
Bee Pollen and Bee Bread as a Source of Bacteria Producing Antimicrobials


1
Department of Pharmaceutical Technology and Biochemistry, Faculty of Chemistry, Gdańsk University of Technology, ul. G. Narutowicza 11/12, 80-233 Gdańsk, Poland
2
Department of Food Science, Cornell University, Ithaca, NY 14853, USA
*
Author to whom correspondence should be addressed.
Antibiotics 2021, 10(6), 713; https://doi.org/10.3390/antibiotics10060713
Submission received: 21 April 2021
/
Revised: 5 June 2021
/
Accepted: 9 June 2021
/
Published: 13 June 2021
(This article belongs to the Special Issue Honey Bee Products as an Alternative or Complement to Classical Antibiotics – 2nd Edition)
Abstract
:The principal objective of the study was the isolation and identification of bacteria that are present in mature bee bread (BB) and dried (ready for selling and consumption) bee pollen (BP). Obtained isolates were screened for their potential to inhibit select human pathogenic bacteria and their ability to produce enzymes of particular industrial importance. Four and five samples of BP and BB, respectively, were used for the study. In total, 81 strains of bacteria were isolated, and 34 (42%) of them exhibited antagonistic interactions with at least one reference strain of pathogenic bacteria, namely Staphylococcus aureus ATCC 25923, Staphylococcus aureus ATCC 29213, Staphylococcus epidermidis 12228, Pseudomonas aeruginosa ATCC 27857, and Escherichia coli ATCC 25922. The sequencing of the 16S rRNA gene revealed that all strains producing antimicrobials belong to the genus Bacillus spp., and among them, five species were identified: B. pumilus (n = 17), B. altitudinis (n = 9), B. licheniformis (n = 4), B. subtilis (n = 2), and B. safensis (n = 1). Furthermore, 69, 54, 39, and 29 of the strains exhibited lipolytic, proteolytic, cellulolytic, and esterolytic activity, respectively. Alpha amylase and beta galactosidase activity were rarely observed, and none of the strains produced laccase. The outcomes of the study revealed that BP and BB can be considered potential sources of bacteria producing antimicrobial agents and/or enzymes of particular industrial importance. Of course, additional research is required to verify this hypothesis, but the results of preliminary studies are promising.
1. Introduction
Since ancient times, honey bee products, particularly honey and propolis, have been used as traditional remedies. Both these product exhibit high antimicrobial activity and have mostly been applied for treatment of infected and difficult to heal wounds. As in the case of many other natural products, the investigation of their antibacterial/antifungal potential was impeded by the discovery of antibiotics. However, renewed interest in the investigation and use of the pharmacological (not only antimicrobial) potential of bee products has been observed within the last couple of decades. Recent studies performed with modern analytical techniques and using in vitro and in vivo models have proved that the chemical components of bee products exhibit a broad range of health-beneficial properties including antimicrobial, antioxidant, anti-inflammatory, anti-cancer, and immunomodulatory activity [1,2,3,4,5]. It has been found that the enzymatic production (by glucose oxidase (GOx)) of hydrogen peroxide is a dominant mechanism by which honey collected by bees from most plant sources exert bacteriostatic and bactericidal activity. The physiochemical properties of these products, namely high osmotic pressure and low pH, as well as some of their components, e.g., polyphenols and bee defensin−1, only support the antimicrobial effect of H2O2 [1,6,7]. Thus, these honeys are called peroxide honeys. However, interesting results presented by Brudzynski and coworkers [7,8,9,10], Bucekova et al., [11] and Grecka and colleagues [12] suggested an important role of plant-derived phytochemicals (mostly polyphenols) for the level of production of hydrogen peroxide in some honey types and probably also the transformation of H2O2 for most active radical products, e.g., OH˙ [7,8,9,10,11,12]. A unique mechanism of antimicrobial activity has been identified for New Zealand’s manuka honey and several Australian and Malaysian honeys. High efficiency in the inhibition of bacterial growth by these product is attributed to a non-peroxide component—methylglyoxal [13,14,15].
Propolis is a highly agglutinative, resinous substance of complex chemical composition that is collected by bees from flower and leaf buds. Propolis-containing extracts exhibit a broad spectrum of biological activities, among which antimicrobial potential has been the most intensively investigated [3,4,16]. The research carried out in our research group revealed the high antibacterial—particularly anti-staphylococcal [17] and antifungal [18]—activity of propolis collected in Polish apiaries, and flavonoids (flavonols, flavones, and flavanones) have been identified as components crucial for the antimicrobial activity of these products. Many trials have confirmed usefulness of propolis-containing products (e.g., extracts, ointments, wound materials, and dental materials such as toothpaste, glass-ionomer cement (GIC), and dental varnish) for treatment and prophylaxis against bacterial and fungal infections [4,19]. Interesting health-associated properties including antimicrobial potential have also been identified and described for lesser known and less popular bee products, namely royal jelly [20,21], bee wax [22], and bee venom [23].
The bee products that have recently gained particular popularity are bee pollen (BP) and bee bread (BB) [5]. Because of their high nutrition values, both of them are classified as functional foods [24] and both exhibit a wide range of therapeutic properties, such as antimicrobial, antioxidant, anti-radiation, anti-inflammatory, anti-tumor, hepatoprotective, and chemopreventive/chemoprotective benefits [5,24,25,26,27]. The term “bee bread” refers to the collected pollen that is processed by bees and fermented [5]. The exact mechanism of the biotransformation of BP to BB is still not fully elucidated. However, it known that enzymes from bees’ glands (e.g., amylases that are responsible for starch hydrolysis), as well as bacteria (mostly lactic acid bacteria—LAB) and some yeasts sourced from bees’ saliva and surfaces of pollen loads, play crucial roles in BP fermentation and BB production [5,28,29,30]. Some of the BB is stored in the wells of the honeycomb through the winter, and in the spring it is used as a main source of proteins for the new populations of bee larvae. It has been also found that ethanolic or methanolic extracts of components of both BB and BP exhibit antimicrobial potential. The outcomes of our recent study revealed a considerably higher antimicrobial potential of extracts produced from BB compared to BP extracts [31]. We also found the efficient inhibition of growth of Staphylococcus aureus in water suspensions of both products [31].
Important gaps in our knowledge remain regarding the microbial ecosystem of bee products, including both bacteria and fungi. Still very little is known about species composition and the role of these microorganisms in maturing bee products (e.g., the biotransformation of BP to BB) and in the protection of honey and bee bread against microbial spoilage, which is crucial for the health of bees (both mature and larvae) and humans who consume these products. There is mounting evidence implicating microbial ecosystem of the bee raw materials (nectar and pollen)–bee products (honey and bee bread)–honey bee axis involved in the production of a range of antimicrobial agents. These agents are used as weaponry in competitive interspecies interactions to effectively kill competing microorganisms in the fight for nutrients and space in each of these niches (nectar, pollen, honey, bee bread, and honey bee). Among the secondary metabolites produced by microorganisms that constitute the microflora of bee products are antimicrobial peptides, bacteriocins, surfactants, siderophores, proteolytic enzymes, and cell wall-degrading enzymes [32]. The main goal of this study was to investigate the ability of bacteria that constitute the microbiome of BP and BB for the growth inhibition of selected pathogenic microorganisms. Most of isolated strains of bacteria were identified as Bacillus spp., and some of them exhibited high antagonistic activity against important clinical human pathogens including staphylococci, E. coli, and P. aeruginosa. Our future studies will be focused on the identification of the molecular mechanism or metabolites that are responsible for these antagonistic interactions. We believe this could lead to the identification of producers of new antimicrobial agents. Moreover, most of isolates derived from both raw materials revealed high proteolytic, lipolytic, esterolytic, and cellulolytic activity. The outcomes of the study revealed that bacteria isolated from BP and BB can be considered a possible source of novel antimicrobial compounds and enzymes of particular industrial importance.
2. Results
As is shown in Table 1, the investigated samples of BP and BB presented different, though generally low, levels of microbial contamination—only aerobic and facultative aerobic were considered in this study. Four products (44%), two samples of each BB and BP, exhibited a level of contamination of above 103 CFU (colony forming units) per gram of the raw material. The other five samples contained less bacteria, from 100 to 600 CFU per gram of the raw material. No evident differences in the level of microbial contamination between BP and BB were observed in this study.
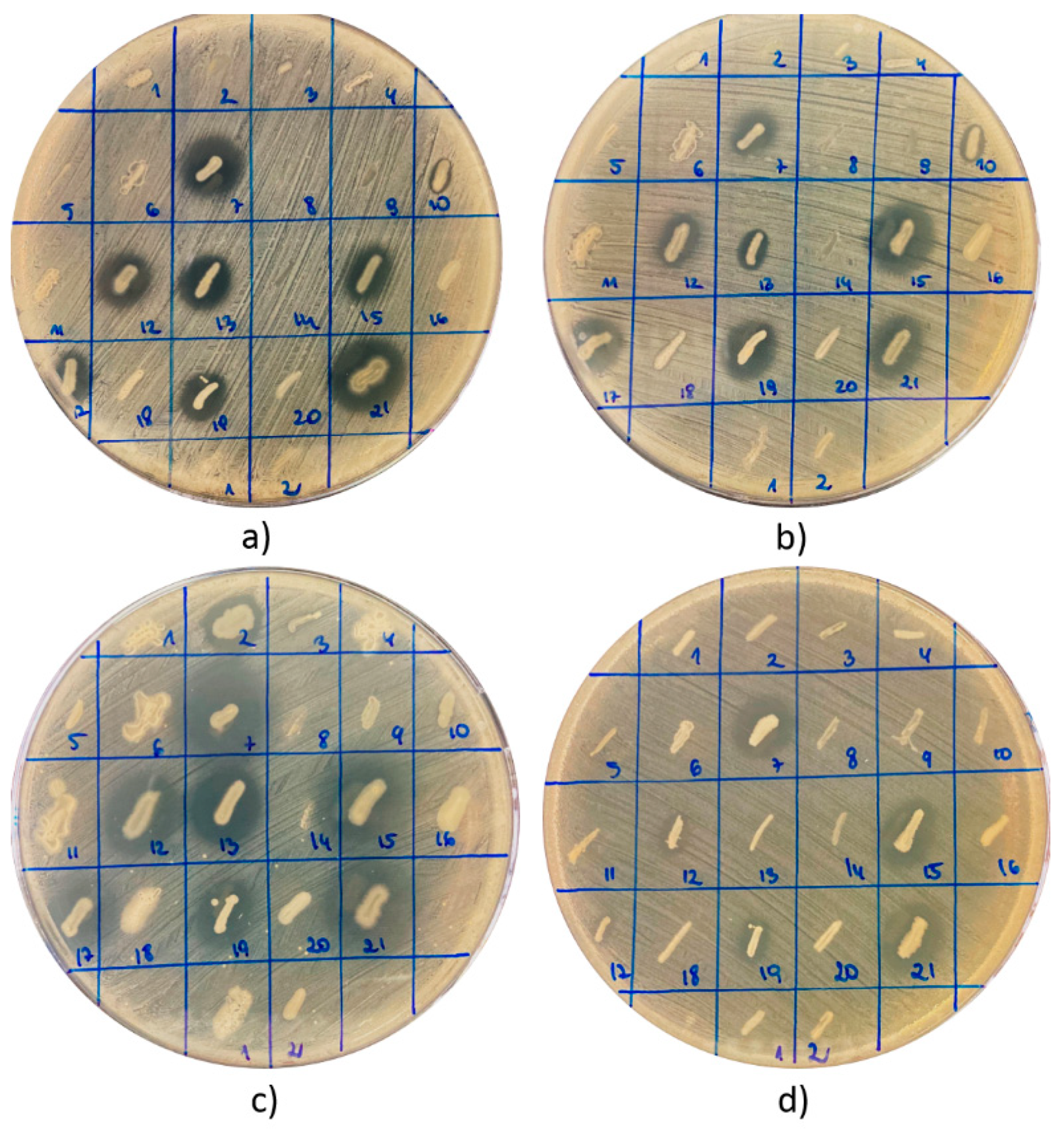
In total, 81 strains of bacteria were recovered from nine tested products (Table 1 and Table 2). In each case, the bacteria were cultivated from a 0.1 mL suspension of the raw material in sterile water (1:10 w/v). All these isolates were screened for antagonistic interactions with pathogenic bacteria and the production of select essential hydrolytic enzymes. The antagonistic relationship was investigated through the observation of growth inhibition zones (GIZs) of indicator strains of bacteria around the growing colonies of tested strains—isolates from BB or BP (Figure 1).
Considering Gram-positive staphylococci, the antagonistic activity was observed for 27 (33.3%), 29 (35.8%), and 22 (27.2%) strains against S. aureus ATCC 25923, S. aureus ATCC 29213, and S. epidermidis ATCC 12228, respectively. A considerable number of strains, n = 32 (39.5%), inhibited the growth of Pseudomonas aeruginosa ATCC 27853, while activity against E. coli ATCC 25922 was not so common and was confirmed for 15 isolates (18.5%). The largest number of active isolates was recovered from bee bread assigned as BB19. Eight strains inhibited the growth of staphylococci, and an antagonistic relationship with E. coli ATCC 25922 and P. aeruginosa ATCC 27853 was confirmed for five (23.8%) and nine (42.9%) strains, respectively. Forty percent or more of isolates derived from products BP3, BP15, BP20, and BB10 exhibited antagonistic potential against both strains of S. aureus and P. aeruginosa ATCC 27853. On the other hand, the only strain recovered from the raw material was assigned as BP12, and both isolates from BB3 did not exhibit any antimicrobial properties. Relatively low percentage levels of active strains were also found in the cases of BB6 and BB15.
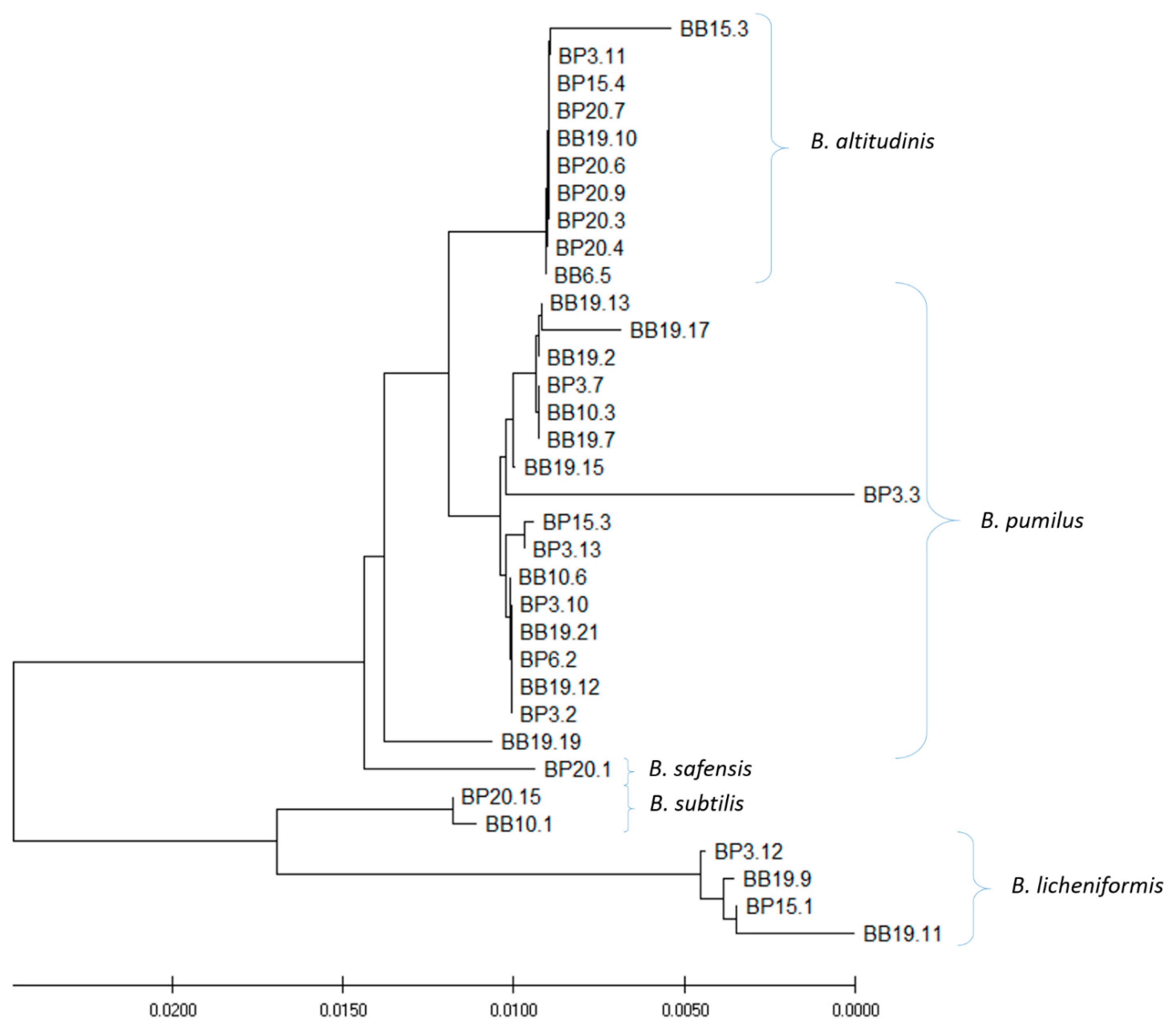
The 34 out of 81 isolated strains that exhibited antagonistic activity against at least one indicator strain were selected for species identification and amplification and sequencing of the gene coding for 16S rRNA (Table 2). The bioinformatics analysis of 16S rRNA gene sequences (carried out with the BLAST software) revealed that all tested strains belong to the genus Bacillus. In general, five species were distinguished among the isolates. Most of the strains (eighteen) were classified as B. pumilus. Nine isolates were identified as a second most common species—B. altitudinis. Four, two, and one strains were recognized as B. licheniformis, subtilis, and safensis, respectively. However, it is necessary to remember that sequences of the 16S rRNA gene of different species of Bacillus spp. are characterized by a high level of similarity or even identity. Thus, further analysis, e.g., whole genome sequencing or mass spectrometry, would be required for final species identification. The phylogenetic analysis based on results of comparative analysis of the sequences of 16S rRNA genes revealed some diversity between the tested strains (Figure 2) and generally confirmed results of the classification of the species (Figure 2).
Five clusters of the strains could be distinguished, which was in agreement with species identification.
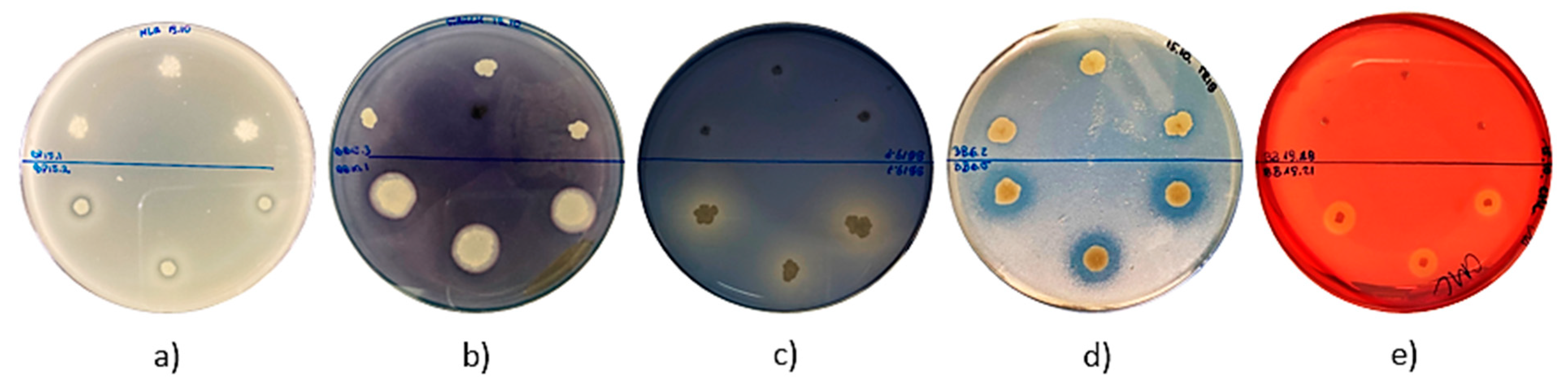
Some interesting observations were also made regarding the assessment of the enzymatic activity of isolated strains of bacteria (Table 3 and Figure 3). None of the strains were able to produce laccase. No discoloration of the bacterial colonies was observed on the agar medium supplemented with guaiacol. Furthermore, a small number of strains (n = 3; 3.7%) exhibited amylolytic activity. Flooding the active strains with Lugol’s solution resulted in the appearance of clear halos around the colonies (Figure 3b). Eleven strains (13.58%) formed lightly blue colonies on the LA agar medium supplemented with X−gal, which probably confirmed β−galactosidase production, though with a low efficiency (data not shown). Almost half of the isolates presented esterolytic (n = 29; 36%) and proteolytic (n = 39; 48%) activity. In the case of strains isolated from BP and BB that are able to produce proteases and esterases, clear halos around the grown bacterial colonies were observed on the media supplemented with skimmed milk and tributyrin, respectively (Figure 3a,d). Moreover, a significant number of strains (n = 54; 67%) displayed cellulolytic activity, and 18 of them were classified as strong producers. In this case, the halos zones appeared around the colonies grown on LA agar medium supplemented with carboxymethylcellulose and flooded with Congo red solution (Figure 3e). Interestingly, the largest number of isolates (n = 69; 85.1%) exhibited lipolytic activity. The isolates contributed to the formation of bright halos around the colonies on Spirit Blue Agar (Figure 3c).
3. Discussion
The recently observed increase in the popularity and consumption level of BP and BB is mainly a consequence of the high nutritional value and health benefit properties of these products, including the high contents of vitamins, minerals, amino acids, some fatty acids, and polyphenols (antioxidants) that seem to be the most important [5,24,25,26]. However, very little is known about the microbiome of both these products. Both bacterial and fungal communities associated with BP and BB are important in at least four different aspects: (1) the process of biotransformation of BP to BB; (2) the stability of BB in the hive (in the cells of honey combs) during long term storage in the winter season; (3) the microbial safety of bees, particularly bee larvae that are fed with the BP/BB; and (4) the influence on the health of people who are consumers of BP/BB. The collected pollen loads by bee workers are prone to microbial deterioration—mostly due to molds. The BP that is sold in markets must be dried, which inhibits the growth of molds and other pathogenic microorganisms. For bees, the BP is, in fact, the only raw material for BB production. The BP loads collected by bee workers are mixed with small amounts of the secretion from the bee’s saliva, tightly packed in honeycomb cells, and finally covered with a thin layer of honey and a wax lid. Subsequently, under these anaerobic conditions, the BP undergoes the biotransformation process to BB. The exact biochemical mechanism of the biotransformation processes remains not fully understood. However, it is known that different enzymes from bees’ glandular secretion, as well as bacteria that are present in bees’ saliva and on the surface of pollen loads, are crucial for this process [5,33,34]. It still remains unclear which species of bacteria participate in BB maturing. However, the outcomes of several investigations have suggested that lactic acid bacteria are of primary importance. Vasquez and Olofsson (2009) observed the intense growth of these bacteria within maturing BB for about two weeks—the first step of BP biotransformation [30]. The LAB are important part of Apis mellifera gut community and are probably introduced to the raw material—BP from bees’ saliva [35]. The presence of LAB seems to be particularly important from the point of view of the microbial stability and preservation of the final product—BB. These bacteria produce lactic acid, bacteriocins, and aliphatic acids (products of lipids hydrolysis) that efficiently inhibit the growth of not only pathogenic (for both bees and humans, the consumers of BB) microorganisms but also bacteria, yeasts, and molds that could cause microbial deterioration or undesirable sensory changes [35]. Iorizzo and coworkers (2020) revealed the high inhibitory activity of LAB, namely Lactobacillus kunkeei and Lactiplantibacillus plantarum (isolated from bees’ gastrointestinal tract and bee products) against the important bee pathogens Ascosphaera apis and Paenibacillus larvae, respectively [36,37]. It also has been found that yeasts and molds participate in BP biotransformation. Detry et al. (2020) identified Starmerella, Metschnikowia, and Zygosaccharomyces as the most common yeast species in bee bread. However, the high abundance of yeasts in fresh bee bread decreased rapidly with the storage duration. Starmerella species dominated fresh bee bread, while mostly Zygosaccharomyces members were isolated from aged bee bread [38]. Disayathanoowat et al. (2020) investigated dynamic of bacterial and fungal community structures in corbicula pollen and hive−stored BB collected in China. They found that corbicula pollen was colonized by the Enterobacteriaceae bacterium (Escherichia−Shigella, Panteoa, and Pseudomonas) group; however, the number of bacteria significantly decreased in hive−stored bee bread in less than 72 h. In contrast, Acinetobacter was highly abundant and could utilize protein sources. In terms of the fungal community, the genus Cladosporium remained abundant in both corbicula pollen and hive−stored bee bread. The authors also concluded that filamentous fungus might encourage honey bees to reserve pollen by releasing organic acids [29].
Both mature BB and dried BP—ready for sale in markets—are considered microbial−safe and free from dangerous pathogenic microorganisms. However, none of these products are sterile, and very little is known about the microbiota of these products, including species composition and the metabolic and enzymatic properties of bacterial and fungal communities present in BP and BB. The herein presented results confirmed the generally low level of microbial contamination of samples of both products, with a maximum level of contamination of approximately 2.1 × 103 CFU/g. Interestingly, all of 34 isolates that exhibited antimicrobial activity were classified into the genus Bacillus spp., and five different species were identified: B. subtilis, B. licheniformis, B. pumilus, B. altitudinis, and B. safensis. The above−mentioned studies did not show the presence of Bacillus spp. in BB samples [29,30,38]. In our opinion, this difference could be explained by the fact that only mature BB samples harvested from honeycomb cells and stored for about four months (under refrigeration) and dried BP samples were used in our study. The phytochemicals present in the raw material and in honey added to the BP, as well as the metabolites of bacteria growing in the maturing product, formed an unfavorable environment for the growth and development of most microorganisms. Thus, only highly resistant bacteria, e.g., spore−forming Bacillus spp. bacteria, can survive under these conditions. The outcomes of our previous investigation revealed very similar species composition and properties of microorganisms isolated from honey samples [39]. Most of these isolates were classified as Bacillus spp., and most of them exhibited the ability to produce metabolites of antibacterial activity [39]. One of these strains, namely Paenibacillus alvei MP1, was found to be an efficient producer of proteinaceous agent that exhibited promising activity against a broad spectrum of pathogenic bacteria [40]. Moreover, several genes responsible for antimicrobial activity have been identified in the genome of P. alvei MP1 [41]. Some other research groups have also reported the isolation of antimicrobials producing bacteria from honey. Lee et al. (2008) screened six US honeys and two manuka honeys originating from New Zealand. The researchers reported that 92.5% of a total of 2398 strains exhibited antimicrobial activity [42]. One of the isolates, identified as Paenibacillus polymyxa, showed a broad range of antibacterial activity against Gram−positive and −negative bacteria including P. larvae ssp. larvae ATCC 25747 and foodborne pathogens such as Bacillus cereus F4552 and Escherichia coli O157:H7 ATCC 43895 [43]. Zulkhairi Amin and coworkers (2020) revealed probiotic properties, including the production of antibacterial metabolites, of Bacillus spp. strains isolated from honey of the stingless bee Heterotrigona itama [44]. Khalili Samani et al. (2021) isolated several bacteriocin−producing strains of Bacillus spp. and Gram−positive cocci from the samples of Iranian honey. In contrast to the bacteriocins produced by these isolates, most of produced metabolites characterized in this study that were BP− and BB−derived strains that exhibited activity against both Gram− positive and Gram−negative bacteria [45]. However, to date, Bacillus spp.producing antimicrobial agents have not been isolated from BB or BP. A small amount of honey is added to BP before biotransformation, and this could be the source of Bacillus spp. in the final product—BB. On the other hand, bacteria of the genus Bacillus are common in the environment and could be present on the surfaces of pollen grains collected by bee workers.
Important and interesting information provided in this study included the investigation of enzymatic potential of the isolates. The production of lipases, cellulases, and proteinases were most common among tested strains. It can be assumed that these activities were essential for the “extraction” of basic food ingredients: amino acids, fatty acids, and glucose from components of pollen grains—proteins, lipids, and cellulose. These activities also improve the nutritional value of BB through the pre−digestion of biopolymers (e.g., cellulose and proteins), which is important for bees and human consumers. Moreover, the release of aliphatic acids from lipids can be important for the preservation of BB. Markiewicz−Żukowska and coworkers (2013) identified aliphatic acids as important antimicrobial components of BB, and unsaturated, α−linolenic, linoleic, oleic, and 11,14,17−eicosatrienoic acids formed more than a half of them (40.63 ± 4.5%) [46]. Neither β−galactosidase nor laccase are crucial for surviving in BB or BP, so these activities are rarely observed or not observed at all. Surprisingly, a relatively low percentage level of alpha amylase positive isolates was identified. Starch is important component of BP, and only 3 out of 81 tested strains exhibited strong potential for the hydrolysis of this polysaccharide. To our knowledge, the enzymatic potential of bacteria, other than LAB, isolated from BB or BP had not been investigated to date. Most of bacteria that belong to the genus Bacillus are not harmful to mammalians, with the exception of B. cereus and B. anthracis. Thus, the strains isolated from bee products, including BB or BP, can be considered sources of antimicrobials or enzymes. Moreover, they are also suitable candidates for probiotic bacteria [44].
4. Materials and Methods
4.1. Essential Chemical Reagents and Growth Media
All chemicals and growth media were purchased from commercial sources. LB broth and LB agar medium were bought from A&A Biotechnology (Gdynia; Poland). Mannitol salt phenol−red agar, Spirit blue agar, tributyrin agar, skimmed milk, starch, carboxymethylcellulose, guaiacol, X−gal (5−bromo−4−chloro−3−indolyl−β−D−galactopyranoside), Tween 80, cottonseed oil, Lugol’s solution, Congo red dye, and PBS tablets (pH 7.4) were purchased from Merck (Darmstadt, Germany). Ultrapure H2O (18.0 MΩ) was produced with the Milli−Q Advantage A10 system (Millipore, Billerica, MA, USA).
4.2. Bee Pollen and Bee Bread Samples and Isolation of Bacterial Strains
The samples of bee pollen (n = 4) and bee bread (n = 5) were provided by Polish apiaries. All samples of BP were dried (to protect the product against microbial spoilage). The BB samples were directly recovered from honeycombs in late summer or autumn 2019; thus, only mature bee bread was used for the study. All products were not older than six months counting from the date of harvesting to the date of using them for the experiment. The samples of BP were stored in dark conditions at ambient temperature, and BB was kept refrigerated at 4 °C. The suspensions of BP and BB in sterile deionized water at a 1:10 (w/v) ratio were performed for sample preparation. Subsequently, 100 µL of each suspension were streaked on the LB (Luria–Bertani) agar medium. The plates with inoculated agar medium were incubated at 37 °C for 24 h. Thereafter, the growing colonies were enumerated, and the level of microbial contamination of BB and BP samples (CFU/g of the product) was calculated. Each colony was individually transferred onto new Petri dish with an LB agar medium and incubated overnight at 37 °C. Then, a collection of isolates from BP and BB was obtained for further investigation.
4.3. Growth Inhibitory Assay
For assessing the antimicrobial activity of isolated bacteria, the colonies from the collection were transferred with sterile pipette tips onto LB agar plates inoculated with reference strains: S. aureus ATCC 253923, S. aureus ATCC 29213, S. epidermidis ATCC 12228, E. coli ATCC 25922, and P. aeruginosa ATCC 27853. Reference indicator strains were inoculated using a sterile cotton swab soaked in a diluted suspension of each tested strain prepared in a phosphate buffered solution (final optical density of each solution OD600 = 0.1; approximately 1–5 × 108 CFU/mL). Agar plates were incubated for 24 h at 37 °C. Thereafter, the presence and size of the GIZs of indicator strains were observed and recorded. The antimicrobial activity of each isolate was determined on the basis of the sizes of growth inhibition zones of indicatory strains observed around the colonies of bacteria isolated from BP or BB. All tests were performed in at least triplicate. Isolated bacteria with antimicrobial activity against reference strains were cataloged for further investigation.
4.4. Investigation of Enzymatic Activity of Isolated Strains
The bacterial isolates from BP and BB were tested to confirm or exclude their ability to produce enzymes from the group of hydrolases such as proteases, cellulases, amylases, esterases, lipases, laccases, and β−galactosidases. Two aspects were taken into account for the selection of the set of enzymatic activities that were investigated: the chemical composition of the BP (and therefore the substances that are available for the bacteria) and the industrial relevance of the enzymes regarding their application in industry. In order to determine the individual hydrolytic activities of isolates, the following media were applied: LB agar with skimmed milk (1.5% w/v) for proteolytic activity, LB agar with carboxymethylcellulose (2% w/v) for cellulolytic activity, LB agar with starch (2% w/v) for amylases activity, LB agar with guaiacol (100 µL/L) for laccase activity, LB agar with X−gal (20 mg/L) for β−galactosidase activity, a Tributyrin agar with neutral tributyrin (10 g/L) for esterolytic activity and a Spirit blue agar supplemented with 30 mL/L of lipase substrate (400 mL of warm distilled water, 1 mL of Tween 80, and 100 mL of cottonseed oil) for lipase activity. Using sterile pipette tips, each of isolates was applied on the appropriate agar medium in triplicate and incubated for 24 h at 37 °C. Following incubation, the appearance of halos around the colonies was observed for the confirmation of proteinase, esterase, and lipase activities. Halo zones around colonies indicated amylases and cellulolytic activities. However, the confirmation of the production of these enzymes required the flooding of the agar medium with Lugol’s solution or Congo red solution, respectively. Staining the growing colonies with blue or brownish dye was able to confirm the production of beta−galactosidase or laccase, respectively.
4.5. Identification of Bacterial Species of Isolates That Exhibited Antagonistic Activity against Selected Pathogenic Microorganisms
The identification of the isolates that exhibited antagonistic activity against selected pathogenic bacteria was executed by sequencing of the 16S rRNA gene. The DNA was isolated using Genomic Mini AX Bacteria+ (A&A Biotechnology, Gdynia, Poland) according to the protocol purchased from the manufacturer of the kit.
The PCR amplification of the targeted gene was determined with a pair of primers:
rP1 5′ CCCGGGATCCAAGCTTAGAGTTTGATCCTGGCTCAG 3′
Fd2 5′ CCCAATTCGTCGACAACACGGCTACCTTGTTACGACTT 3′
The amplified products sequencing was carried out by Macrogen (Amsterdam, the Netherlands). The purification of the amplified gene coding for 16S rRNA was executed using the enzymatic Post−PCR Immediate Cleanup (EPPiC) purification kit (A&A Biotechnology, Gdynia, Poland) following the protocol provided by the producer.
4.6. DNA Sequence Analysis
BLAST (Basic Local Alignment Search Tool) was used to the sequence analyses. Multiple sequence alignment in the MEGA X software was performed using the MUSCLE algorithm. The phylogenetic tree was constructed from the 16S rRNA sequences from previously generated FASTA sequence using the MEGA X software. The phylogenetic tree was assembled using the neighbor−joining method and sorting by distance.
5. Conclusions
Bacteria of the genus Bacillus have been identified as most important component of mature and stored BB, as well as dried BP. Moreover, the outcomes of the study revealed that BP and BB can be considered to be potential sources of bacteria producing antimicrobial agents and/or enzymes of particular industrial importance. Of course, additional research is required to verify this hypothesis, but the results of preliminary studies are promising.
Author Contributions
Conceptualization, P.S., R.W.W. and K.P.; methodology, K.P. and P.S.; formal analysis, P.S., K.P. and R.W.W.; investigation, K.P., J.W. and P.S.; data curation, K.P. and P.S.; software, K.P. and P.S.; writing—original draft preparation, P.S., K.P. and R.W.W.; writing—review and editing, P.S. and R.W.W.; visualization, K.P.; supervision, P.S. and R.W.W.; project administration, P.S.; funding acquisition, P.S. All authors have read and agreed to the published version of the manuscript.
Funding
The studies were financed by the grant2015/18/E/NZ6/00700 from the National Science Centre, Poland.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We are grateful to the company IGO PHU Maria Borowik from Lublin and beekeepers from the Apiary Hawran located in Miłogoszcz for some of the BP and BB samples delivered for our research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Combarros−Fuertes, P.; Fresno, J.M.; Estevinho, M.M.; Sousa−Pimenta, M.; Tornadijo, M.E.; Estevinho, L.M. Honey: Another Alternative in the Fight against Antibiotic−Resistant Bacteria? Antibiotics 2020, 9, 774. [Google Scholar] [CrossRef]
- Khan, S.U.; Anjum, S.I.; Rahman, K.; Ansari, M.J.; Khan, W.U.; Kamal, S.; Khattak, B.; Muhammad, A.; Khan, H.U. Honey: Single food stuff comprises many drugs. Saudi J. Biol. Sci. 2018, 25, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Silva−Carvalho, R.; Baltazar, F.; Almeida−Aguiar, C. Propolis: A complex natural product with a plethora of biological activities that can be explored for drug development. Evid. Based Complement. Alternat. Med. 2015, 2015, 206439. [Google Scholar] [CrossRef]
- Rojczyk, E.; Klama−Baryła, A.; Łabuś, W.; Wilemska−Kucharzewska, K.; Kucharzewski, M. Historical and modern research on propolis and its application in wound healing and other fields of medicine and contributions by Polish studies. J. Ethnopharmacol. 2020, 15, 113159. [Google Scholar] [CrossRef] [PubMed]
- Didaras, N.A.; Karatasou, K.; Dimitriou, T.G.; Amoutzias, G.D.; Mossialos, D. Antimicrobial Activity of Bee−Collected Pollen and Beebread: State of the Art and Future Perspectives. Antibiotics 2020, 9, 811. [Google Scholar] [CrossRef] [PubMed]
- Kwakman, P.H.; te Velde, A.A.; de Boer, L.; Speijer, D.; Vandenbroucke−Grauls, C.M.; Zaat, S.A. How honey kills bacteria. FASEB J. 2010, 24, 2576–2582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brudzynski, K. A current perspective on hydrogen peroxide production in honey. Food Chem. 2020, 332, 127229. [Google Scholar] [CrossRef] [PubMed]
- Brudzynski, K.; Sjaarda, C.P. Colloidal structure of honey and its influence on antibacterial activity. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2063–2080. [Google Scholar] [CrossRef]
- Brudzynski, K.; Abubaker, K.; Miotto, D. Unraveling a mechanism of honey antibacterial action: Polyphenol/h(2)o(2)−induced oxidative effect on bacterial cell growth and on DNA degradation. Food Chem. 2012, 133, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Brudzynski, K.; Miotto, D.; Kim, L.; Sjaarda, C.; Maldonado−Alvarez, L.; Fukś, H. Active macromolecules of honey form colloidal particles essential for honey antibacterial activity and hydrogen peroxide production. Sci. Rep. 2017, 7, 7637. [Google Scholar] [CrossRef] [Green Version]
- Bucekova, M.; Jardekova, L.; Juricova, V.; Bugarova, V.; Di Marco, G.; Gismondi, A.; Leonardi, D.; Farkasovska, J.; Godocikova, J.; Laho, M.; et al. Antibacterial Activity of Different Blossom Honeys: New Findings. Molecules 2019, 24, 1573. [Google Scholar] [CrossRef] [Green Version]
- Grecka, K.; Kuś, P.M.; Worobo, R.W.; Szweda, P. Study of the Anti−Staphylococcal Potential of Honeys Produced in Northern Poland. Molecules 2018, 23, 260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mavric, E.; Wittmann, S.; Barth, G.; Henle, T. Identification and quantification of methylglyoxal as the dominant antibacterial constituent of manuka (Leptospermum scoparium) honeys from New Zealand. Mol. Nutr. Food Res. 2008, 52, 483–489. [Google Scholar] [CrossRef]
- Irish, J.; Blair, S.; Carter, D.A. The antibacterial activity of honey derived from Australian flora. PLoS ONE 2011, 6, e18229. [Google Scholar] [CrossRef] [PubMed]
- Zainol, M.I.; Mohd Yusoff, K.; Mohd Yusof, M.Y. Antibacterial activity of selected Malaysian honey. BMC Complement. Altern. Med. 2013, 13, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Przybyłek, I.; Karpiński, T.M. Antibacterial Properties of Propolis. Molecules 2019, 24, 2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grecka, K.; Kuś, P.M.; Okińczyc, P.; Worobo, R.W.; Walkusz, J.; Szweda, P. The Anti−Staphylococcal Potential of Ethanolic Polish Propolis Extracts. Molecules 2019, 24, 1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gucwa, K.; Kusznierewicz, B.; Milewski, S.; Van Dijck, P.; Szweda, P. Antifungal Activity and Synergism with Azoles of Polish Propolis. Pathogens 2018, 7, 56. [Google Scholar] [CrossRef] [Green Version]
- Curuțiu, C.; Dițu, L.M.; Grumezescu, A.M.; Holban, A.M. Polyphenols of Honeybee Origin with Applications in Dental Medicine. Antibiotics 2020, 9, 856. [Google Scholar] [CrossRef]
- Ahmad, S.; Campos, M.G.; Fratini, F.; Altaye, S.Z.; Li, J. New Insights into the Biological and Pharmaceutical Properties of Royal Jelly. Int. J. Mol. Sci. 2020, 21, 382. [Google Scholar] [CrossRef] [Green Version]
- Collazo, N.; Carpena, M.; Nuñez−Estevez, B.; Otero, P.; Simal−Gandara, J.; Prieto, M.A. Health Promoting Properties of Bee Royal Jelly: Food of the Queens. Nutrients 2021, 13, 543. [Google Scholar] [CrossRef]
- Fratini, F.; Cilia, G.; Turchi, B.; Felicioli, A. Beeswax: A minireview of its antimicrobial activity and its application in medicine. Asian Pac J. Trop. Med. 2016, 9, 839–843. [Google Scholar] [CrossRef] [PubMed]
- El−Seedi, H.; Abd El−Wahed, A.; Yosri, N.; Musharraf, S.G.; Chen, L.; Moustafa, M.; Zou, X.; Al−Mousawi, S.; Guo, Z.; Khatib, A.; et al. Antimicrobial Properties of Apis mellifera’s Bee Venom. Toxins 2020, 12, 451. [Google Scholar] [CrossRef]
- Kostić, A.Ž.; Milinčić, D.D.; Barać, M.B.; Ali Shariati, M.; Tešić, Ž.L.; Pešić, M.B. The Application of Pollen as a Functional Food and Feed Ingredient—The Present and Perspectives. Biomolecules 2020, 10, 84. [Google Scholar] [CrossRef] [Green Version]
- Kieliszek, M.; Piwowarek, K.; Kot, A.; Błażejak, S.; Chlebowska−Śmigiel, A.; Wolska, I. Pollen and bee bread as new health−oriented products: A review. Trends Food Sci. Technol. 2018, 71, 170–180. [Google Scholar] [CrossRef]
- Fatrcová−Šramková, K.; Nôžková, J.; Máriássyová, M.; Kačániová, M. Biologically active antimicrobial and antioxidant substances in the Helianthus annuus L. bee pollen. J. Environ. Sci. Health Part B 2016, 51, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Didaras, N.A.; Kafantaris, I.; Dimitriou, T.G.; Mitsagga, C.; Karatasou, K.; Giavasis, I.; Stagos, D.; Amoutzias, G.D.; Hatjina, F.; Mossialos, D. Biological Properties of Bee Bread Collected from Apiaries Located across Greece. Antibiotics 2021, 10, 555. [Google Scholar] [CrossRef] [PubMed]
- Gilliam, M. Microbiology of pollen and bee bread: The yeasts. Apidologie 1979, 10, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Disayathanoowat, T.; Li, H.; Supapimon, N.; Suwannarach, N.; Lumyong, S.; Chantawannakul, P.; Guo, J. Different Dynamics of Bacterial and Fungal Communities in Hive−Stored Bee Bread and Their Possible Roles: A Case Study from Two Commercial Honey Bees in China. Microorganisms 2020, 8, 264. [Google Scholar] [CrossRef] [Green Version]
- Vásquez, A.; Olofsson, T.C. The lactic acid bacteria involved in the production of bee pollen and bee bread. J. Apic. Res. 2009, 48, 189–195. [Google Scholar] [CrossRef]
- Pełka, K.; Otłowska, O.; Worobo, R.W.; Szweda, P. Bee Bread Exhibits Higher Antimicrobial Potential Compared to Bee Pollen. Antibiotics 2021, 10, 125. [Google Scholar] [CrossRef]
- Brudzynski, K. Honey as an Ecological Reservoir of Antibacterial Compounds Produced by Antagonistic Microbial Interactions in Plant Nectars, Honey and Honey Bee. Antibiotics 2021, 10, 551. [Google Scholar] [CrossRef] [PubMed]
- Casteel, D.B. The behavior of the honey bee in pollen collection. USDA Bur. Entomol. Bull. 1912, 121, 1–36. [Google Scholar]
- Komosińska−Vassev, K.; Olczyk, P.; Kaźmierczak, J.; Mencner, Ł.; Olczyk, K. Bee pollen: Chemical composition and therapeutic application. Evid. Based Complement. Altern. Med. 2015, 2015, 297425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowak, A.; Szczuka, D.; Górczyńska, A.; Motyl, I.; Kręgiel, D. Characterization of Apis mellifera Gastrointestinal Microbiota and Lactic Acid Bacteria for Honeybee Protection—A Review. Cells 2021, 10, 701. [Google Scholar] [CrossRef] [PubMed]
- Iorizzo, M.; Lombardi, S.J.; Ganassi, S.; Testa, B.; Ianiro, M.; Letizia, F.; Succi, M.; Tremonte, P.; Vergalito, F.; Cozzolino, A.; et al. Antagonistic Activity against Ascosphaera apis and Functional Properties of Lactobacillus kunkeei Strains. Antibiotics 2020, 9, 262. [Google Scholar] [CrossRef] [PubMed]
- Iorizzo, M.; Testa, B.; Lombardi, S.J.; Ganassi, S.; Ianiro, M.; Letizia, F.; Succi, M.; Tremonte, P.; Vergalito, F.; Cozzolino, A.; et al. Antimicrobial Activity against Paenibacillus larvae and Functional Properties of Lactiplantibacillus plantarum Strains: Potential Benefits for Honeybee Health. Antibiotics 2020, 9, 442. [Google Scholar] [CrossRef]
- Detry, R.; Simon−Delso, N.; Bruneau, E.; Daniel, H.-M. Specialisation of Yeast Genera in Different Phases of Bee Bread Maturation. Microorganisms 2020, 8, 1789. [Google Scholar] [CrossRef] [PubMed]
- Pajor, M.; Worobo, R.W.; Milewski, S.; Szweda, P. The Antimicrobial Potential of Bacteria Isolated from Honey Samples Produced in the Apiaries Located in Pomeranian Voivodeship in Northern Poland. Int. J. Environ. Res. Public Health 2018, 15, 2002. [Google Scholar] [CrossRef] [Green Version]
- Pajor, M.; Xiong, Z.R.; Worobo, R.W.; Szweda, P. Paenibacillus alvei MP1 as a Producer of the Proteinaceous Compound with Activity against Important Human Pathogens, Including Staphylococcus aureus and Listeria monocytogenes. Pathogens 2020, 9, 319. [Google Scholar] [CrossRef]
- Pajor, M.; Sogin, J.; Worobo, R.; Szweda, P. Draft genome sequence of antimicrobial producing Paenibacillus alvei strain MP1 reveals putative novel antimicrobials. BMC Res. Notes 2020, 13, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Churey, J.J.; Worobo, R.W. Antimicrobial activity of bacterial isolates from different floral sources of honey. Int. J. Food Microbiol. 2008, 126, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Churey, J.J.; Worobo, R.W. Isolation and characterization of a protective bacterial culture isolated from honey active against American Foulbrood disease. FEMS Microbiol. Lett. 2009, 296, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Zulkhairi Amin, F.A.; Sabri, S.; Ismail, M.; Chan, K.W.; Ismail, N.; Mohd Esa, N.; Mohd Lila, M.A.; Zawawi, N. Probiotic Properties of Bacillus Strains Isolated from Stingless Bee (Heterotrigona itama) Honey Collected across Malaysia. Int. J. Environ. Res. Public Health 2020, 17, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalili Samani, M.; Noormohammadi, Z.; Fazeli, M.R. Bacteriocin activity of various iranian honey-associated bacteria and development of a simple medium for enhanced bacteriocin activity. J. Environ. Health Sci. Eng. 2021, 19, 427–435. [Google Scholar] [CrossRef]
- Markiewicz−Żukowska, R.; Naliwajko, S.; Bartosiuk, E.; Moskwa, J.; Isidorov, V.; Soroczyńska, J.; Borawska, M. Chemical composition and antioxidant activity of beebread, and its influence on the glioblastoma cell line (U87MG). J. Apic. Sci. 2013, 57, 147–157. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
An example of antagonistic interaction between isolated strains (strains 1–21 were isolated from the product BB19, and strains 1 and 2 were derived from the product BB3) and indicatory/reference strains of pathogenic bacteria: (a) S. aureus ATCC 25923, (b) S. aureus ATCC 29213, (c) S. epidermidis ATCC 12228, and (d) E. coli ATCC 25922. Interactions with P. aeruginosa ATCC 27853 were a separately analyzed (results not presented).
Figure 1.
An example of antagonistic interaction between isolated strains (strains 1–21 were isolated from the product BB19, and strains 1 and 2 were derived from the product BB3) and indicatory/reference strains of pathogenic bacteria: (a) S. aureus ATCC 25923, (b) S. aureus ATCC 29213, (c) S. epidermidis ATCC 12228, and (d) E. coli ATCC 25922. Interactions with P. aeruginosa ATCC 27853 were a separately analyzed (results not presented).

Figure 2.
Phylogenetic tree of 34 strains that exhibited antagonistic potential against at least of one reference strain of pathogenic bacteria. MUSCLE multiple alignment/construction was done using the neighbor−joining method.
Figure 2.
Phylogenetic tree of 34 strains that exhibited antagonistic potential against at least of one reference strain of pathogenic bacteria. MUSCLE multiple alignment/construction was done using the neighbor−joining method.

Figure 3.
Enzymatic activity of isolates. Selected agar plates are presented with observed (a) proteolytic, (b) amylolytic, (c) lipolytic, (d) esterolytic, and (e) cellulolytic activity. In the upper part of the plates, there are strains that did not exhibit enzymatic activity, while in the lower part of the plates, there are strains capable of producing hydrolases.
Figure 3.
Enzymatic activity of isolates. Selected agar plates are presented with observed (a) proteolytic, (b) amylolytic, (c) lipolytic, (d) esterolytic, and (e) cellulolytic activity. In the upper part of the plates, there are strains that did not exhibit enzymatic activity, while in the lower part of the plates, there are strains capable of producing hydrolases.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Level of microbial contamination of investigated BP and BB samples and the antagonistic activity of isolates against reference strains of pathogenic bacteria.
Table 1.
Level of microbial contamination of investigated BP and BB samples and the antagonistic activity of isolates against reference strains of pathogenic bacteria.
| Sample | No. of Colonies | CFU/g of Product | Activity against S. aureus ATCC 25923 | Activity against S. aureus ATCC 29213 | Activity against S. epidermidis ATCC 12228 | Activity against E. coli ATCC 25922 | Activity against P. aeruginosa ATCC 27853 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. of Colonies | % | No. of Colonies | % | No. of Colonies | % | No. of Colonies | % | No. of Colonies | % | |||
| BP3 | 14 | 1400 | 6 | 42.86 | 6 | 42.86 | 5 | 35.71 | 5 | 35.71 | 7 | 50.00 |
| BP15 | 5 | 500 | 2 | 40.00 | 2 | 40.00 | 2 | 40.00 | 2 | 40.00 | 3 | 60.00 |
| BP12 | 1 | 100 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 |
| BP20 | 15 | 1500 | 6 | 40.00 | 7 | 46.67 | 3 | 20.00 | 1 | 6.67 | 7 | 46.67 |
| BB3 | 2 | 200 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 |
| BB6 | 13 | 1300 | 1 | 7.69 | 2 | 15.38 | 1 | 7.69 | 1 | 7.69 | 2 | 15.38 |
| BB10 | 6 | 600 | 3 | 50.00 | 3 | 50.00 | 3 | 50.00 | 1 | 16.67 | 3 | 50.00 |
| BB15 | 4 | 400 | 1 | 25.00 | 1 | 25.00 | 0 | 0.00 | 0 | 0.00 | 1 | 25.00 |
| BB19 | 21 | 2100 | 8 | 38.10 | 8 | 38.10 | 8 | 38.10 | 5 | 23.81 | 9 | 42.86 |
| TOTAL | 81 | 27 | 33.33 | 29 | 35.80 | 22 | 27.20 | 15 | 18.52 | 32 | 39.51 | |
Table 2.
Species classification based on the BLAST analysis of 16S rRNA gene sequences and antagonistic activity against reference strains of pathogenic bacteria.
Table 2.
Species classification based on the BLAST analysis of 16S rRNA gene sequences and antagonistic activity against reference strains of pathogenic bacteria.
| Sample | Species Classification * | Exhibited Activity | ||||
|---|---|---|---|---|---|---|
| S. aureus ATCC 25923 | S. aureus ATCC 29213 | S. epidermidis ATCC 12228 | E. coli ATCC 25922 | P. aeruginosa ATCC 27853 | ||
| BP3.2 | Bacillus spp. (pumilus, zhangzhouensis) | ++ | +++ | +++ | +++ | +++ |
| BP3.3 | Bacillus spp. (pumilus, zhangzhouensis) | + | ++ | ++ | ++ | ++ |
| BP3.7 | Bacillus spp. (pumilus, zhangzhouensis) | + | + | + | ++ | ++ |
| BP3.10 | Bacillus spp. (pumilus, zhangzhouensis) | +++ | +++ | +++ | +++ | +++ |
| BP3.11 | Bacillus spp. (altitudinis, stratosphericus) | + | + | − | − | +++ |
| BP3.12 | Bacillus spp. (licheniformis, aerius) | − | − | − | − | ++ |
| BP3.13 | Bacillus spp. (pumilus, zhangzhouensis) | +++ | +++ | +++ | +++ | +++ |
| BP15.1 | Bacillus spp. (lichemiformis) | − | − | − | − | +++ |
| BP15.3 | Bacillus spp. (pumilus, zhangzhouensis) | ++ | ++ | +++ | +++ | +++ |
| BP15.4 | Bacillus spp. (altitudinis, stratosphericus) | +++ | +++ | +++ | +++ | ++ |
| BP20.1 | Bacillus spp. (safensis, pumilus) | + | + | − | − | ++ |
| BP20.3 | Bacillus spp. (altitudinis, stratosphericus) | + | + | − | − | ++ |
| BP20.4 | Bacillus spp. (altitudinis, stratosphericus) | + | + | − | − | + |
| BP20.6 | Bacillus spp. (altitudinis, stratosphericus) | − | + | − | − | +++ |
| BP20.7 | Bacillus spp. (altitudinis, stratosphericus) | + | + | +++ | − | ++ |
| BP20.9 | Bacillus spp. (altitudinis, stratosphericus) | +++ | ++ | ++ | ++ | ++ |
| BP20.15 | Bacillus spp. (subtilis) | +++ | ++ | + | − | +++ |
| BB6.2 | Bacillus spp. (pumilus, zhangzhouensis) | +++ | ++ | +++ | ++ | ++ |
| BB6.5 | Bacillus spp. (altitudinis, stratosphericus) | − | ++ | − | − | + |
| BB10.1 | Bacillus spp. (subtilis) | +++ | +++ | +++ | − | +++ |
| BB10.3 | Bacillus spp. (pumilus, zhangzhouensis) | + | + | + | − | ++ |
| BB10.6 | Bacillus spp. (pumilus, zhangzhouensis) | +++ | +++ | +++ | +++ | +++ |
| BB15.3 | Bacillus spp. (altitudinis, stratosphericus) | + | ++ | − | − | +++ |
| BB19.2 | Bacillus spp. (pumilus, zhangzhouensis) | − | − | ++ | − | − |
| BB19.7 | Bacillus spp. (pumilus, zhangzhouensis) | +++ | ++ | +++ | +++ | +++ |
| BB19.9 | Bacillus spp. (licheniformis, aerius) | − | − | − | − | ++ |
| BB19.10 | Bacillus spp. (altitudinis, aerius) | + | + | − | − | + |
| BB19.11 | Bacillus spp. (licheniformis, paralicheniformis) | − | − | − | − | +++ |
| BB19.12 | Bacillus spp. (pumilus, zhangzhouensis) | ++ | ++ | +++ | + | +++ |
| BB19.13 | Bacillus spp. (pumilus, zhangzhouensis) | +++ | ++ | +++ | − | ++ |
| BB19.15 | Bacillus spp. (pumilus, zhangzhouensis) | ++ | +++ | +++ | +++ | +++ |
| BB19.17 | Bacillus spp. (pumilus, zhangzhouensis) | +++ | +++ | ++ | − | +++ |
| BB19.19 | Bacillus spp. (pumilus, zhangzhouensis) | +++ | +++ | ++ | ++ | − |
| BB19.21 | Bacillus spp. (pumilus, zhangzhouensis) | +++ | +++ | +++ | +++ | +++ |
*—sequences of gene coding for 16S rRNA of different species of the genus Bacillus exhibit high level of similarity. Thus, in most cases, two most possible species are proposed. The classification of antagonistic interaction as S—strong (+++); M—moderate (++); W—weak (+); or L—lack (−) was based on the measurement of the size of the growth inhibition zone (SGIZ) of indicatory strain counted from the edge of the colony of the investigated isolate. The following scale was used for the classification of antagonistic interactions: strong—SGIZ > 3 mm; moderate—SGIZ in the range from 1 to 3 mm; and weak—SGIZ ≤ 1 mm. The mean value of this parameter from three independent experiments was used for final classification of each strain tested.
Table 3.
Enzymatic activity of isolates.
| Isolates | Proteolytic Activity | Amylolytic Activity | Lipolytic Activity | Esterolytic Activity | Cellulolytic Activity | Presence of Beta−Galactosidase | Presence of Laccase | |
|---|---|---|---|---|---|---|---|---|
| 1 | BP3.1 | − | − | − | − | − | − | − |
| 2 | BP3.2 | +++ | − | + | − | ++ | − | − |
| 3 | BP3.3 | ++ | − | ++ | − | ++ | − | − |
| 4 | BP3.4 | − | − | − | − | − | − | − |
| 5 | BP3.5 | +++ | − | ++ | ++ | +++ | − | − |
| 6 | BP3.6 | + | − | + | − | + | − | − |
| 7 | BP3.7 | + | − | ++ | ++ | − | − | − |
| 8 | BP3.8 | − | − | − | − | − | − | − |
| 9 | BP3.9 | − | +++ | ++ | ++ | +++ | + | − |
| 10 | BP3.10 | +++ | − | ++ | − | ++ | − | − |
| 11 | BP3.11 | + | − | + | − | + | − | − |
| 12 | BP3.12 | − | − | +++ | + | +++ | + | − |
| 13 | BP3.13 | ++ | − | + | − | ++ | − | − |
| 14 | BP3.14 | + | − | + | − | + | − | − |
| 15 | BP12.1 | ++ | − | +++ | − | ++ | + | − |
| 16 | BP15.1 | − | − | +++ | − | ++ | − | − |
| 17 | BP15.2 | ++ | − | + | + | ++ | − | − |
| 18 | BP15.3 | +++ | − | ++ | − | ++ | − | − |
| 19 | BP15.4 | + | − | ++ | − | +++ | − | − |
| 20 | BP15.5 | − | − | ++ | − | − | − | − |
| 21 | BP20.1 | ++ | − | ++ | − | − | − | − |
| 22 | BP20.2 | ++ | − | +++ | − | + | + | − |
| 23 | BP20.3 | ++ | − | ++ | + | +++ | − | − |
| 24 | BP20.4 | ++ | − | ++ | + | ++ | − | − |
| 25 | BP20.5 | − | − | +++ | − | + | − | − |
| 26 | BP20.6 | +++ | − | ++ | + | ++ | − | − |
| 27 | BP20.7 | ++ | − | ++ | + | ++ | − | − |
| 28 | BP20.8 | ++ | − | +++ | − | − | + | − |
| 29 | BP20.9 | ++ | − | ++ | + | +++ | − | − |
| 30 | BP20.10 | ++ | − | − | − | − | − | − |
| 31 | BP20.11 | − | − | + | − | + | + | − |
| 32 | BP20.12 | − | − | + | ++ | − | − | − |
| 33 | BP20.13 | − | − | − | − | − | − | − |
| 34 | BP20.14 | − | − | − | − | − | − | − |
| 35 | BP20.15 | +++ | +++ | +++ | + | +++ | − | − |
| 36 | BB3.1 | − | − | ++ | − | +++ | − | − |
| 37 | BB3.2 | − | − | − | − | − | − | − |
| 38 | BB6.1 | − | − | ++ | − | − | + | − |
| 39 | BB6.2 | +++ | − | ++ | + | ++ | − | − |
| 40 | BB6.3 | − | − | ++ | − | − | + | − |
| 41 | BB6.4 | − | − | − | − | − | − | − |
| 42 | BB6.5 | − | − | +++ | +++ | ++ | − | − |
| 43 | BB6.6 | − | − | − | − | − | − | − |
| 44 | BB6.7 | − | − | + | − | − | − | − |
| 45 | BB6.8 | ++ | − | ++ | − | + | + | − |
| 46 | BB6.9 | − | − | +++ | − | − | − | − |
| 47 | BB6.10 | − | − | − | − | − | − | − |
| 48 | BB6.11 | − | − | + | ++ | +++ | − | − |
| 49 | BB6.12 | − | − | +++ | − | +++ | − | − |
| 50 | BB6.13 | − | − | ++ | + | ++ | − | − |
| 51 | BB10.1 | + | +++ | + | ++ | +++ | − | − |
| 52 | BB10.2 | − | − | +++ | − | ++ | − | − |
| 53 | BB10.3 | + | − | ++ | ++ | + | − | − |
| 54 | BB10.4 | − | − | +++ | − | ++ | − | − |
| 55 | BB10.5 | − | − | +++ | − | +++ | + | − |
| 56 | BB10.6 | ++ | − | + | ++ | ++ | − | − |
| 57 | BB15.1 | − | − | + | + | + | − | − |
| 58 | BB15.2 | − | − | + | − | + | − | − |
| 59 | BB15.3 | + | − | + | + | ++ | − | − |
| 60 | BB15.4 | − | − | +++ | − | +++ | − | − |
| 61 | BB19.1 | − | − | +++ | − | ++ | − | − |
| 62 | BB19.2 | +++ | − | ++ | + | + | − | − |
| 63 | BB19.3 | − | − | ++ | − | − | − | − |
| 64 | BB19.4 | − | − | ++ | − | − | − | − |
| 65 | BB19.5 | − | − | ++ | + | − | − | − |
| 66 | BB19.6 | + | − | ++ | − | ++ | − | − |
| 67 | BB19.7 | + | − | +++ | + | ++ | − | − |
| 68 | BB19.8 | − | − | ++ | − | − | − | − |
| 69 | BB19.9 | − | − | − | + | + | − | − |
| 70 | BB19.10 | ++ | − | ++ | − | +++ | − | − |
| 71 | BB19.11 | − | − | + | − | + | + | − |
| 72 | BB19.12 | +++ | − | ++ | + | +++ | − | − |
| 73 | BB19.13 | ++ | − | ++ | + | ++ | − | − |
| 74 | BB19.14 | − | − | + | − | − | − | − |
| 75 | BB19.15 | + | − | ++ | − | +++ | − | − |
| 76 | BB19.16 | − | − | − | − | − | − | − |
| 77 | BB19.17 | ++ | − | +++ | − | +++ | − | − |
| 78 | BB19.18 | − | − | ++ | − | − | − | − |
| 79 | BB19.19 | ++ | − | +++ | + | − | − | − |
| 80 | BB19.20 | − | − | ++ | − | ++ | − | − |
| 81 | BB19.21 | +++ | − | +++ | + | +++ | − | − |
| TOTAL | ||||||||
| S | 10 (12%) | 3 (4%) | 19 (23%) | 1 (1%) | 18 (22%) | 0 (0%) | 0 | |
| M | 18 (22%) | 0 | 33 (41%) | 8 (10%) | 23 (28%) | 0 (0%) | 0 | |
| W | 11 (14%) | 0 | 17 (21%) | 20 (25%) | 13 (16%) | 11 (14%) | 0 | |
| L | 42 (52%) | 78 (96%) | 12 (15%) | 52 (64%) | 27 (33%) | 70 (86%) | 81 (100%) | |
S—strong (+++); M—moderate (++); W—weak (+); L—lack (−).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pełka, K.; Worobo, R.W.; Walkusz, J.; Szweda, P. Bee Pollen and Bee Bread as a Source of Bacteria Producing Antimicrobials. Antibiotics 2021, 10, 713. https://doi.org/10.3390/antibiotics10060713
AMA Style
Pełka K, Worobo RW, Walkusz J, Szweda P. Bee Pollen and Bee Bread as a Source of Bacteria Producing Antimicrobials. Antibiotics. 2021; 10(6):713. https://doi.org/10.3390/antibiotics10060713
Chicago/Turabian StylePełka, Karolina, Randy W. Worobo, Justyna Walkusz, and Piotr Szweda. 2021. "Bee Pollen and Bee Bread as a Source of Bacteria Producing Antimicrobials" Antibiotics 10, no. 6: 713. https://doi.org/10.3390/antibiotics10060713
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.