Uptake of Manganese from the Manganese-Lysine Complex in Primary Chicken Intestinal Epithelial Cells

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Antibodies
2.2. Intestinal Epithelial Cells Isolation and Culture
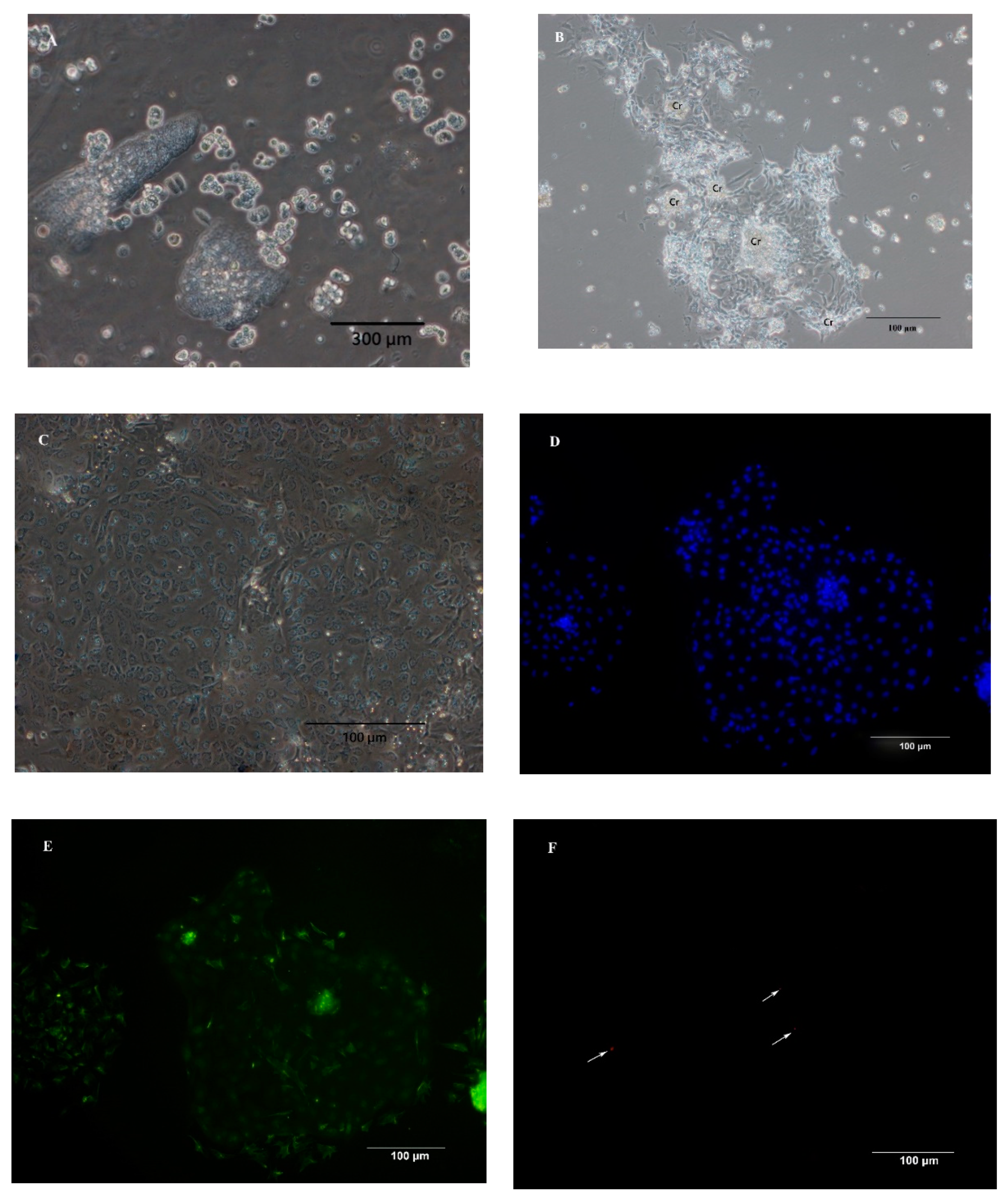
2.3. Characterization of IEC Primary Culture
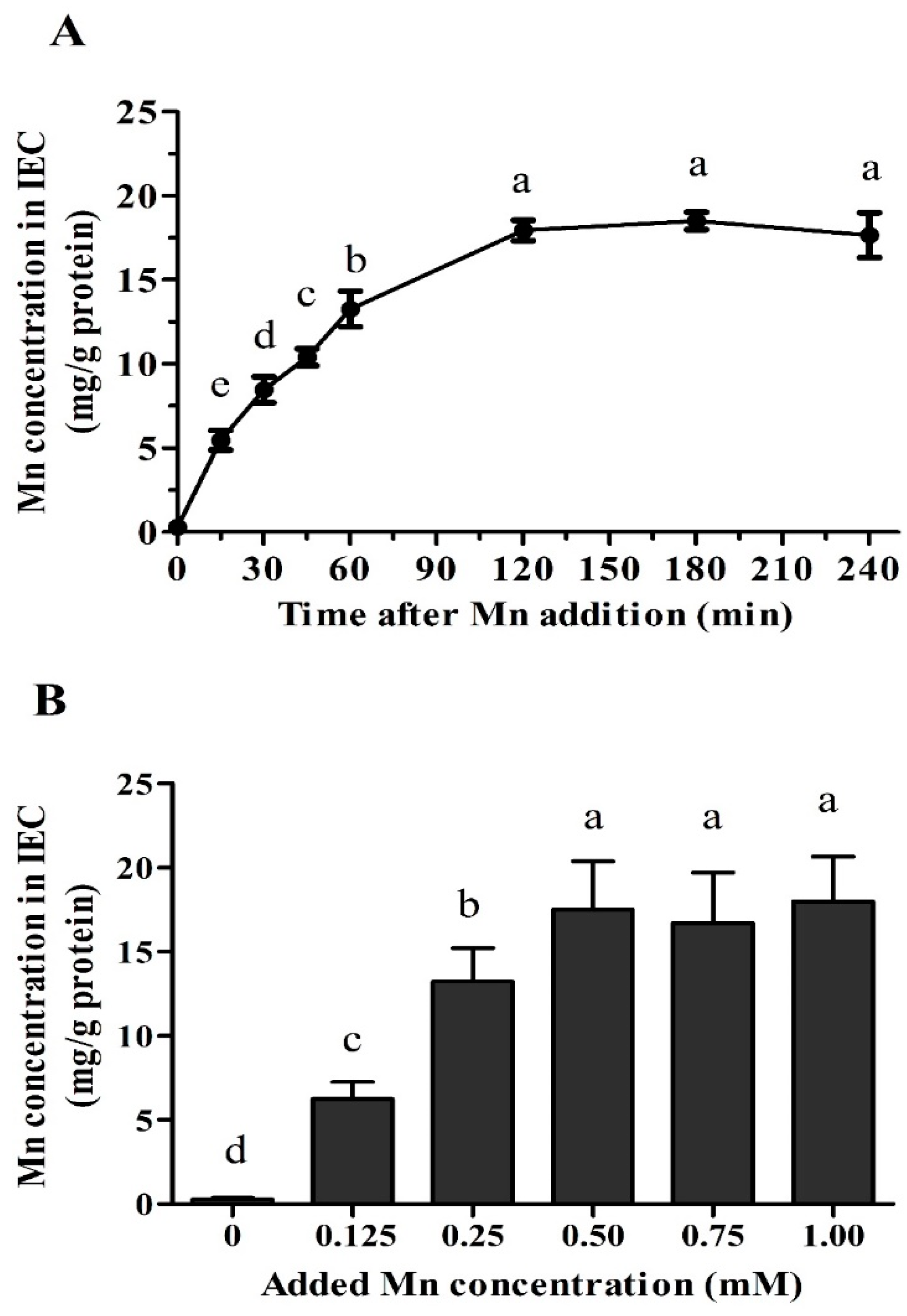
2.4. Time- and Dose-Dependent Mn Uptake
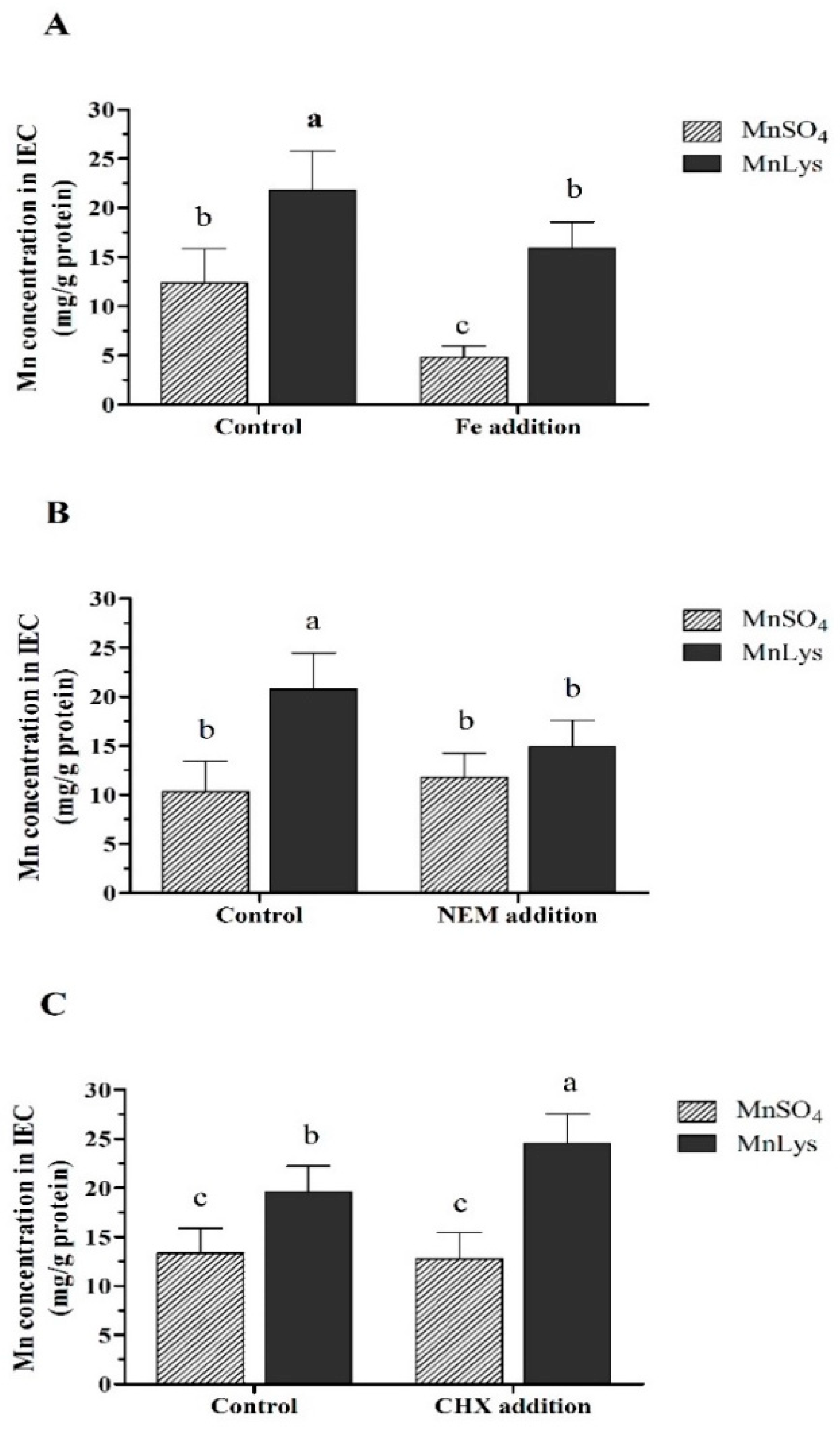
2.5. Uptake of Mn from Different Mn Sources
2.6. Mn and Protein Concentrations Determination
2.7. Quantitative Real-Time RT-PCR
2.8. Statistical Analysis
3. Results
3.1. Isolation of IEC and Characterization of Primary IEC Cultures
3.2. Time- and Dose-Dependent Mn Uptake
3.3. Uptake of Mn in Different Mn Sources
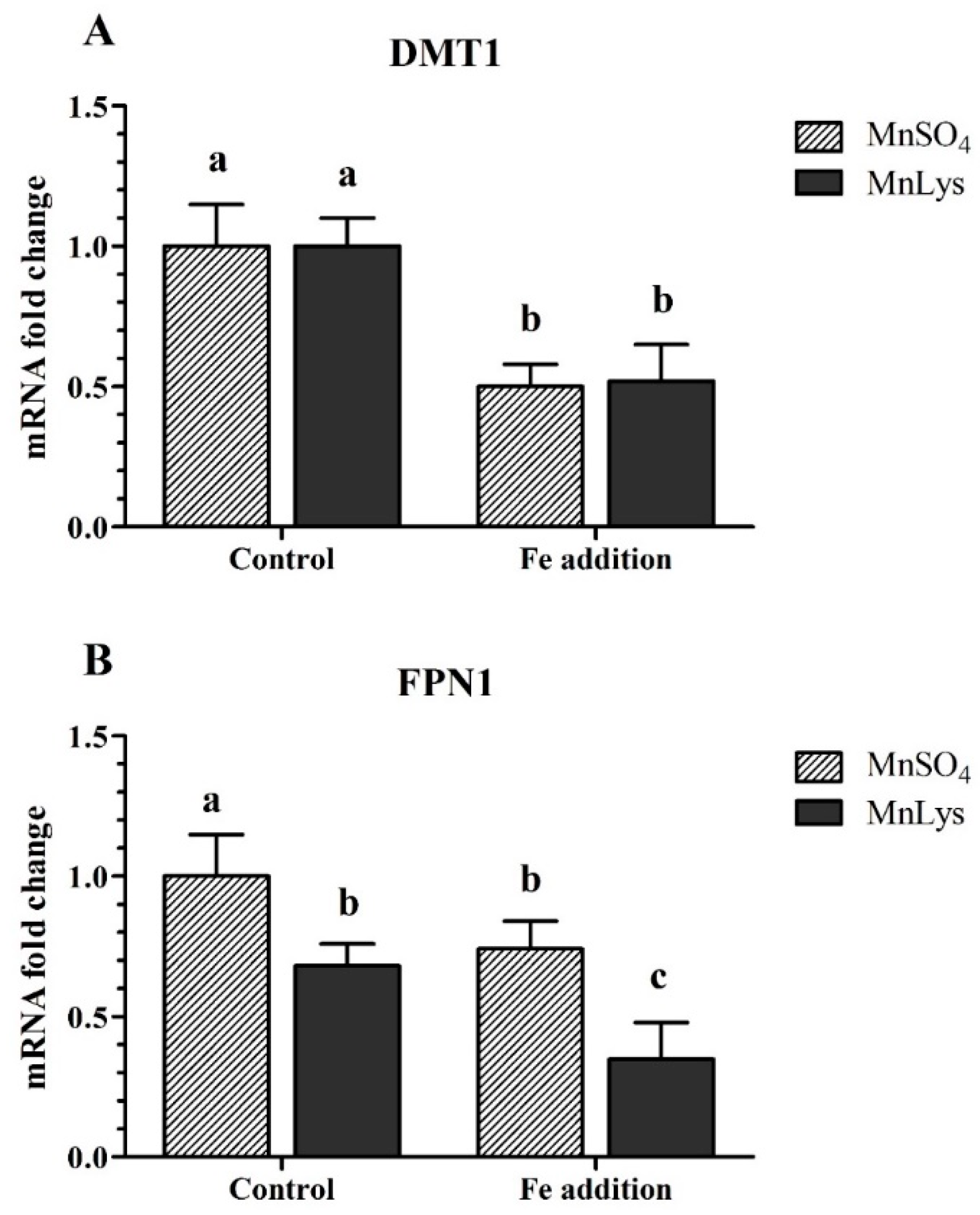
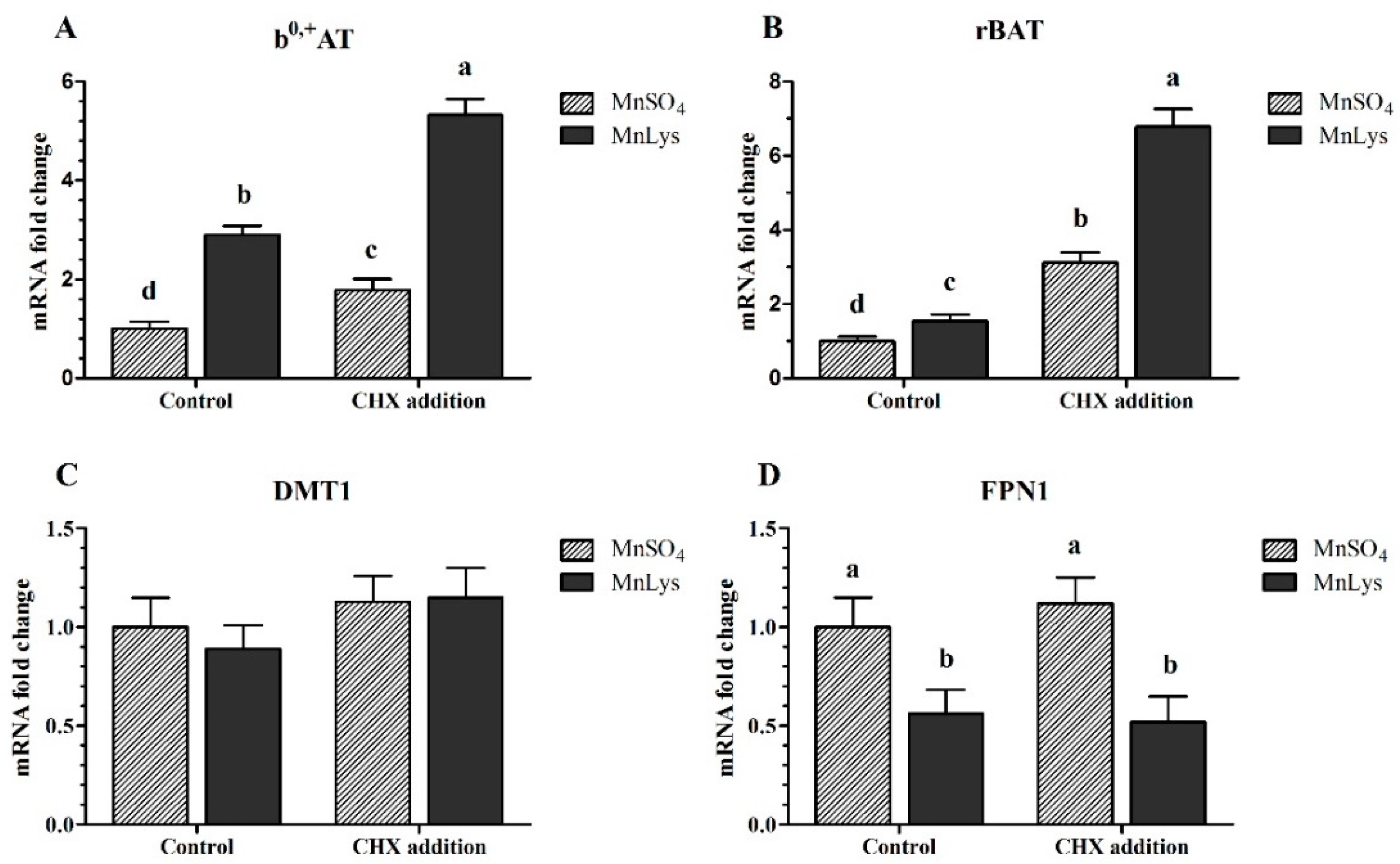
3.4. Transporters Expression
4. Discussion
Author Contributions
Acknowledgment
Conflicts of Interest
References
- Li, L.; Yang, X. The essential element manganese, oxidative stress, and metabolic diseases: Links and interactions. Oxid. Med. Cell. Longev. 2018, 2018, 7580707. [Google Scholar] [CrossRef]
- Henry, P.R. Manganese bioavailability. In Bioavailability of Nutrients for Animals: Amino Acids, Minerals, and Vitamins; Ammerman, C.B., Baker, D.H., Lewis, A.J., Eds.; Academic Press: San Diego, CA, USA, 1995; pp. 239–256. [Google Scholar] [CrossRef]
- Halpin, K.M.; Baker, D.H. Manganese utilization in the chick: Effects of corn, soybean, fish meal, wheat bran, and rice bran on tissue uptake of manganese. Poult. Sci. 1986, 65, 995–1003. [Google Scholar] [CrossRef]
- Bai, S.P.; Lu, L.; Luo, X.G.; Liu, B. Kinetics of manganese absorption in ligated small intestinal segments of broilers. Poult. Sci. 2008, 87, 2596–2604. [Google Scholar] [CrossRef]
- Bai, S.P.; Lu, L.; Wang, R.L.; Xi, L.; Zhang, L.Y.; Luo, X.G. Manganese source affects manganese transport and gene expression of divalent metal transporter 1 in the small intestine of broilers. Br. J. Nutr. 2012, 108, 267–276. [Google Scholar] [CrossRef]
- Li, X.; Xie, J.; Lu, L.; Zhang, L.; Zou, Y.; Wang, Q.; Luo, X.G. Kinetics of manganese transport and gene expressions of manganese transport carriers in Caco-2 cell monolayers. Biometals 2013, 26, 941–953. [Google Scholar] [CrossRef]
- Baker, D.H.; Halpin, K.M. Efficacy of a manganese-protein chelate compared with that of manganese sulfate for chicks. Poult. Sci. 1987, 66, 1561–1563. [Google Scholar] [CrossRef]
- Scheideler, S.E. Interaction of dietary calcium, manganese, and manganese source (Mn oxide or Mn methionine complex) on chick performance and manganese utilization. Biol. Trace Elem. Res. 1991, 29, 217–228. [Google Scholar] [CrossRef]
- Ochiai, H.; Moriyama, J.; Kanemaki, N.; Sato, R.; Onda, K. Analysis of cationic amino acid transport activity in canine lens epithelial cells. Exp. Anim. 2013, 62, 311–317. [Google Scholar] [CrossRef]
- De Santa Barbara, P.; Van Den Brink, G.R.; Roberts, D.J. Development and differentiation of the intestinal epithelium. Cell. Mol. Life Sci. 2003, 60, 1322–1332. [Google Scholar] [CrossRef]
- Maldonado-Contreras, A.L.; McCormick, B.A. Intestinal epithelial cells and their role in innate mucosal immunity. Cell Tissue Res. 2011, 343, 5–12. [Google Scholar] [CrossRef]
- Dimier, I.H.; Bout, D.T. Interferon-gamma-activated primary enterocytes inhibit Toxoplasma gondii replication: A role for intracellular iron. Immunology 1998, 94, 488–495. [Google Scholar] [CrossRef]
- Ní Shúilleabháin, S.; Mothersill, C.; Sheehan, D.; O’Brien, N.M.; O’Halloran, J.; Van Pelt, F.N.; Kilemade, M.; Davoren, M. Cellular responses in primary epidermal cultures from rainbow trout exposed to zinc chloride. Ecotoxicol. Environ. Saf. 2006, 65, 332–341. [Google Scholar] [CrossRef]
- Wang, Y.; Yan, X.; Fu, L. Effect of selenium nanoparticles with different sizes in primary cultured intestinal epithelial cells of crucian carp, Carassius auratus gibelio. Int. J. Nanomed. 2013, 8, 4007–4013. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Li, Y.; Buckett, P.D.; Bohlke, M.; Thompson, K.J.; Takahashi, M.; Maher, T.J.; Wessling-Resnick, M. Iron-responsive olfactory uptake of manganese improves motor function deficits associated with iron deficiency. PLoS ONE 2012, 7, e33533. [Google Scholar] [CrossRef]
- Yin, Z.; Jiang, H.; Lee, E.S.; Ni, M.; Erikson, K.M.; Milatovic, D.; Bowman, A.B.; Aschner, M. Ferroportin is a manganese-responsive protein that decreases manganese cytotoxicity and accumulation. J. Neurochem. 2010, 112, 1190–1198. [Google Scholar] [CrossRef] [Green Version]
- Madejczyk, M.S.; Ballatori, N. The iron transporter ferroportin can also function as a manganese exporter. Biochim. Biophys. Acta BBA Biomembr. 2012, 1818, 651–657. [Google Scholar] [CrossRef] [Green Version]
- Ashmead, H.D. Comparative intestinal absorption and subsequent metabolism of metal amino acid chelates and inorganic metal salts. In The Roles of Amino Acid Chelates in Animal Nutrition; Ashmead, H.D., Ed.; Noyes Publications: Park Ridge, NJ, USA, 1993; pp. 47–74. [Google Scholar]
- Li, S.; Luo, X.G.; Liu, B.; Crenshaw, T.D.; Kuang, X.; Shao, G.; Yu, S. Use of chemical characteristics to predict the relative bioavailability of supplemental organic manganese sources for broilers. J. Anim. Sci. 2004, 82, 2352–2363. [Google Scholar] [CrossRef]
- Lowe, J.A.; Wiseman, J.; Cole, D.J. Zinc source influences zinc retention in hair and hair growth in the dog. J. Nutr. 1994, 124, 2575S–2576S. [Google Scholar] [CrossRef]
- Glover, C.N.; Wood, C.M. Histidine absorption across apical surfaces of freshwater rainbow trout intestine: Mechanistic characterization and the influence of copper. J. Membr. Biol. 2008, 221, 87–95. [Google Scholar] [CrossRef]
- Grandjean, P.; Landrigan, P.J. Neurobehavioural effects of developmental toxicity. Lancet Neurol. 2014, 13, 330–338. [Google Scholar] [CrossRef] [Green Version]
- Thwaites, D.T.; Markovich, D.; Murer, H.; Simmons, N.L. Na+-independent lysine transport in human intestinal Caco-2 cells. J. Membr. Biol. 1996, 151, 215–224. [Google Scholar] [CrossRef]
- Torras-Llort, M.; Ferrer, R.; Soriano-Garcia, J.F.; Moreto, M. L-lysine transport in chicken jejunal brush border membrane vesicles. J. Membr. Biol. 1996, 152, 183–193. [Google Scholar] [CrossRef]
- Torras-Llort, M.; Soriano-Garcia, J.F.; Ferrer, R.; Moreto, M. Effect of a lysine-enriched diet on L-lysine transport by the brush-border membrane of the chicken jejunum. Am. J. Physiol. 1998, 274, R69–R75. [Google Scholar] [CrossRef]
- Deves, R.; Boyd, C.A. Transporters for cationic amino acids in animal cells: Discovery, structure, and function. Physiol. Rev. 1998, 78, 487–545. [Google Scholar] [CrossRef]
- Peluffo, R.D. L-Arginine currents in rat cardiac ventricular myocytes. J. Physiol. 2007, 580, 925–936. [Google Scholar] [CrossRef]
- Nishino, H.; Christopher, C.W.; Schiller, R.M.; Gammon, M.T.; Ullrey, D.; Isselbacher, K.J. Sodium-dependent amino acid transport by cultured hamster cells: Membrane vesicles retain transport changes due to glucose starvation and cycloheximide. Proc. Natl. Acad. Sci. USA 1987, 75, 5048–5051. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC382979/ (accessed on 25 March 2019). [CrossRef]
- Evans, G.S.; Flint, N.; Somers, A.S.; Eyden, B.; Potten, C.S. The development of a method for the preparation of rat intestinal epithelial cell primary cultures. J. Cell Sci. 1992, 101, 219–231. [Google Scholar]
- Satsu, H.; Hyun, J.S.; Shin, H.S.; Shimizu, M. Cycloheximide treatment induces the uptake of neutral and dibasic amino acids via the activation of system b0,+ in human intestinal Caco-2 cells. J. Nutr. Sci. Vitaminol. 2009, 55, 44–51. [Google Scholar] [CrossRef]
- Bar Shira, E.; Friedman, A. Innate immune functions of avian intestinal epithelial cells: Response to bacterial stimuli and localization of responding cells in the developing avian digestive tract. PLoS ONE 2018, 13, e0200393. [Google Scholar] [CrossRef]
- Qin, B.; Dawson, H.D.; Schoene, N.W.; Polansky, M.M.; Anderson, R.A. Cinnamon polyphenols regulate multiple metabolic pathways involved in insulin signaling and intestinal lipoprotein metabolism of small intestinal enterocytes. Nutrition 2012, 28, 1172–1179. [Google Scholar] [CrossRef]
- Zhang, H.; Gilbert, E.R.; Zhang, K.; Ding, X.; Luo, Y.; Wang, J.; Zeng, Q.; Bai, S. Uptake of manganese from manganese-lysine complex in the primary rat intestinal epithelial cells. J. Anim. Physiol. Anim. Nutr. 2017, 101, 147–158. [Google Scholar] [CrossRef]
- Bai, S.; Huang, L.; Luo, Y.; Wang, L.; Ding, X.; Wang, J.; Zeng, Q.; Zhang, K. Dietary manganese supplementation influences the expression of transporters involved in iron metabolism in chickens. Biol. Trace Elem. Res. 2014, 160, 352–360. [Google Scholar] [CrossRef]
- Uni, Z.; Geyra, A.; Ben-Hur, H.; Sklan, D. Small intestinal development in the young chick: Crypt formation and enterocyte proliferation and migration. Br. Poult. Sci. 2000, 41, 544–551. [Google Scholar] [CrossRef]
- Kaiser, A.; Willer, T.; Steinberg, P.; Rautenschlein, S. Establishment of an in vitro intestinal cell culture model of avian origin. Avian Dis. 2017, 61, 229–236. [Google Scholar] [CrossRef]
- Nelson, W.J. Development of maintenance of epithelial polarity: A role for the submembranous cytoskeleton. In Functional Epithelial Cells in Culture; Matlin, K.S., Valentich, J.D., Eds.; Alan, R. Liss Inc.: New York, NY, USA, 1989; pp. 3–42. [Google Scholar]
- Deves, R.; Angelo, S.; Chavez, P. N-ethylmaleimide discriminates between two lysine transport systems in human erythrocytes. J. Physiol. 1993, 468, 753–766. [Google Scholar] [CrossRef]
- Yokomichi, T.; Morimoto, K.; Oshima, N.; Yamada, Y.; Fu, L.; Taketani, S.; Ando, M.; Kataoka, T. Ursolic acid inhibits Na+/K+-ATPase activity and presents TNF-α-induced gene expression by blocking amino acid transport and cellular protein synthesis. Biomolecules 2011, 1, 32–47. [Google Scholar] [CrossRef]
- Kedinger, M.; Haffen, K.; Simon-Assmann, P. Intestinal tissue and cell cultures. Differentiation 1987, 36, 71–85. [Google Scholar] [CrossRef]
- Erikson, K.M.; Aschner, M. Increased manganese uptake by primary astrocyte cultures with altered iron status is mediated primarily by divalent metal transporter. Neurotoxicology 2006, 27, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Li, S.F.; Luo, X.G.; Lu, L.; Crenshaw, T.D.; Bu, Y.Q.; Liu, B.; Kuang, X.; Shao, G.Z.; Yu, S.X. Bioavailability of organic manganese sources in broilers fed high dietary calcium. Anim. Feed Sci. Technol. 2005, 123–124, 703–715. [Google Scholar] [CrossRef]
- Ji, F.; Luo, X.G.; Lu, L.; Liu, B.; Yu, S.X. Effects of manganese source and calcium on manganese uptake by in vitro everted gut sacs of broilers’ intestinal segments. Poult. Sci. 2006, 85, 1217–1225. [Google Scholar] [CrossRef]
- Turi, J.L.; Yang, F.; Garrick, M.D.; Piantadosi, C.A.; Ghio, A.J. The iron cycle and oxidative stress in the lung. Free Radic. Biol. Med. 2004, 36, 850–857. [Google Scholar] [CrossRef]
- Chua, A.C.; Morgan, E.H. Manganese metabolism is impaired in the Belgrade laboratory rat. J. Comp. Physiol. B 1997, 167, 361–369. [Google Scholar] [CrossRef]
- Fleming, R.E.; Migas, M.C.; Zhou, X.; Jiang, J.; Britton, R.S.; Brunt, E.M.; Tomatsu, S.; Waheed, A.; Bacon, B.R.; Sly, W.S. Mechanism of increased iron absorption in murine model of hereditary hemochromatosis: Increased duodenal expression of the iron transporter DMT1. Proc. Natl. Acad. Sci. USA 1999, 96, 3143–3148. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC15909/ (accessed on 25 March 2019). [CrossRef] [Green Version]
- Gunshin, H.; Allerson, C.R.; Polycarpou-Schwarz, M.; Rofts, A.; Rogers, J.T.; Kishi, F.; Hentze, M.W.; Rouault, T.A.; Andrews, N.C.; Hediger, M.A. Iron-dependent regulation of the divalent metal ion transporter. FEBS Lett. 2001, 509, 309–316. Available online: https://www.ncbi.nlm.nih.gov/pubmed/11741608 (accessed on 25 March 2019). [CrossRef] [Green Version]
- Hubert, N.; Hentze, M.W. Previously uncharacterized isoforms of divalent metal transporter (DMT)-1: Implications for regulation and cellular function. Proc. Natl. Acad. Sci. USA 2002, 99, 12345–12350. [Google Scholar] [CrossRef] [Green Version]
- Davis, C.D.; Wolf, T.L.; Greger, J.L. Varying levels of manganese and iron affect absorption and gut endogenous losses of manganese by rats. J. Nutr. 1992, 122, 1300–1308. [Google Scholar] [CrossRef]
- Heilig, E.A.; Thompson, K.; Molina, R.M.; Ivanov, A.R.; Brain, J.D.; Wessling-Resnick, M. Manganese and iron transport across pulmonary epithelium. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 290, 1247–1259. [Google Scholar] [CrossRef]
- Troadec, M.B.; Ward, D.M.; Lo, E.; Kaplan, J.; Domenico, I. Induction of FPN1 transcription by MTF-1 reveals a role for ferroportin in transition metal efflux. Blood 2010, 116, 4657–4664. [Google Scholar] [CrossRef] [Green Version]
- Ashmead, H.D. Amino Acid Chelation in Human and Animal Nutrition; CRC Press: Boca Raton, FL, USA, 2012; pp. 45–67. [Google Scholar]
- Ashmead, H. Tissue transportation of organic trace minerals. J. Appl. Nutr. 1970, 22, 42–51. [Google Scholar]
- Fang, S.M.; Burton, S.A.; Petersen, R.V. Bioavailability of zinc: Effect of amino acid chelation. In Chelated Mineral Nutrition in Plants, Animals and Man; Ashmead, H.D., Ed.; Thomas: Springfield, IL, USA, 1987; pp. 137–151. [Google Scholar]
- Cannon, V.T.; Kalups, R.K.; Barfuss, D.W. Amino acid transporters involved in luminal transport of mercuric conjugates of cysteine in rabbit proximal tubule. J. Pharmacol. Exp. Ther. 2001, 298, 780–789. Available online: https://www.ncbi.nlm.nih.gov/pubmed/11454942 (accessed on 25 March 2019).
- Pan, M.; Malandro, M.; Stevens, B.R. Regulation of system y+ arginine transport capacity in differentiating human intestinal Caco-2 cells. Am. J. Physiol. Gastrointest. Liver Physiol. 1995, 268, G578–G585. [Google Scholar] [CrossRef]
- Hatzoglou, M.; Fernandez, J.; Yaman, I.; Closs, E. Regulation of cationic amino acid transport: The story of the CAT-1 transporter. Annu. Rev. Nutr. 2004, 24, 377–399. [Google Scholar] [CrossRef]
- Gao, S.; Yin, T.; Xu, B.; Ma, Y.; Hu, M. Amino acid facilitates absorption of copper in the Caco-2 cell culture model. Life Sci. 2014, 109, 50–56. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′-3′) | GeneBank ID |
|---|---|---|---|
| DMT1 | F | CATGTACTTCGTGGTGGCCT | EF635923 |
| R | GATCAGACACAGCCACGTCA | ||
| FPN1 | F | GATGCATTCTGAACAACCAAGGA | GI 61098365 |
| R | GGAGACTGGGTGGACAAGAACTC | ||
| CAT1 | F | CAAGAGGAAAACTCCAGTAATTGCA | XM_417116 |
| R | AAGTCGAAGAGGAAGGCCATAA | ||
| CAT2 | F | TGCTCGCGTTCCCAAGA | XM_420685 |
| R | GGCCCACAGTTCACCAACAG | ||
| b0,+AT | F | CAGTAGTGAATTCTCTGAGTGTGAAGCT | XM_414130 |
| R | GCAATGATTGCCACAACTACCA | ||
| rBAT | F | CCCGCCGTTCAACAAGAG | XM_426125 |
| R | AATTAAATCCATCGACTCCTTTGC | ||
| β-actin | F | FGAGAAATTGTGCGTGACATCA | L08165 |
| R | CCTGAACCTCTCATTGCCA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, S.; Zhang, K.; Ding, X.; Wang, J.; Zeng, Q.; Peng, H.; Bai, J.; Xuan, Y.; Su, Z.; Wu, B. Uptake of Manganese from the Manganese-Lysine Complex in Primary Chicken Intestinal Epithelial Cells. Animals 2019, 9, 559. https://doi.org/10.3390/ani9080559
Bai S, Zhang K, Ding X, Wang J, Zeng Q, Peng H, Bai J, Xuan Y, Su Z, Wu B. Uptake of Manganese from the Manganese-Lysine Complex in Primary Chicken Intestinal Epithelial Cells. Animals. 2019; 9(8):559. https://doi.org/10.3390/ani9080559
Chicago/Turabian StyleBai, Shiping, Keying Zhang, Xuemei Ding, Jianping Wang, Qiufeng Zeng, Huanwei Peng, Jie Bai, Yue Xuan, Zuowei Su, and Bin Wu. 2019. "Uptake of Manganese from the Manganese-Lysine Complex in Primary Chicken Intestinal Epithelial Cells" Animals 9, no. 8: 559. https://doi.org/10.3390/ani9080559