
Evaluating the Age-Dependent Potential for Protein Deposition in Naked Neck Meat Type Chicken

Abstract
:Simple Summary
Abstract
1. Introduction
2. Experimental Section
2.1. Stock and Husbandry
2.2. Feeding and Sampling

{kind=link}
{kind=link}
| Ingredients | Diets | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Starter (Days 10–20) | Grower (Days 25–35) | |||||||||
| Diet 1 | N1 | N2 | N3 | N4 | N5 | N1 | N2 | N3 | N4 | N5 |
| Maize | 6.62 | 10.59 | 14.56 | 18.45 | 22.50 | 5.95 | 9.70 | 13.67 | 18.08 | 22.05 |
| Wheat | 5.09 | 8.14 | 11.19 | 14.19 | 17.30 | 4.58 | 7.46 | 10.51 | 13.90 | 16.95 |
| Soy protein concentrate | 10.15 | 16.23 | 22.32 | 28.29 | 34.50 | 9.13 | 14.88 | 20.96 | 27.72 | 33.81 |
| Fish meal | 1.91 | 3.06 | 4.21 | 5.33 | 6.50 | 1.72 | 2.80 | 3.95 | 5.22 | 6.37 |
| Wheat gluten | 1.76 | 2.82 | 3.88 | 4.92 | 6.00 | 1.59 | 2.59 | 3.65 | 4.82 | 5.88 |
| Soybean oil | 3.09 | 4.94 | 6.79 | 8.61 | 10.50 | 3.24 | 5.28 | 7.44 | 9.84 | 12.00 |
| Cellulose | 1.80 | 1.35 | 0.90 | 0.46 | - | 1.82 | 1.40 | 0.95 | 0.45 | - |
| Wheat starch | 65.54 | 49.17 | 32.78 | 16.72 | - | 68.42 | 52.70 | 35.99 | 17.45 | 0.78 |
| Premix 2 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| DCP | 2.50 | 2.08 | 1.65 | 1.25 | 0.80 | 2.30 | 1.89 | 1.45 | 0.97 | 0.55 |
| CaCO3 | 0.30 | 0.40 | 0.52 | 0.60 | 0.74 | 0.06 | 0.15 | 0.28 | 0.41 | 0.50 |
| NaCl | 0.24 | 0.22 | 0.20 | 0.18 | 0.16 | 0.19 | 0.15 | 0.15 | 0.12 | 0.10 |
| Nutrients | Diets | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Starter (Days 10–20) | Grower (Days 25–35) | |||||||||
| Diet | N1 | N2 | N3 | N4 | N5 | N1 | N2 | N3 | N4 | N5 |
| Crude protein | 10.85 | 17.44 | 24.10 | 30.70 | 37.63 | 9.75 | 15.96 | 22.59 | 30.05 | 36.83 |
| Ether extract | 3.94 | 6.26 | 8.62 | 10.96 | 13.41 | 4.04 | 6.55 | 9.24 | 12.25 | 14.99 |
| Crude fiber | 2.35 | 2.36 | 2.37 | 2.39 | 2.40 | 2.30 | 2.31 | 2.33 | 2.34 | 2.35 |
| Crude ash | 5.43 | 5.86 | 6.32 | 6.75 | 7.24 | 4.78 | 5.14 | 5.62 | 6.11 | 6.55 |
| N-free extract | 70.69 | 60.59 | 50.13 | 40.87 | 30.40 | 72.48 | 62.95 | 52.67 | 41.18 | 30.75 |
| Starch | 65.02 | 55.65 | 46.17 | 36.78 | 26.89 | 70.35 | 60.04 | 49.75 | 37.58 | 27.16 |
| Total sugars | 1.64 | 1.64 | 1.64 | 1.63 | 1.63 | 1.74 | 1.70 | 1.69 | 1.64 | 1.61 |
| AMEn1 (MJ/kg) | 14.73 | 15.5 | 15.33 | 15.16 | 15.37 | 15.02 | 15.21 | 15.43 | 15.47 | 15.59 |
| Amino Acids | Amino Acid Composition (g/100g Crude Protein) | Amino Acid Ratio (Lys = 100) | ||||||||
| Lys | 5.09 | 100 | ||||||||
| Met | 1.44 | 28 | ||||||||
| Cys | 1.46 | 29 | ||||||||
| Thr | 3.63 | 71 | ||||||||
| Trp | 0.96 | 19 | ||||||||
| Arg | 6.26 | 123 | ||||||||
| Ile | 4.03 | 79 | ||||||||
| Leu | 7.50 | 147 | ||||||||
| Val | 4.24 | 83 | ||||||||
| Phe | 4.71 | 92 | ||||||||
2.3. Laboratory Analysis
2.4. Model Parameter Assessment and Statistics
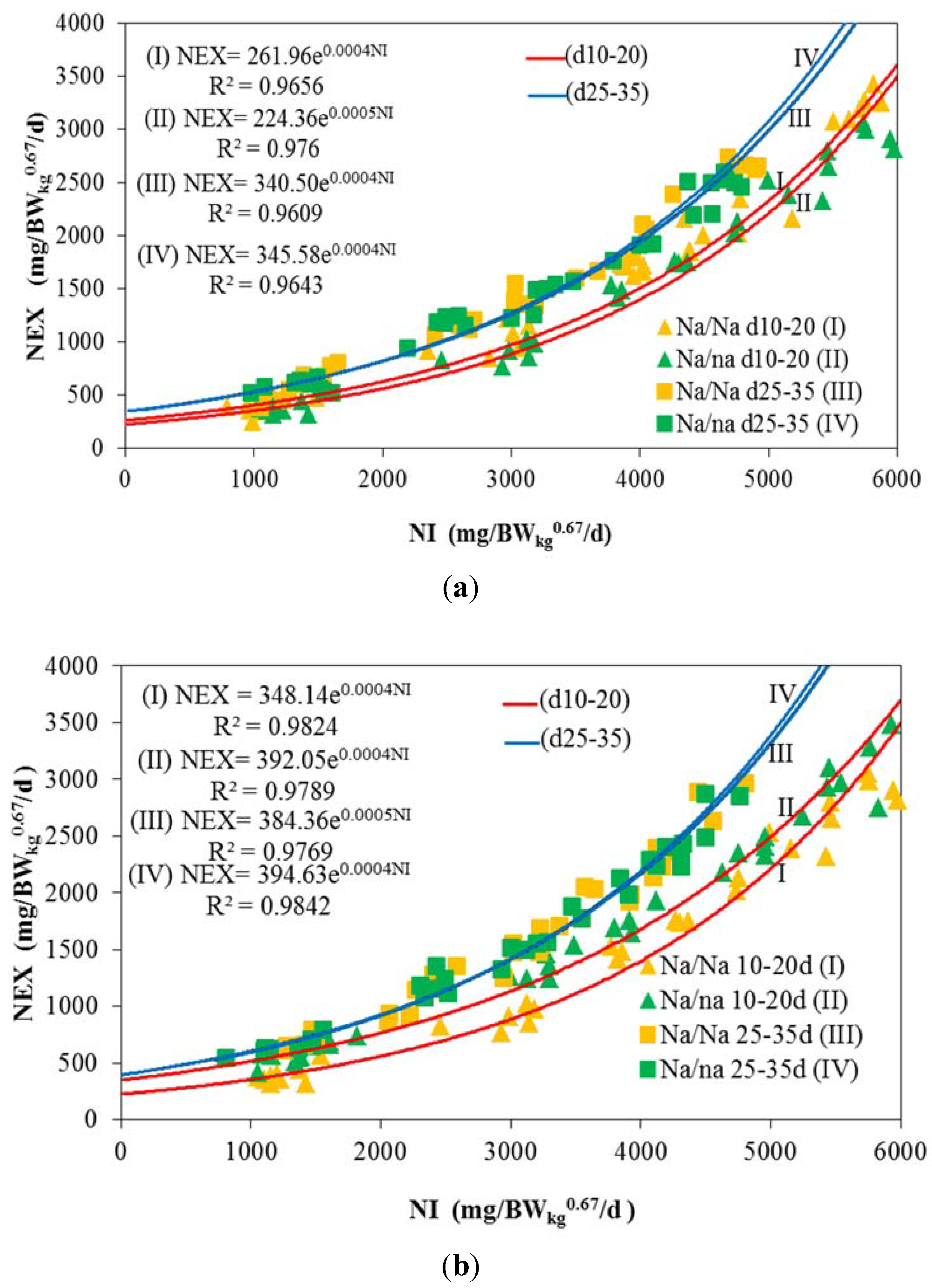
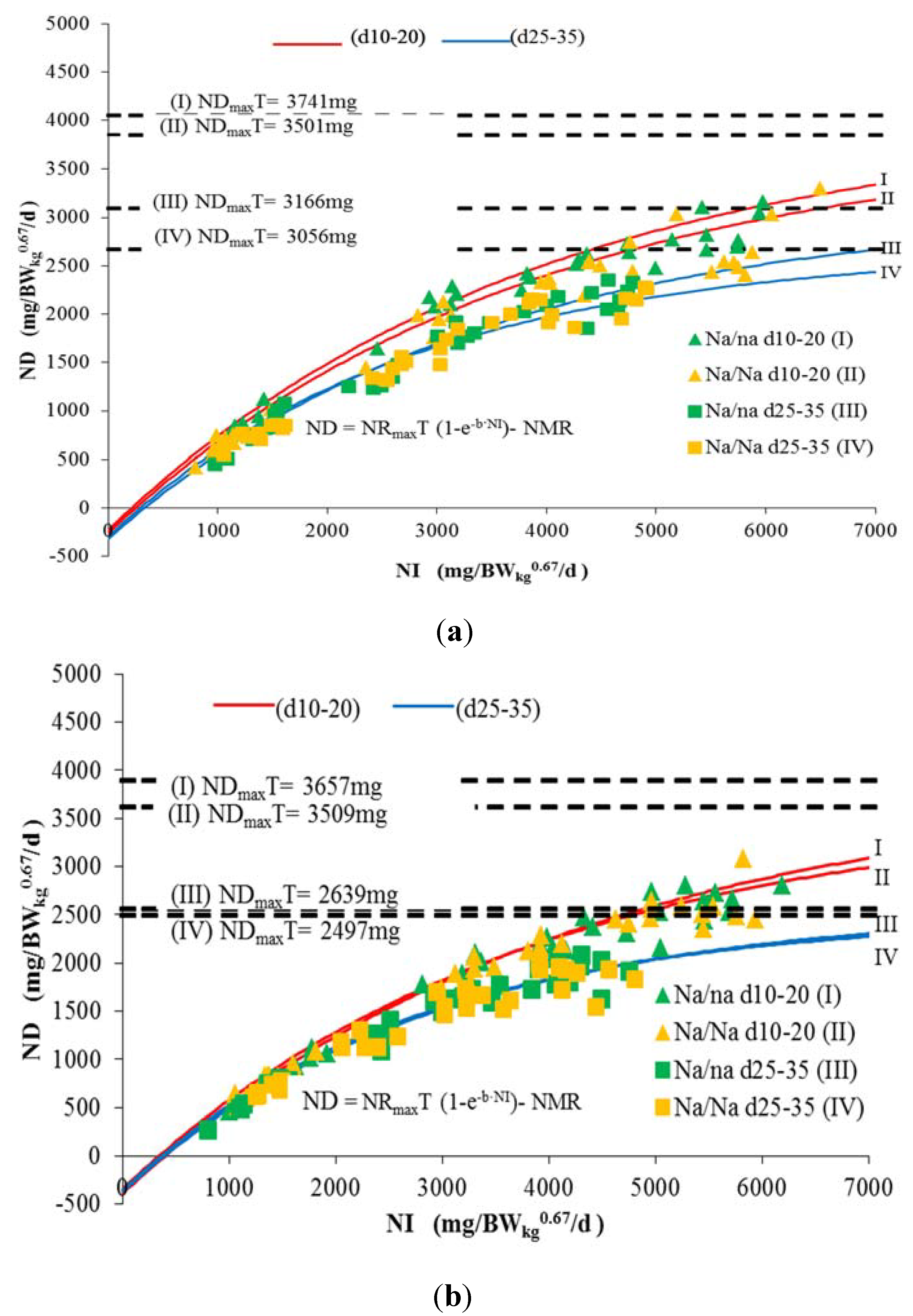
3. Results and Discussion
| Diet | Geno-Type | Sex | Age Period | Diet × Genotype | Diet × Sex | Diet × Age | Genotype × Sex | Genotype × Age | Sex × Age | |
|---|---|---|---|---|---|---|---|---|---|---|
| Probabilities | ||||||||||
| BW (g) | <0.001 | 0.633 | 0.274 | <0.001 | 0.956 | 0.945 | 0.002 | 0.432 | 0.547 | <0.001 |
| DMI (g/d) | <0.001 | 0.827 | 0.559 | <0.001 | 0.985 | 0.882 | 0.001 | 0.327 | 0.578 | <0.001 |
| NI 1 | <0.001 | 0.844 | 0.055 | <0.001 | 0.579 | 0.001 | <0.001 | 0.635 | 0.952 | 0.002 |
| NEX 1 | <0.001 | 0.113 | <0.001 | <0.001 | 0.002 | 0.120 | <0.001 | 0.103 | 0.384 | 0.004 |
| ND 1 | <0.001 | 0.079 | <0.001 | <0.001 | 0.552 | 0.007 | <0.001 | 0.030 | 0.366 | 0.194 |
| b-value 2 | <0.001 | <0.001 | <0.001 | <0.001 | 0.328 | 0.950 | 0.017 | 0.183 | 0.914 | 0.001 |
| Item | Diet (Crude Protein as % of Dry Matter) | ||||
|---|---|---|---|---|---|
| N1 (10.8) | N2 (17.4) | N3 (24.1) | N4 (30.7) | N5 (37.6) | |
| Males (Na/Na) | |||||
| BW (g) | 152a ± 7 | 312b ± 40 | 381b ± 58 | 375b ± 74 | 389b ± 46 |
| DMI (g/d) | 17.9a ± 1.1 | 47.1b ± 4.7 | 57.1b ± 7.1 | 49.9b ± 7.8 | 51.4b ± 4.7 |
| NI 2 | 1110a ± 72 | 2895b ± 115 | 4213c ± 96 | 4770d ± 171 | 5856e ± 109 |
| NEX 2 | 412a ± 33 | 1023b ± 59 | 1781c ± 58 | 2168d ± 66 | 3187e ± 44 |
| ND 2 | 697a ± 53 | 1872b ± 99 | 2432c ± 43 | 2602c ± 183 | 2669c ± 114 |
| b-value 3 | 267a ± 7 | 291a ±12 | 299a ± 4 | 311a ± 36 | 274a ± 29 |
| Females (Na/Na) | |||||
| BW (g) | 246a ± 14 | 425b ± 52 | 555b ±72 | 544b ± 65 | 478b ± 58 |
| DMI (g/d) | 31.2a ± 2.1 | 64.6b ± 4.5 | 68.2b ±5.3 | 65.7b ± 4.6 | 56.7b ± 4.6 |
| NI 2 | 1388a ± 77 | 3246b ± 68 | 3942c ±67 | 4917d ± 86 | 5659e ± 85 |
| NEX 2 | 589a ± 30 | 1348b ± 51 | 1752c ±63 | 2402d ± 68 | 3084e ± 108 |
| ND 2 | 799a ± 54 | 1898b ± 47 | 2190bc ±36 | 2515c ± 35 | 2575c ±105 |
| b-value 3 | 256a ± 5 | 270a ± 7 | 273a ± 7 | 276a ± 5 | 258a ± 24 |
| Males (Na/na) | |||||
| BW (g) | 185a ± 8 | 321ab ± 40 | 410b ± 58 | 388b ± 63 | 430b± 58 |
| DMI (g/d) | 23.2a ± 1.2 | 49.3b ± 4.6 | 58.1b ± 7.1 | 54.0b ± 7.1 | 53.6b ± 5.4 |
| NI 2 | 1249a ± 54 | 2970b ± 109 | 4066c ± 111 | 5020d ± 134 | 5718e ±95 |
| NEX 2 | 388a ± 26 | 891b ± 40 | 1609c ± 62 | 2368d ± 140 | 2787e ± 109 |
| ND2 | 862a ± 44 | 2079b ± 93 | 2458c ± 56 | 2652cd ± 51 | 2931d ±80 |
| b-value 3 | 257a ± 5 | 294a ± 9 | 278a ± 4 | 259a ± 11 | 282a ± 17 |
| Females (Na/na) | |||||
| BW (g) | 241a ± 18 | 409ab ± 48 | 494b ± 63 | 521b ±62 | 484b ± 52 |
| DMI (g/d) | 33.0a ± 3.0 | 61.4b ± 4.2 | 65.1b ± 5.0 | 61.0b ± 3.7 | 56.4b ± 3.7 |
| NI 2 | 1483a ± 98 | 3162b ± 85 | 4076c ± 60 | 4721d ± 151 | 5605e ±98 |
| NEX 2 | 666a ± 31 | 1300b ± 64 | 1867c ± 54 | 2405d ± 119 | 2956e ± 95 |
| ND 2 | 817a ± 70 | 1862b ± 70 | 2209c ± 61 | 2316c ± 129 | 2648d ± 45 |
| b-value 3 | 241a ± 3 | 258a ± 9 | 253a ± 9 | 238a ± 17 | 249a ± 8 |
| Item | Diet (Crude Protein as % of Dry Matter) | ||||
|---|---|---|---|---|---|
| N1 (9.8) | N2 (16.0) | N3 (22.6) | N4 (30.0) | N5 (36.8) | |
| Males (Na/Na) | |||||
| BW (g) | 1187a ± 45 | 1519abc ± 93 | 1526ac ± 90 | 1722c ± 114 | 1604c ± 137 |
| DMI (g/d) | 98.2a ± 6.1 | 133.2b ± 7.0 | 112.8a ± 3.4 | 116.4a ± 5.4 | 109.7a ± 6.7 |
| NI2 | 1362a ± 67 | 2571b ± 41 | 3091c ± 105 | 3899d ± 57 | 4718e ± 100 |
| NEX 2 | 608a ± 43 | 1162b ± 25 | 1404c ± 59 | 1844d ± 77 | 2606e ± 48 |
| ND 2 | 754a ± 31 | 1409b ± 40 | 1688c ± 72 | 2055d ± 41 | 2111d ± 69 |
| b-value 3 | 288a ± 8 | 282a ± 6 | 295a ± 10 | 315b ± 12 | 273a ± 10 |
| Females (Na/Na) | |||||
| BW (g) | 1180a ± 44 | 1346ab ± 68 | 1511bc ± 88 | 1544bc ± 103 | 1741c ± 79 |
| DMI (g/d) | 99.5a ± 3.5 | 108.1a ± 5.2 | 112.9a ± 2.8 | 103.9a ± 3.0 | 107.3a ± 1.8 |
| NI 2 | 1392a ± 39 | 2265b ± 83 | 3109c ± 52 | 3763d ± 116 | 4380e ± 112 |
| NEX 2 | 694a ± 22 | 1083b ± 83 | 1498c ± 60 | 1990d ± 70 | 2565e ± 132 |
| ND 2 | 698a ± 22 | 1182b ± 31 | 1611c ± 45 | 1773c ± 81 | 1816c ± 64 |
| b-value 3 | 339a ± 5 | 349a ± 15 | 382a ± 16 | 374a ± 22 | 335a ± 23 |
| Males (Na/na) | |||||
| BW (g) | 1223a ± 44 | 1418ab ± 83 | 1577b ± 96 | 1576b ± 166 | 1704b ± 93 |
| DMI (g/d) | 101.0a ± 5.2 | 122.3a ± 6.6 | 121.3a ± 4.4 | 111.7a ± 8.6 | 111.0a± 3.6 |
| NI 2 | 1381a ± 65 | 2471b ± 64 | 3245c ± 67 | 3967d ± 92 | 4593e± 53 |
| NEX 2 | 591a ± 17 | 1158b ± 45 | 1431c ± 61 | 1867d ± 51 | 2439e± 54 |
| ND 2 | 790a ± 64 | 1313b ± 38 | 1814c ± 35 | 2100c ± 45 | 2154c± 58 |
| b-value 3 | 283a ± 7 | 259a ± 6 | 295a ± 8 | 301a ± 5 | 273a± 11 |
| Females (Na/na) | |||||
| BW (g) | 1138a ± 48 | 1368ab ± 68 | 1513b ± 104 | 1536b ± 99 | 1594b ± 74 |
| DMI (g/d) | 88.1a ± 7.4 | 116.4bc ± 3.7 | 112.7c ± 3.1 | 105.0abc ± 2.5 | 100.4abc ± 2.5 |
| NI2 | 1255a ± 90 | 2414b ± 34 | 3105c ± 65 | 3814d ± 149 | 4346e ± 82 |
| NEX2 | 651a ± 26 | 1187b ± 40 | 1491c ± 43 | 1999d ± 82 | 2490e ± 86 |
| ND2 | 603a ± 68 | 1227b ± 46 | 1613c ± 37 | 1815c ± 83 | 1855c ± 43 |
| b-value3 | 319a ± 5 | 318a ± 12 | 350a ± 8 | 346a ± 16 | 314a ± 12 |
| Starter period (days 10–20) | Grower period (days 25–35) | |||||||
|---|---|---|---|---|---|---|---|---|
| Na/Na | Na/na | Na/Na | Na/na | |||||
| Males | Females | Males | Females | Males | Females | Males | Females | |
| NMR 1 | 262 | 348 | 224 | 392 | 341 | 384 | 346 | 395 |
| NRmaxT 2 | 3763 | 3857 | 3965 | 4049 | 3397 | 2881 | 3512 | 3034 |
| NDmaxT 3 | 3501 | 3509 | 3741 | 3657 | 3056 | 2497 | 3166 | 2639 |
| PDmaxT 4 | 13.8 | 13.8 | 14.7 | 14.4 | 25.1 | 20.5 | 26.0 | 21.6 |
| Probabilities | ||||||||
| NMR 1 | NRmaxT 2 | NDmaxT 3 | PDmaxT 4 | |||||
| Genotype | 0.912 | 0.626 | 0.663 | 0.827 | ||||
| Sex | 0.035 | 0.545 | 0.419 | 0.572 | ||||
| Age | 0.192 | 0.005 | 0.004 | <0.001 | ||||
3.1. Nitrogen Maintenance Requirement (NMR)

3.2. Nitrogen Deposition Potential (NDmaxT)

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mc Kay, J.C.; Barton, N.F.; Koerhuis, A.N.M.; Mc Adam, J. The challenge of genetic change in the broiler chicken. In The Challenge of Genetic Change in Animal Production; Occasional Publication No. 27; Hill, W.G., Bishop, S.C., Mc Guirk, B., Mc Kay, J.C., Simm, G., Webb, A.J., Eds.; British Society of Animal Science: Edinburgh, UK, 2000; pp. 1–7. [Google Scholar]
- Havenstein, G.B.; Ferket, P.R.; Qureshi, M.A. Growth, livability, and feed conversion of 1957 versus 2001 broilers when fed representative 1957 and 2001 broiler diets. Poult. Sci. 2003, 82, 1500–1508. [Google Scholar] [CrossRef] [PubMed]
- Janke, O.; Tzschentke, B.; Boerjan, M. Comparative Investigations of Heat Production and Body Temperature in Embryos of Modern Chicken Breeds. Avian Poult. Biol. Rev. 2004, 15, 191–196. [Google Scholar] [CrossRef]
- Garcês, A.; Horst, P. Productive performance of naked neck, frizzle and dwarf laying hens under various natural climates and two nutritional treatments. South Afr. J. Anim. Sci. 2001, 31, 174–180. [Google Scholar]
- Yunis, R.; Cahaner, A. The effects of the naked neck (Na) and frizzle (F) genes on growth and meat yield of broilers and their interactions with ambient temperatures and potential growth rate. Poult. Sci. 1999, 78, 1347–1352. [Google Scholar] [CrossRef] [PubMed]
- Ajang, O.; Prijono, S.; Smith, W. Effect of dietary protein content on growth and body composition of fast and slow feathering broiler chickens. Br. Poult. Sci. 1993, 34, 73–91. [Google Scholar] [CrossRef]
- Yalçin, S.; Özkan, S.; Açikgöz, Z.; Özkan, K. Effect of dietary protein content on live and carcase performance of heterozygous naked neck and normally feathered broilers. Br. Poult. Sci. 1996, 37, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Samadi; Liebert, F. Modeling of Threonine Requirement in Fast-Growing Chickens, Depending on Age, Sex, Protein Deposition, and Dietary Threonine Efficiency. Poult. Sci. 2006, 85, 1961–1968. [Google Scholar]
- Samadi; Liebert, F. Threonine requirement of slow-growing male chickens depends on age and dietary efficiency of threonine utilization. Poult. Sci. 2007, 86, 1140–1148. [Google Scholar]
- Samadi; Liebert, F. Modelling the optimal lysine to threonine ratio in growing chickens depending on age and efficiency of dietary amino acid utilisation. Br. Poult. Sci. 2008, 49, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Pastor, A.; Wecke, C.; Liebert, F. Assessing the age-dependent optimal dietary branched-chain amino acid ratio in growing chicken by application of a nonlinear modeling procedure. Poult. Sci. 2013, 92, 3184–3195. [Google Scholar] [CrossRef] [PubMed]
- Wecke, C.; Liebert, F. Improving the Reliability of Optimal In-Feed Amino Acid Ratios Based on Individual Amino Acid Efficiency Data from N Balance Studies in Growing Chicken. Animals 2013, 3, 558–573. [Google Scholar] [CrossRef]
- Samadi, F.; Liebert, F. Estimation of nitrogen maintenance requirements and potential for nitrogen deposition in fast-growing chickens depending on age and sex. Poult. Sci. 2006, 85, 1421–1429. [Google Scholar] [CrossRef] [PubMed]
- NRC (National Research Council). Nutrient Requirements of Poultry, 9th ed.; National Academic Press: Washigton, DC, USA, 1994. [Google Scholar]
- GRRS (German Recommendations of Requirement Standards). Empfehlungen zur Energie- und Nährstoffversorgung der Legenhennen und Masthühner (Broiler); Ausschuss für Bedarfsnormen der Gesellschaft für Ernährungsphysiologie, Ed.; DLG-Verlag: Frankfurt, Germany, 1999. [Google Scholar]
- WPSA (World’s Poultry Science Association). The prediction of apparent metabolizable energy values for poultry in compound feeds. Worlds Poult. Sci. J. 1984, 40, 181–182. [Google Scholar]
- Naumann, C.; Bassler, R. VDLUFA-Methodenbuch. Vol. III. Die Chemischen Untersuchungen von Futtermitteln; VDLUFA-Verlag: Darmstadt, Germany, 1997. [Google Scholar]
- Thong, H.T.; Liebert, F. Amino acid requirement of growing pigs depending on amino acid efficiency and level of protein deposition. 2nd communication: Threonine. Arch. Anim. Nutr. 2004, 58, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Liebert, F.; Sünder, A.; Mohamed, K. Assessment of nitrogen maintenance requirement and potential for protein deposition in juvenile Tilapia genotypes by application of an exponential nitrogen utilization. Aquaculture 2006, 261, 1346–1355. [Google Scholar] [CrossRef]
- Wecke, C.; Liebert, F. Lysine requirement studies in modern genotype barrows dependent on age, protein deposition and dietary lysine efficiency. J. Anim. Physiol. Anim. Nutr. 2009, 93, 295–304. [Google Scholar] [CrossRef]
- Liebert, F. Basics and applications of an exponential nitrogen utilization model (“Goettingen approach”) for assessing amino acid requirements in growing pigs and meat type chickens based on dietary amino acid efficiency. In Nutritional Modelling for Pigs and Poultry; Sakomura, N.K., Gous, R., Kyriazakis, I., Hauschild, L., Eds.; CABI Publishing: Wallingford, UK, 2015; pp. 73–87. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, D.R.; Wecke, C.; Sharifi, A.R.; Liebert, F. Evaluating the Age-Dependent Potential for Protein Deposition in Naked Neck Meat Type Chicken. Animals 2015, 5, 56-70. https://doi.org/10.3390/ani5010056
Khan DR, Wecke C, Sharifi AR, Liebert F. Evaluating the Age-Dependent Potential for Protein Deposition in Naked Neck Meat Type Chicken. Animals. 2015; 5(1):56-70. https://doi.org/10.3390/ani5010056
Chicago/Turabian StyleKhan, Daulat R., Christian Wecke, Ahmad R. Sharifi, and Frank Liebert. 2015. "Evaluating the Age-Dependent Potential for Protein Deposition in Naked Neck Meat Type Chicken" Animals 5, no. 1: 56-70. https://doi.org/10.3390/ani5010056