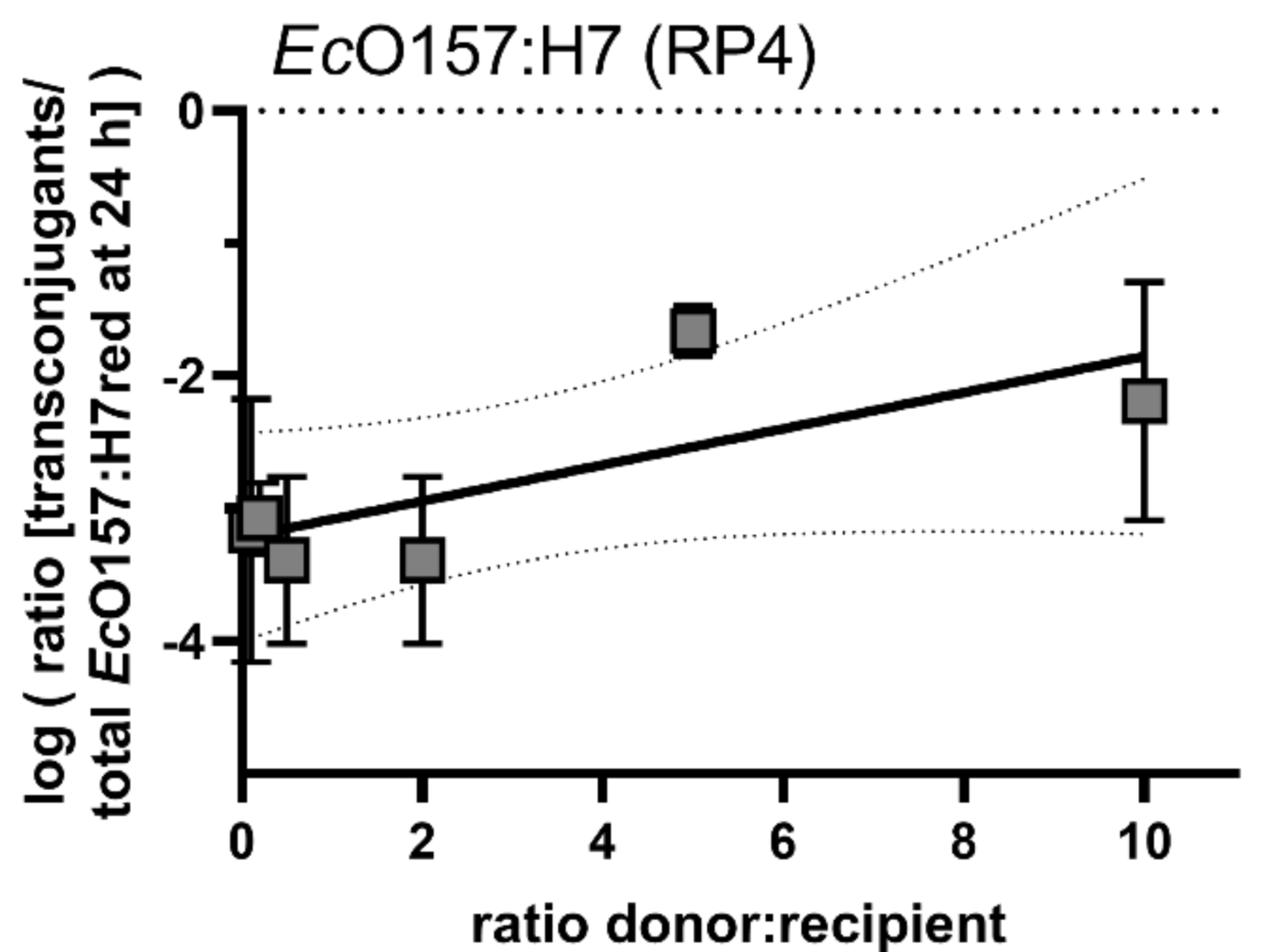
2.5. Effect of Ratio Differences between Donor Cells to Recipients on Transconjugant Frequencies in Planta
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Plasmids Used in the Study
4.3. Conjugation Experiments
4.3.1. Conjugation on Nitrocellulose Filters
4.3.2. Plant Growth
4.3.3. Plant Inoculation with Conjugation Partners and Harvest
4.3.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kardos, N.; Demain, A.L. Penicillin: The medicine with the greatest impact on therapeutic outcomes. Appl. Microbiol. Biotechnol. 2011, 92, 677–687. [Google Scholar] [CrossRef]
- Aminov, R.I. A brief history of the antibiotic era: Lessons learned and challenges for the future. Front. Microbiol. 2010, 1, 134. [Google Scholar] [CrossRef] [Green Version]
- Neu, H.C. The crisis in antibiotic resistance. Science 1992, 257, 1064–1073. [Google Scholar] [CrossRef] [Green Version]
- Kumarasamy, K.K.; Toleman, M.A.; Walsh, T.R.; Bagaria, J.; Butt, F.; Balakrishnan, R.; Chaudhary, U.; Doumith, M.; Giske, C.G.; Irfan, S.; et al. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: A molecular, biological, and epidemiological study. Lancet Infect. Dis. 2010, 10, 597–602. [Google Scholar] [CrossRef]
- D’Costa, V.M.; King, C.E.; Kalan, L.; Morar, M.; Sung, W.W.L.; Schwarz, C.; Froese, D.; Zazula, G.; Calmels, F.; Debruyne, R.; et al. Antibiotic resistance is ancient. Nature 2011, 477, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Martínez, J.L.; Coque, T.M.; Baquero, F. What is a resistance gene? Ranking risk in resistomes. Nat. Rev. Microbiol. 2015, 13, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Palumbi, S.R. Humans as the World’s Greatest Evolutionary Force. Science 2001, 293, 1786–1790. [Google Scholar] [CrossRef] [PubMed]
- Cantas, L.; Shah, S.Q.A.; Cavaco, L.M.; Manaia, C.; Walsh, F.; Popowska, M.; Garelick, H.; Bürgmann, H.; Sørum, H. A brief multi-disciplinary review on antimicrobial resistance in medicine and its linkage to the global environmental microbiota. Front. Microbiol. 2013, 4, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, T.F.; Pla, M.D.P.; Mayer, K.H.; Kishi, H.; Gilleece, E.; Syvanen, M.; Hopkins, J.D. Intercontinental spread of a new antibiotic resistance gene on an epidemic plasmid. Science 1985, 230, 87–88. [Google Scholar] [CrossRef]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [Green Version]
- Allen, H.K.; Donato, J.; Wang, H.H.; Cloud-Hansen, K.A.; Davies, J.; Handelsman, J. Call of the wild: Antibiotic resistance genes in natural environments. Nat. Rev. Microbiol. 2010, 8, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.M.; Nielsen, K.M. Mechanisms of, and barriers to, horizontal gene transfer between bacteria. Nat. Rev. Microbiol. 2005, 3, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Burrus, V.; Pavlovic, G.; Decaris, B.; Guédon, G. Conjugative transposons: The tip of the iceberg. Mol. Microbiol. 2002, 46, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Baquero, F.; Tedim, A.P.; Coque, T.M. Antibiotic resistance shaping multi-level population biology of bacteria. Front. Microbiol. 2013, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Klümper, U.; Riber, L.; Dechesne, A.; Sannazzarro, A.; Hansen, L.H.; Sørensen, S.J.; Smets, B.F. Broad host range plasmids can invade an unexpectedly diverse fraction of a soil bacterial community. ISME J. 2015, 9, 934–945. [Google Scholar] [CrossRef] [Green Version]
- Colombi, E.; Straub, C.; Künzel, S.; Templeton, M.D.; McCann, H.C.; Rainey, P.B. Evolution of copper resistance in the kiwifruit pathogen Pseudomonas syringae pv. actinidiae through acquisition of integrative conjugative elements and plasmids. Environ. Microbiol. 2017, 19, 819–832. [Google Scholar] [PubMed]
- Powell, B.J.; Purdy, K.J.; Thompson, I.P.; Bailey, M.J. Demonstration of tra plasmid activity in bacteria indigenous to the phyllosphere of sugar beet; gene transfer to a recombinant pseudomonad. FEMS Microbiol. Ecol. 1993, 12, 195–206. [Google Scholar] [CrossRef]
- Normander, B.; Christensen, B.B.; Molin, S.; Kroer, N. Effect of bacterial distribution and activity on conjugal gene transfer on the phylloplane of the bush bean (Phaseolus vulgaris). Appl. Environ. Microbiol. 1998, 64, 1902–1909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björklöf, K.; Nurmiaho-Lassila, E.L.; Klinger, N.; Haahtela, K.; Romantschuk, M. Colonization strategies and conjugal gene transfer of inoculated Pseudomonas syringae on the leaf surface. J. Appl. Microbiol. 2000, 89, 423–432. [Google Scholar] [CrossRef]
- van Elsas, J.D.; Turner, S.; Bailey, M.J. Horizontal gene transfer in the phytosphere. New Phytol. 2003, 157, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Blau, K.; Bettermann, A.; Jechalke, S.; Fornefeld, E.; Vanrobaeys, Y.; Stalder, T.; Top, E.M.; Smalla, K. The transferable resistome of produce. bioRxiv 2018, 350629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruinen, J. The phyllosphere. Plant Soil 1961, 15, 81–109. [Google Scholar] [CrossRef]
- Landers, T.F.; Cohen, B.; Wittum, T.E.; Larson, E.L. A review of antibiotic use in food animals: Perspective, policy, and potential. Public Health Rep. 2012, 127, 4–22. [Google Scholar] [CrossRef] [Green Version]
- Heuer, H.; Schmitt, H.; Smalla, K. Antibiotic resistance gene spread due to manure application on agricultural fields. Curr. Opin. Microbiol. 2011, 14, 236–243. [Google Scholar] [CrossRef]
- Wolters, B.; Kyselková, M.; Krögerrecklenfort, E.; Kreuzig, R.; Smalla, K. Transferable antibiotic resistance plasmids from biogas plant digestates often belong to the IncP-1ε subgroup. Front. Microbiol. 2014, 5, 765. [Google Scholar]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef]
- Rastogi, G.; Coaker, G.L.; Leveau, J.H.J. New insights into the structure and function of phyllosphere microbiota through high-throughput molecular approaches. FEMS Microbiol. Lett. 2013, 348, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandl, M.T. Fitness of human enteric pathogens on plants and implications for food safety. Annu. Rev. Phytopathol. 2006, 44, 367–392. [Google Scholar] [CrossRef] [Green Version]
- Heaton, J.C.; Jones, K. Microbial contamination of fruit and vegetables and the behaviour of enteropathogens in the phyllosphere: A review. J. Appl. Microbiol. 2008, 104, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Innerebner, G.; Knief, C.; Vorholt, J.A. Protection of Arabidopsis thaliana against leaf-pathogenic Pseudomonas syringae by Sphingomonas strains in a controlled model system. Appl. Environ. Microbiol. 2011, 77, 3202–3210. [Google Scholar] [CrossRef] [Green Version]
- Remus-Emsermann, M.N.P.; Lücker, S.; Müller, D.B.; Potthoff, E.; Daims, H.; Vorholt, J.A. Spatial distribution analyses of natural phyllosphere-colonizing bacteria on Arabidopsis thaliana revealed by fluorescence in situ hybridization. Environ. Microbiol. 2014, 16, 2329–2340. [Google Scholar] [CrossRef]
- Heinemann, J.A. Genetics of gene transfer between species. Trends Genet. 1991, 7, 181–185. [Google Scholar] [CrossRef]
- Simon, R.; Priefer, U.; Pühler, A. A broad host range mobilization system for in vivo genetic engineering: Transposon mutagenesis in gram negative bacteria. Biotechnology 1983, 1, 784–791. [Google Scholar] [CrossRef]
- Wilson, M.; Lindow, S.E. Ecological Similarity and Coexistence of Epiphytic Ice-Nucleating (Ice+) Pseudomonas syringae Strains and a Non-Ice-Nucleating (Ice−) Biological Control Agent. Appl. Environ. Microbiol. 1994, 60, 3128–3137. [Google Scholar] [CrossRef] [Green Version]
- Smits, T.H.M.; Rezzonico, F.; Kamber, T.; Blom, J.; Goesmann, A.; Ishimaru, C.A.; Frey, J.E.; Stockwell, V.O.; Duffy, B. Metabolic versatility and antibacterial metabolite biosynthesis are distinguishing genomic features of the fire blight antagonist Pantoea vagans C9-1. PLoS ONE 2011, 6, e22247. [Google Scholar] [CrossRef] [Green Version]
- Remus-Emsermann, M.N.P.; Kim, E.B.; Marco, M.L.; Tecon, R.; Leveau, J.H.J. Draft Genome Sequence of the Phyllosphere Model Bacterium Pantoea agglomerans 299R. Genome Announc. 2013, 1, e00036-13. [Google Scholar] [CrossRef] [Green Version]
- Lilley, A.K.; Bailey, M.J.; Barr, M.; Kilshaw, K.; Timms-Wilson, T.M.; Day, M.J.; Norris, S.J.; Jones, T.H.; Godfray, H.C.J. Population dynamics and gene transfer in genetically modified bacteria in a model microcosm: Gene transfer in phytosphere bacteria. Mol. Ecol. 2003, 12, 3097–3107. [Google Scholar] [CrossRef]
- Remus-Emsermann, M.N.P.; Schlechter, R.O. Phyllosphere microbiology: At the interface between microbial individuals and the plant host. New Phytol. 2018, 218, 1327–1333. [Google Scholar] [CrossRef] [PubMed]
- Remus-Emsermann, M.N.P.; Tecon, R.; Kowalchuk, G.A.; Leveau, J.H.J. Variation in local carrying capacity and the individual fate of bacterial colonizers in the phyllosphere. ISME J. 2012, 6, 756–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tecon, R.; Leveau, J.H.J. The mechanics of bacterial cluster formation on plant leaf surfaces as revealed by bioreporter technology. Environ. Microbiol. 2012, 14, 1325–1332. [Google Scholar] [CrossRef] [PubMed]
- Monier, J.-M.; Lindow, S.E. Spatial organization of dual-species bacterial aggregates on leaf surfaces. Appl. Environ. Microbiol. 2005, 71, 5484–5493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopatkin, A.J.; Huang, S.; Smith, R.P.; Srimani, J.K.; Sysoeva, T.A.; Bewick, S.; Karig, D.K.; You, L. Antibiotics as a selective driver for conjugation dynamics. Nat. Microbiol. 2016, 1, 16044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, K.-H.; Schweizer, H.P. mini-Tn7 insertion in bacteria with single attTn7 sites: Example Pseudomonas aeruginosa. Nat. Protoc. 2006, 1, 153–161. [Google Scholar] [CrossRef]
- Barth, P.T.; Grinter, N.J. Map of plasmid RP4 derived by insertion of transposon C. J. Mol. Biol. 1977, 113, 455–474. [Google Scholar] [CrossRef]
- Sengeløv, G.; Kristensen, K.J.; Sørensen, A.H.; Kroer, N.; Sørensen, S.J. Effect of genomic location on horizontal transfer of a recombinant gene cassette between Pseudomonas strains in the rhizosphere and spermosphere of barley seedlings. Curr. Microbiol. 2001, 42, 160–167. [Google Scholar] [CrossRef]
- Remus-Emsermann, M.N.P.; Gisler, P.; Drissner, D. MiniTn7-transposon delivery vectors for inducible or constitutive fluorescent protein expression in Enterobacteriaceae. FEMS Microbiol. Lett. 2016, 363, fnw178. [Google Scholar] [CrossRef] [PubMed]
- Vogel, C.; Innerebner, G.; Zingg, J.; Guder, J.; Vorholt, J.A. Forward genetic in planta screen for identification of plant-protective traits of Sphingomonas sp. strain Fr1 against Pseudomonas syringae DC3000. Appl. Environ. Microbiol. 2012, 78, 5529–5535. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain (Notes of Important Properties) | Abbreviation | Antibiotic Resistance | Reference |
| E. coli O157:H7::MRE103 Δstx (grows on lactose, red fluorescent, does not produce Shiga toxin) | EcO157:H7red | Rifampicin | [46] |
| E. coli O157:H7 Δstx (grows on lactose, does not produce Shiga toxin) | EcO157:H7 | Not applicable | NCTC 12900 |
| E. coli S17-1 λpir (auxotroph) | EcS17-1 | Streptomycin | [33] |
| Pantoea eucalypti 299R (grows slowly on lactose) | Pe299R | Rifampicin | [36] |
| Plasmid (Notes of Important Properties; Size) | Abbreviation | Antibiotic Resistance | Reference |
| pUC18T-mini-Tn7T-Gm-eYFP (mobilisable, confers yellow fluorescence; 5.9 Kbp) | pUC18 | Gentamicin | [43] |
| pKJK5::Plac::gfp (self-transmissible, confers green fluorescence; 54 Kbp) | pKJK5 | Kanamycin | [15] |
| RP4::Plac::gfp (self-transmissible, confers green fluorescence; 56 Kbp) | RP4 | Kanamycin | [15] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Remus-Emsermann, M.N.P.; Aicher, D.; Pelludat, C.; Gisler, P.; Drissner, D. Conjugation Dynamics of Self-Transmissible and Mobilisable Plasmids into E. coli O157:H7 on Arabidopsis thaliana Rosettes. Antibiotics 2021, 10, 928. https://doi.org/10.3390/antibiotics10080928
Remus-Emsermann MNP, Aicher D, Pelludat C, Gisler P, Drissner D. Conjugation Dynamics of Self-Transmissible and Mobilisable Plasmids into E. coli O157:H7 on Arabidopsis thaliana Rosettes. Antibiotics. 2021; 10(8):928. https://doi.org/10.3390/antibiotics10080928
Chicago/Turabian StyleRemus-Emsermann, Mitja N. P., David Aicher, Cosima Pelludat, Pascal Gisler, and David Drissner. 2021. "Conjugation Dynamics of Self-Transmissible and Mobilisable Plasmids into E. coli O157:H7 on Arabidopsis thaliana Rosettes" Antibiotics 10, no. 8: 928. https://doi.org/10.3390/antibiotics10080928