Enzymatically Digested Food Waste Altered Fecal Microbiota But Not Meat Quality and Carcass Characteristics of Growing-Finishing Pigs

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Husbandry, Experimental Design, and Dietary Treatments
2.2. Carcass Characteristics and Fresh Meat Quality
2.2.1. Hot Carcass Weight (HCW), Estimated Lean, and Back-Fat
2.2.2. Water Holding Capacity and Objective Tenderness
2.2.3. Subjective Color, Marbling Score, Firmness Score, pH, and Moisture Content
2.3. Carcass Composition
2.4. Fecal Microbiota
2.5. Statistical Analysis
3. Results and Discussion
3.1. Carcass Characteristics and Meat Quality
3.2. Back-Fat Composition and Fatty Acids Profile
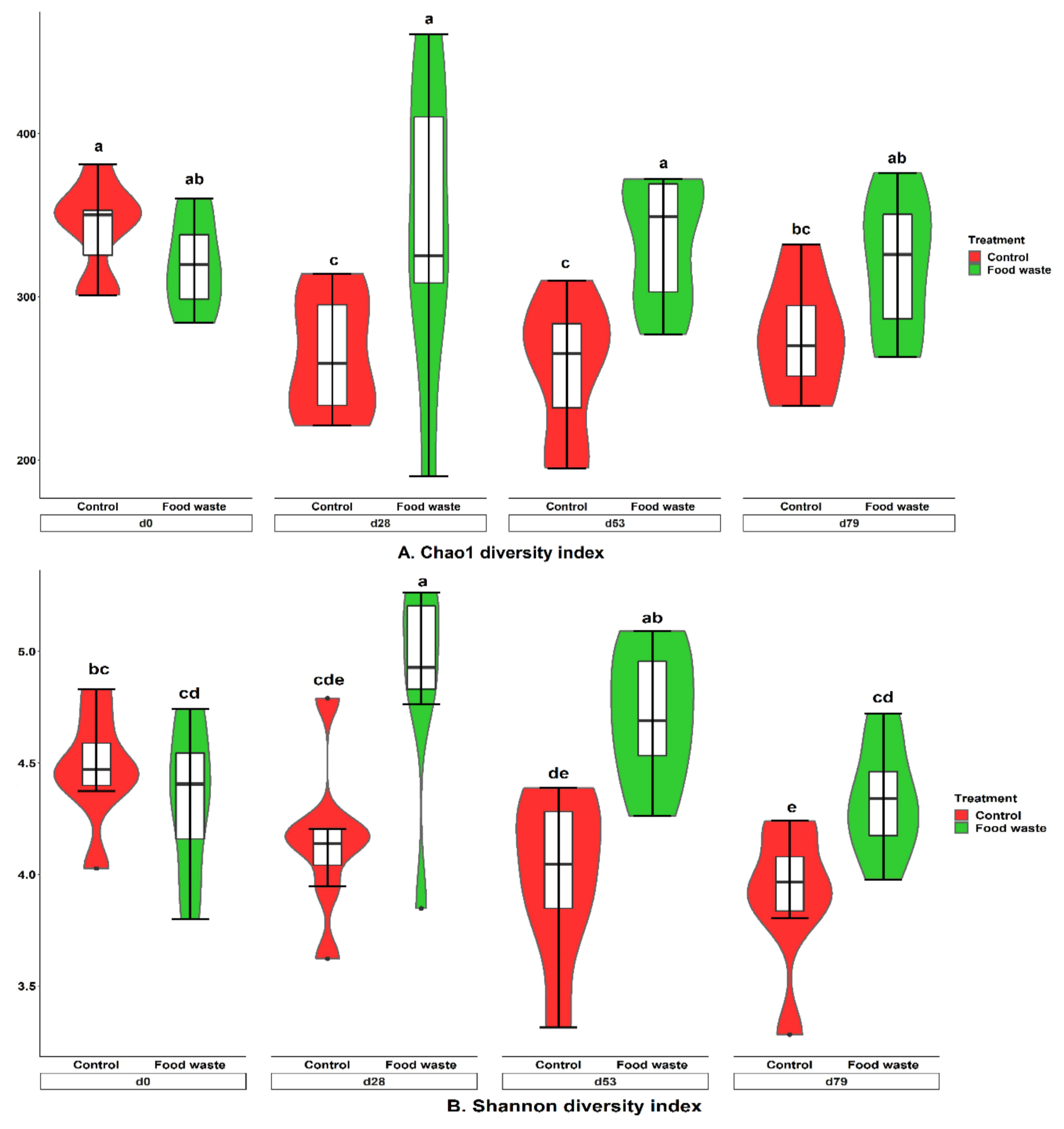
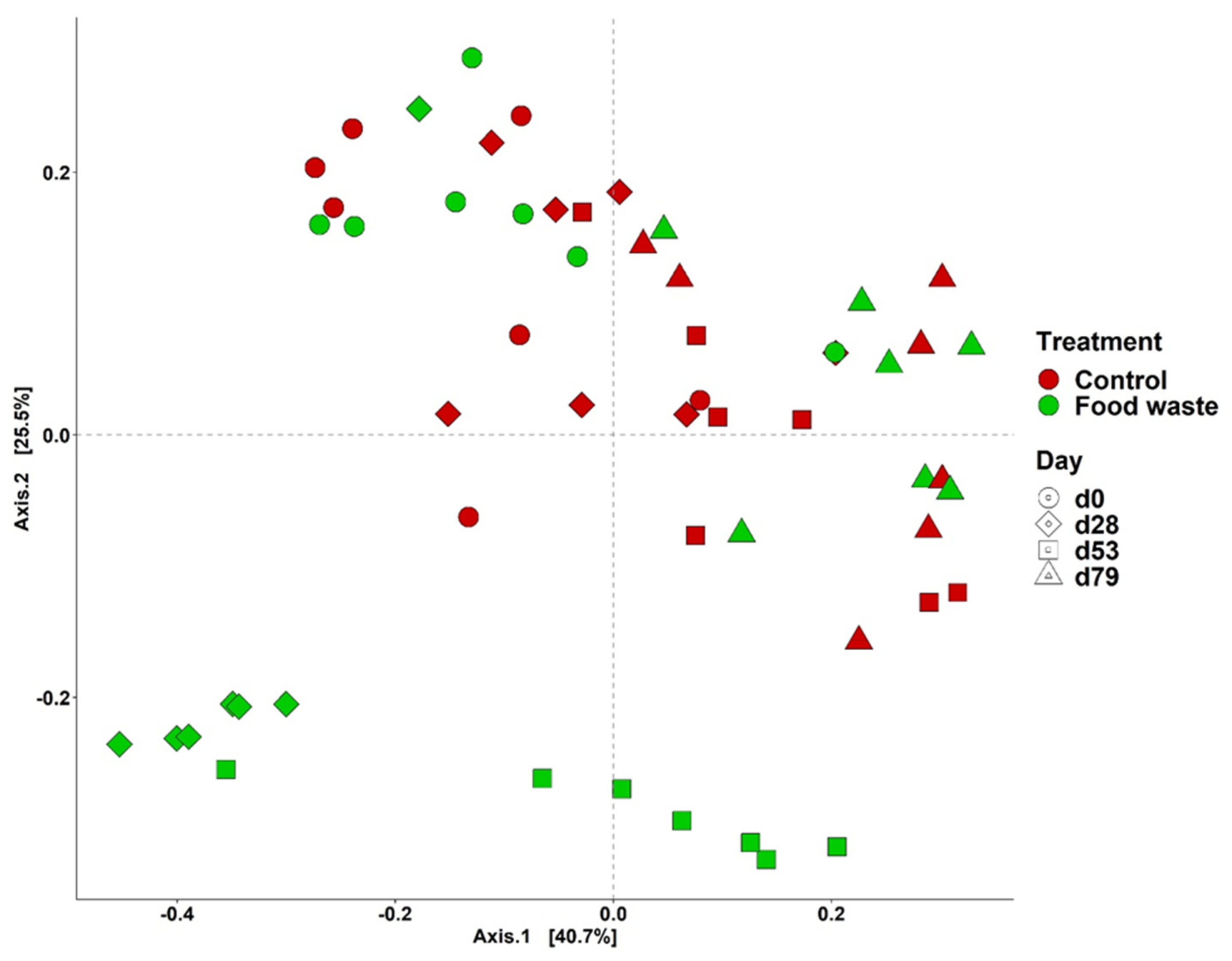
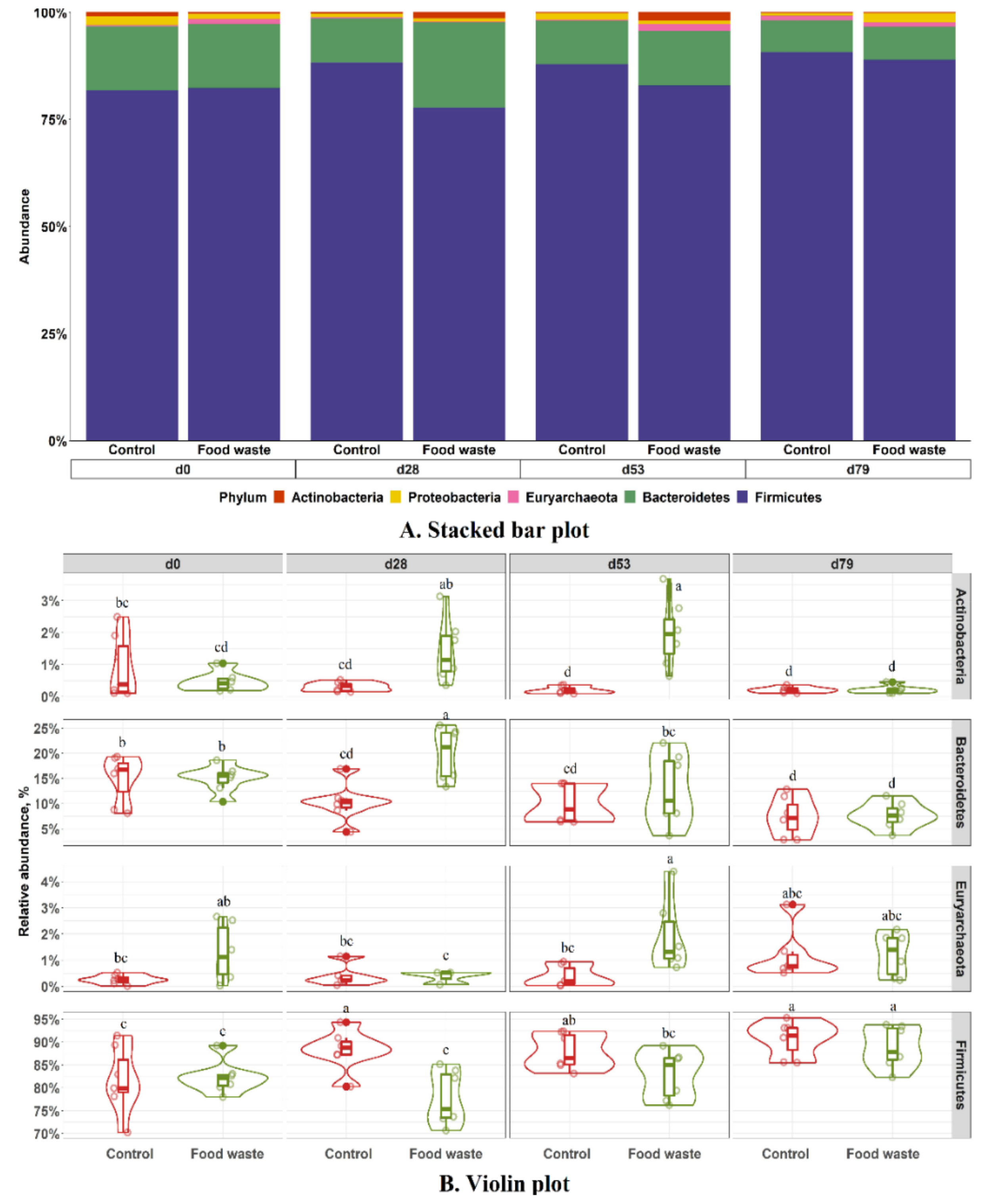
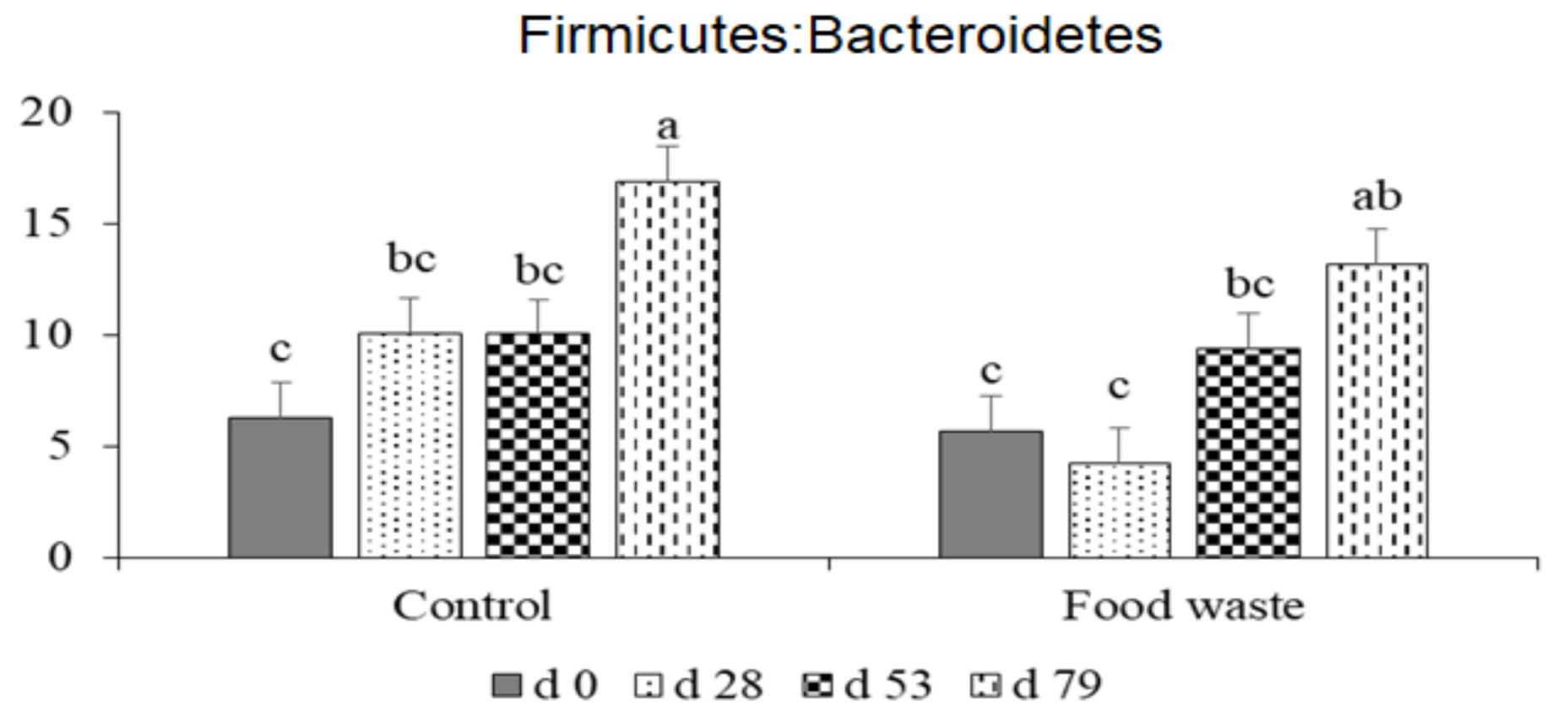
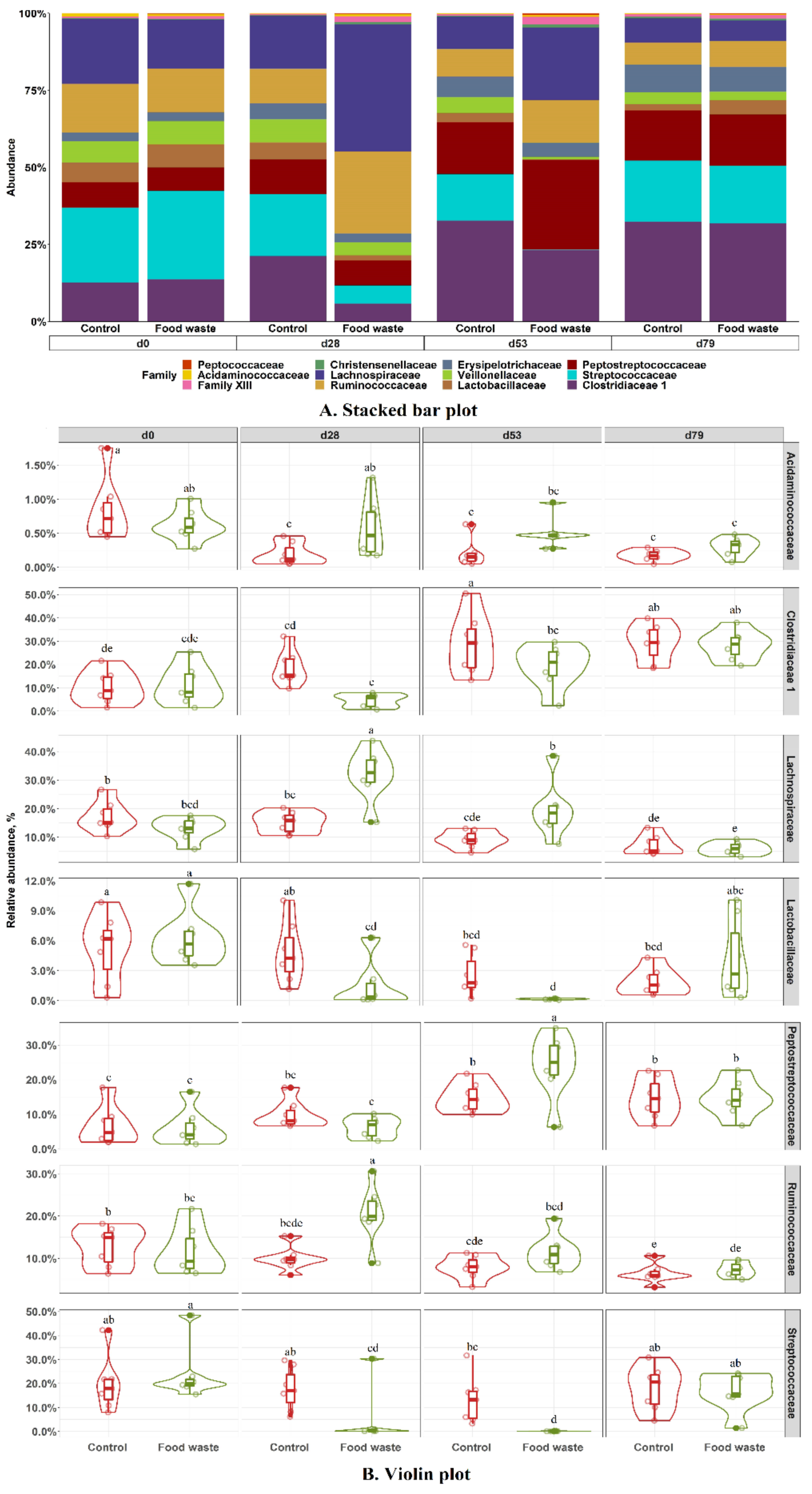
3.3. Fecal Microbiota
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. What is Food Loss and Food Waste? Available online: http://www.fao.org/food-loss-and-food-waste/en/ (accessed on 5 April 2019).
- US EPA. National Overview: Facts and Figures on Materials, Wastes and Recycling. Available online: https://www.epa.gov/facts-and-figures-about-materials-waste-and-recycling/national-overview-facts-and-figures-materials (accessed on 18 April 2019).
- Bingemer, H.G.; Crutzen, P.J. The production of methane from solid wastes. J. Geophys. Res. Atmos. 1987, 92, 2181–2187. [Google Scholar] [CrossRef]
- Gunders, D.; Bloom, J. Wasted: How America is Losing up to 40 Percent of its Food from Farm to Fork to Landfill; Natural Resources Defense Council: New York, NY, USA, 2017. [Google Scholar]
- Kim, J.-D.; Park, J.-S.; In, B.-H.; Kim, D.; Namkoong, W. Evaluation of pilot-scale in vessel composting for food waste treatment. J. Hazard. Mater. 2008, 154, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Shi, W.; Hong, J.; Zhang, F.; Chen, W. Life cycle assessment of food waste-based biogas generation. Renew. Sust. Energ. Rev. 2015, 49, 169–177. [Google Scholar] [CrossRef]
- Jinno, C.; He, Y.; Morash, D.; McNamara, E.; Zicari, S.; King, A.; Stein, H.H.; Liu, Y. Enzymatic digestion turns food waste into feed for growing pigs. Anim. Feed Sci. Technol. 2018, 242, 48–58. [Google Scholar] [CrossRef]
- Pandey, P.; Lejeune, M.; Biswas, S.; Morash, D.; Weimer, B.; Young, G. A new method for converting foodwaste into pathogen free soil amendment for enhancing agricultural sustainability. J. Clean. Prod. 2018, 112, 205–213. [Google Scholar] [CrossRef]
- Frese, S.A.; Parker, K.; Calvert, C.C.; Mills, D.A. Diet shapes the gut microbiome of pigs during nursing and weaning. Microbiome 2015, 3, 28–37. [Google Scholar] [CrossRef]
- Muegge, B.D.; Kuczynski, J.; Knights, D.; Clemente, J.C.; González, A.; Fontana, L.; Henrissat, B.; Knight, R.; Gordon, J.I. Diet drives convergence in gut microbiome functions across mammalian phylogeny and within humans. Science 2011, 332, 970–974. [Google Scholar] [CrossRef]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef]
- Pedersen, H.K.; Gudmundsdottir, V.; Nielsen, H.B.; Hyotylainen, T.; Nielsen, T.; Jensen, B.A.H.; Forslund, K.; Hildebrand, F.; Prifti, E.; Falony, G.; et al. Human gut microbes impact host serum metabolome and insulin sensitivity. Nature 2016, 535, 376–381. [Google Scholar] [CrossRef]
- Yan, H.; Potu, R.; Lu, H.; Almeida, V.V. de; Stewart, T.; Ragland, D.; Armstrong, A.; Adeola, O.; Nakatsu, C.H.; Ajuwon, K.M. Dietary fat content and fiber type modulate hind gut microbial community and metabolic markers in the pig. PLoS ONE 2013, 8, e59581. [Google Scholar] [CrossRef]
- Yang, H.; Xiang, Y.; Robinson, K.; Wang, J.; Zhang, G.; Zhao, J.; Xiao, Y. Gut microbiota is a major contributor to adiposity in pigs. Front. Microbiol. 2018, 9, 3045–3057. [Google Scholar] [CrossRef] [PubMed]
- National Research Council, Division on Earth and Life Studies, Board on Agriculture and Natural Resources, Committee on Nutrient Requirements of Swine. Nutrient Requirements of Swine, Eleventh Revised Edition; National Academic Press: Washington, DC, USA, 2012; ISBN 978-0-309-22423-9. [Google Scholar]
- AOAC. International Official Methods of Analysis of AOAC; AOAC: Gaithersburg, MD, USA, 2007. [Google Scholar]
- Little, K.L.; Bohrer, B.M.; Maison, T.; Liu, Y.; Stein, H.H.; Boler, D.D. Effects of feeding canola meal from high-protein or conventional varieties of canola seeds on growth performance, carcass characteristics, and cutability of pigs. J. Anim. Sci. 2015, 93, 1284–1297. [Google Scholar] [CrossRef] [PubMed]
- Boler, D.; Holmer, S.; Duncan, D.; Carr, S.; Ritter, M.; Stites, C.; Petry, D.; Hinson, R.; Allee, G.; McKeith, F. Fresh meat and further processing characteristics of ham muscles from finishing pigs fed ractopamine hydrochloride. J. Anim. Sci. 2011, 89, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Leick, C.M.; Puls, C.L.; Ellis, M.; Killefer, J.; Carr, T.R.; Scramlin, S.M.; England, M.B.; Gaines, A.M.; Wolter, B.F.; Carr, S.N. Effect of distillers dried grains with solubles and ractopamine (Paylean) on quality and shelf-life of fresh pork and bacon. J. Anim. Sci. 2010, 88, 2751–2766. [Google Scholar] [CrossRef] [PubMed]
- National Live Stock Meat Board. Research Guidelines for Cookery, Sensory Evaluation, and Instrumental Tenderness Measurements of Fresh Meat; American Meat Science Association: Savoy, IL, USA, 1995; ISBN 0-88700-018-5. [Google Scholar]
- NPPC. Procedures to Evaluate Market Hogs, 3rd ed.; Pork Producers Council: Des Moines, IA, USA, 1991. [Google Scholar]
- NPPC. Pork Quality Standards; National Pork Producers Council: Des Moines, IA, USA, 1999. [Google Scholar]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F. QIIME 2: Reproducible, Interactive, Scalable, and Extensible Microbiome Data Science; PeerJ Preprints: Corte Madera, CA, USA, 2018. [Google Scholar]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90–106. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Team, R.C. R: A language and environment for statistical computing; R Foundation for statistical computing: Vienna, Austria, 2019. [Google Scholar]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘Vegan.’ Community Ecology Package Version; R Foundation: Vienna, Austria, 2013. [Google Scholar]
- Anderson, M.J. Distance-based tests for homogeneity of multivariate dispersions. Biometrics 2006, 62, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Martinez Arbizu, P. PairwiseAdonis: Pairwise Multilevel Comparison Using Adonis. R Package Version 00; R Foundation: Vienna, Austria, 2017. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin, Germany, 2016; ISBN 3-319-24277-6. [Google Scholar]
- Chae, B.J.; Choi, S.C.; Kim, Y.G.; Kim, C.H.; Sohn, K.S. Effects of feeding dried food waste on growth and nutrient digestibility in growing-finishing pigs. Asian Australas. J. Anim. Sci. 2000, 13, 1304–1308. [Google Scholar] [CrossRef]
- Kjos, N.P.; Øverland, M.; Bryhni, E.A.; SØrheim, O. Food waste products in diets for growing-finishing pigs: Effect on growth performance, carcass characteristics and meat quality. Acta Agric. Scand. Sect. Anim. Sci. 2000, 50, 193–204. [Google Scholar] [CrossRef]
- Westendorf, M.L.; Dong, Z.C.; Schoknecht, P.A. Recycled cafeteria food waste as a feed for swine: Nutrient content digestibility, growth, and meat quality. J. Anim. Sci. 1998, 76, 2976–2983. [Google Scholar] [CrossRef] [PubMed]
- Choe, J.; Moyo, K.M.; Park, K.; Jeong, J.; Kim, H.; Ryu, Y.; Kim, J.; Kim, J.; Lee, S.; Go, G. Meat quality traits of pigs finished on food waste. Korean J. Food Sci. Anim. Resour. 2017, 37, 690–697. [Google Scholar] [CrossRef]
- Apple, J.; Kegley, E.; Maxwell, C., Jr.; Rakes, L.; Galloway, D.; Wistuba, T. Effects of dietary magnesium and short-duration transportation on stress response, postmortem muscle metabolism, and meat quality of finishing swine. J. Anim. Sci. 2005, 83, 1633–1645. [Google Scholar] [CrossRef]
- Ryu, Y.C.; Kim, B.C. The relationship between muscle fiber characteristics, postmortem metabolic rate, and meat quality of pig longissimus dorsi muscle. Meat Sci. 2005, 71, 351–357. [Google Scholar] [CrossRef]
- Liu, Y.; Kil, D.Y.; Perez-Mendoza, V.G.; Song, M.; Pettigrew, J.E. Supplementation of different fat sources affects growth performance and carcass composition of finishing pigs. J. Anim. Sci. Biotechnol. 2018, 9, 56–63. [Google Scholar] [CrossRef]
- Pettigrew, J.; Moser, R.; Miller, E.; Ullrey, D.; Lewis, A. Swine nutrition. Fat Swine Nutr. 1991, 133–145. [Google Scholar]
- Azain, M. Fat in swine nutrition. Swine Nutr. 2001, 2, 95–105. [Google Scholar]
- Lampe, J.F.; Baas, T.J.; Mabry, J.W. Comparison of grain sources for swine diets and their effect on meat and fat quality traits. J. Anim. Sci. 2006, 84, 1022–1029. [Google Scholar] [CrossRef] [PubMed]
- Allee, G.L.; Baker, D.H.; Leveille, G.A. Influence of level of dietary fat on adipose tissue lipogenesis and enzymatic activity in the pig. J. Anim. Sci. 1971, 33, 1248–1254. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.R.; Knabe, D.A.; Smith, S.B. Depression of lipogenesis in swine adipose tissue by specific dietary fatty acids. J. Anim. Sci. 1996, 74, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Mitchaothai, J.; Yuangklang, C.; Wittayakun, S.; Vasupen, K.; Wongsutthavas, S.; Srenanul, P.; Hovenier, R.; Everts, H.; Beynen, A.C. Effect of dietary fat type on meat quality and fatty acid composition of various tissues in growing-finishing swine. Meat Sci. 2007, 76, 95–101. [Google Scholar] [CrossRef]
- Rezar, V.; Pajk, T.; Marinsek Logar, R.; Jese Janezic, V.; Salobir, K.; Oresnik, A.; Salobir, J. Wheat bran and oat bran effectively reduce oxidative stress induced by high-fat diets in pigs. Ann. Nutr. Metab. 2003, 47, 78–84. [Google Scholar] [CrossRef]
- Tao, L. Oxidation of polyunsaturated fatty acids and its impact on food quality and human health. Adv. Food Technol. Nutr. Sci. Open J. 2015, 1, 135–142. [Google Scholar] [CrossRef]
- Whittington, F.M.; Prescott, N.J.; Wood, J.D.; Enser, M. The effect of dietary linoleic acid on the firmness of backfat in pigs of 85 kg live weight. J. Sci. Food Agric. 1986, 37, 753–761. [Google Scholar] [CrossRef]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 265–270. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Dominianni, C.; Sinha, R.; Goedert, J.J.; Pei, Z.; Yang, L.; Hayes, R.B.; Ahn, J. Sex, body mass index, and dietary fiber intake influence the human gut microbiome. PLoS ONE 2015, 10, e0124599. [Google Scholar] [CrossRef] [Green Version]
- Mathur, R.; Barlow, G.M. Obesity and the microbiome. Expert Rev. Gastroenterol. Hepatol. 2015, 9, 1087–1099. [Google Scholar] [CrossRef] [PubMed]
- Barlow, G.M.; Yu, A.; Mathur, R. Role of the gut microbiome in obesity and diabetes mellitus. Nutr. Clin. Pract. 2015, 30, 787–797. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, Q.; Ma, W.; Tian, F.; Shen, H.; Zhou, M. A combination of quercetin and resveratrol reduces obesity in high-fat diet-fed rats by modulation of gut microbiota. Food Funct. 2017, 8, 4644–4656. [Google Scholar] [CrossRef] [PubMed]
- Lecomte, V.; Kakkoush, N.O.; Maloney, C.A.; Ralpuria, M.; Huinao, K.D.; Mitchell, H.M.; Morris, M.J. Changes in gut microbiota in rats fed a high fat diet correlate with obesity-associated metabolic parameters. PLoS ONE 2015, 10, e0126931. [Google Scholar] [CrossRef] [Green Version]
- Costantini, L.; Molinari, R.; Farinon, B.; Merendino, N. Impact of omega-3 fatty acids on the gut microbiota. Int. J. Mol. Sci. 2017, 18, 2645–2662. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient, g/kg | Control | 100% Food Waste 1 | ||
|---|---|---|---|---|
| Phase 1 | Phase 2 | Phase 3 | ||
| Ground corn | 682.0 | 743.7 | 777.0 | − |
| Soybean meal, 48% crude protein | 270.0 | 210.0 | 180.0 | − |
| Soybean oil | 20.0 | 20.0 | 20.0 | − |
| Limestone | 8.3 | 8.0 | 7.5 | − |
| Dicalcium phosphate | 10.5 | 9.0 | 7.0 | − |
| L-Lysine HCL, 78% Lysine | 1.8 | 2.0 | 1.4 | − |
| DL-Methionine | 0.2 | − | − | − |
| L-Threonine | 0.2 | 0.3 | 0.1 | − |
| Food waste | − | − | − | 998.2 |
| Salt | 4.0 | 4.0 | 4.0 | 1.0 |
| Vitamin-mineral premix 2 | 3.0 | 3.0 | 3.0 | 0.8 |
| Total | 1000.0 | 1000.0 | 1000.0 | 1000.0 |
| Analyzed nutrients, dry matter basis, g/kg | ||||
| Dry matter | 863.4 | 867.3 | 880.2 | 237.2 |
| Crude protein | 212.1 | 173.7 | 158.9 | 257.4 |
| Crude fat | 50.4 | 53.0 | 58.1 | 302.3 |
| Acid detergent fiber | 44.5 | 47.0 | 47.9 | 84.2 |
| Neutral detergent fiber | 124.5 | 107.0 | 112.3 | 113.5 |
| Saturated fatty acid, % of crude fat | ||||
| Myristic (14:0) | 0.08 | 0.07 | 0.06 | 2.48 |
| Pentadecylic (15:0) | 0.02 | 0.02 | 0.02 | 0.40 |
| Palmitic (16:0) | 12.88 | 12.77 | 12.81 | 21.82 |
| Margaric (17:0) | 0.11 | 0.11 | 0.11 | 0.97 |
| Stearic (18:0) | 3.03 | 2.88 | 3.00 | 10.34 |
| Arachidic (20:0) | 0.38 | 0.35 | 0.35 | 0.20 |
| Behenoic (22:0) | 0.25 | 0.25 | 0.26 | 0.10 |
| Total | 16.95 | 16.63 | 16.81 | 36.36 |
| Unsaturated fatty acid, % of crude fat | ||||
| Myristoleic (14:1) | 0.00 | 0.00 | 0.00 | 0.45 |
| Palmitoleic (16:1) | 0.16 | 0.13 | 0.11 | 3.45 |
| Oleic (18:1) | 23.01 | 23.58 | 23.71 | 34.48 |
| Vaccenic (18:1) | 1.22 | 1.17 | 1.20 | 2.19 |
| Linoleic (18:2) | 52.43 | 52.85 | 52.14 | 10.95 |
| Linolenic (18:3) | 4.31 | 4.01 | 4.09 | 1.36 |
| Gondoic (20:1) | 0.35 | 0.33 | 0.33 | 0.44 |
| Eicosadienoic (20:2) | 0.02 | 0.02 | 0.03 | 0.18 |
| Arachidonic (20:4) | 0.00 | 0.00 | 0.00 | 0.34 |
| Eicosapentaenoic (EPA, 20:5) | 0.00 | 0.00 | 0.00 | 0.34 |
| Docosaherxaenoic (DHA, 22:6) | 0.00 | 0.00 | 0.00 | 0.28 |
| Monounsaturated fatty acids | 24.77 | 25.24 | 25.39 | 41.10 |
| Polyunsaturated fatty acids | 56.76 | 56.89 | 56.25 | 13.52 |
| Total fatty acid, % of crude fat | 81.53 | 82.12 | 81.64 | 54.61 |
| Item 1 | Control | Food Waste | SEM | p-Value |
|---|---|---|---|---|
| Ending live weight, kg | 105.79 | 97.86 | 3.36 | 0.050 |
| HCW, kg | 84.11 | 76.98 | 2.91 | 0.028 |
| Carcass yield, % | 79.57 | 78.70 | 0.93 | 0.320 |
| Loin eye area, cm2 | 49.25 | 47.45 | 3.079 | 0.444 |
| 10th rib back-fat, cm | 2.34 | 1.76 | 0.244 | 0.152 |
| Estimated carcass lean, % | 52.65 | 55.57 | 1.344 | 0.186 |
| Item 1 | Control | Food Waste | SEM | p-Value |
|---|---|---|---|---|
| Moisture, % | 74.28 | 74.53 | 0.203 | 0.412 |
| Shear force, kg | 3.16 | 3.23 | 0.232 | 0.757 |
| Cook loss, % | 26.28 | 28.16 | 4.287 | 0.602 |
| pH | 5.45 | 5.42 | 0.082 | 0.444 |
| Objective color | ||||
| L* | 56.85 | 56.94 | 1.26 | 0.962 |
| a* | 8.194 | 7.938 | 0.685 | 0.631 |
| b* | 15.609 | 15.401 | 0.999 | 0.786 |
| Subjective evaluations | ||||
| Color | 1.98 | 2.12 | 0.241 | 0.581 |
| Marbling | 1.671 | 1.671 | 0.346 | 1.000 |
| Firmness | 2.857 | 2.429 | 0.175 | 0.087 |
| Drip loss, % | 3.367 | 4.259 | 0.678 | 0.394 |
| Item 1 | Control | Food Waste | SEM | p-Value |
|---|---|---|---|---|
| Half carcass chilled weight, kg | 40.68 | 37.43 | 1.38 | 0.054 |
| Lean cutting yield 2, % | 63.85 | 65.33 | 3.80 | 0.329 |
| Boneless lean cutting yield 3, % | 48.72 | 50.25 | 2.19 | 0.271 |
| Carcass cutting yield 4, % | 72.78 | 74.58 | 3.94 | 0.256 |
| Boneless carcass cutting yield 5, % | 57.65 | 58.50 | 2.06 | 0.203 |
| Item 1 | Control | Food Waste | SEM | p-Value |
|---|---|---|---|---|
| Moisture, % | 10.41 | 11.24 | 1.14 | 0.54 |
| Crude fat, % | 86.00 | 85.19 | 1.55 | 0.60 |
| Saturated fatty acids, % of crude fat | ||||
| Myristic (14:0) | 1.19 | 1.216 | 0.03 | 0.54 |
| Pentadecylic (15:0) | 0.05 | 0.126 | 0.01 | <0.01 |
| Palmitic (16:0) | 22.40 | 19.482 | 0.68 | <0.01 |
| Margaric (17:0) | 0.30 | 0.555 | 0.05 | <0.01 |
| Stearic (18:0) | 11.36 | 10.372 | 1.06 | 0.16 |
| Arachidic (20:0) | 0.23 | 0.182 | 0.03 | <0.01 |
| Behenoic (22:0) | 0.040 | 0.041 | 0.00 | 0.62 |
| Total | 35.56 | 31.96 | 1.66 | 0.02 |
| Unsaturated fatty acids, % of crude fat | ||||
| Myristoleic (14:1) | 0.02 | 0.07 | 0.01 | <0.01 |
| Palmitoleic (16:1) | 1.76 | 2.24 | 0.17 | <0.01 |
| Oleic (18:1) | 35.21 | 39.64 | 0.68 | <0.01 |
| Vaccenic (18:1) | 2.36 | 2.842 | 0.12 | <0.01 |
| Linoleic (18:2) | 19.51 | 15.08 | 0.64 | <0.01 |
| Linolenic (18:3) | 1.52 | 1.21 | 0.09 | <0.01 |
| Gondoic (20:1) | 0.68 | 0.83 | 0.14 | <0.01 |
| Eicosadienoic (20:2) | 0.77 | 0.63 | 0.04 | 0.02 |
| Arachidonic (20:4) | 0.25 | 0.26 | 0.02 | 0.73 |
| Eicosapentaenoic (EPA, 20:5) | 0.01 | 0.09 | 0.04 | 0.02 |
| Docosaherxaenoic (DHA, 22:6) | 0.01 | 0.17 | 0.06 | <0.01 |
| Monounsaturated fatty acids | 40.04 | 45.64 | 0.83 | <0.01 |
| Polyunsaturated fatty acids | 22.36 | 17.84 | 0.71 | <0.01 |
| Total fatty acids, % of crude fat | 62.52 | 63.61 | 1.34 | 0.33 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jinno, C.; Saelao, P.; Maga, E.A.; King, A.; Morash, D.; Zicari, S.; Yang, X.; Liu, Y. Enzymatically Digested Food Waste Altered Fecal Microbiota But Not Meat Quality and Carcass Characteristics of Growing-Finishing Pigs. Animals 2019, 9, 970. https://doi.org/10.3390/ani9110970
Jinno C, Saelao P, Maga EA, King A, Morash D, Zicari S, Yang X, Liu Y. Enzymatically Digested Food Waste Altered Fecal Microbiota But Not Meat Quality and Carcass Characteristics of Growing-Finishing Pigs. Animals. 2019; 9(11):970. https://doi.org/10.3390/ani9110970
Chicago/Turabian StyleJinno, Cynthia, Perot Saelao, Elizabeth A. Maga, Annie King, Dan Morash, Steve Zicari, Xiang Yang, and Yanhong Liu. 2019. "Enzymatically Digested Food Waste Altered Fecal Microbiota But Not Meat Quality and Carcass Characteristics of Growing-Finishing Pigs" Animals 9, no. 11: 970. https://doi.org/10.3390/ani9110970