
The Metabolic Pathway of Bile Secretion Is Vulnerable to Salmonella enterica Exposure in Porcine Intestinal Epithelial Cells

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Culture
2.2. Salmonella enterica Strain Infection
2.3. Detection of mRNA Expression of hilA and invA
2.4. Cell Apoptosis Assay
2.5. Immunofluorescence Microscopy
2.6. RNA-Seq Data Processing
2.7. Gene Ontology and Pathway Analysis
2.8. Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
3.1. Evaluation of the Deleterious Effect of Salmonella enterica on IPEC-J2 Cells
3.2. Alterations in the Transcriptomic and Biological Processes in IPEC-J2 Cells under Salmonella enterica Infection
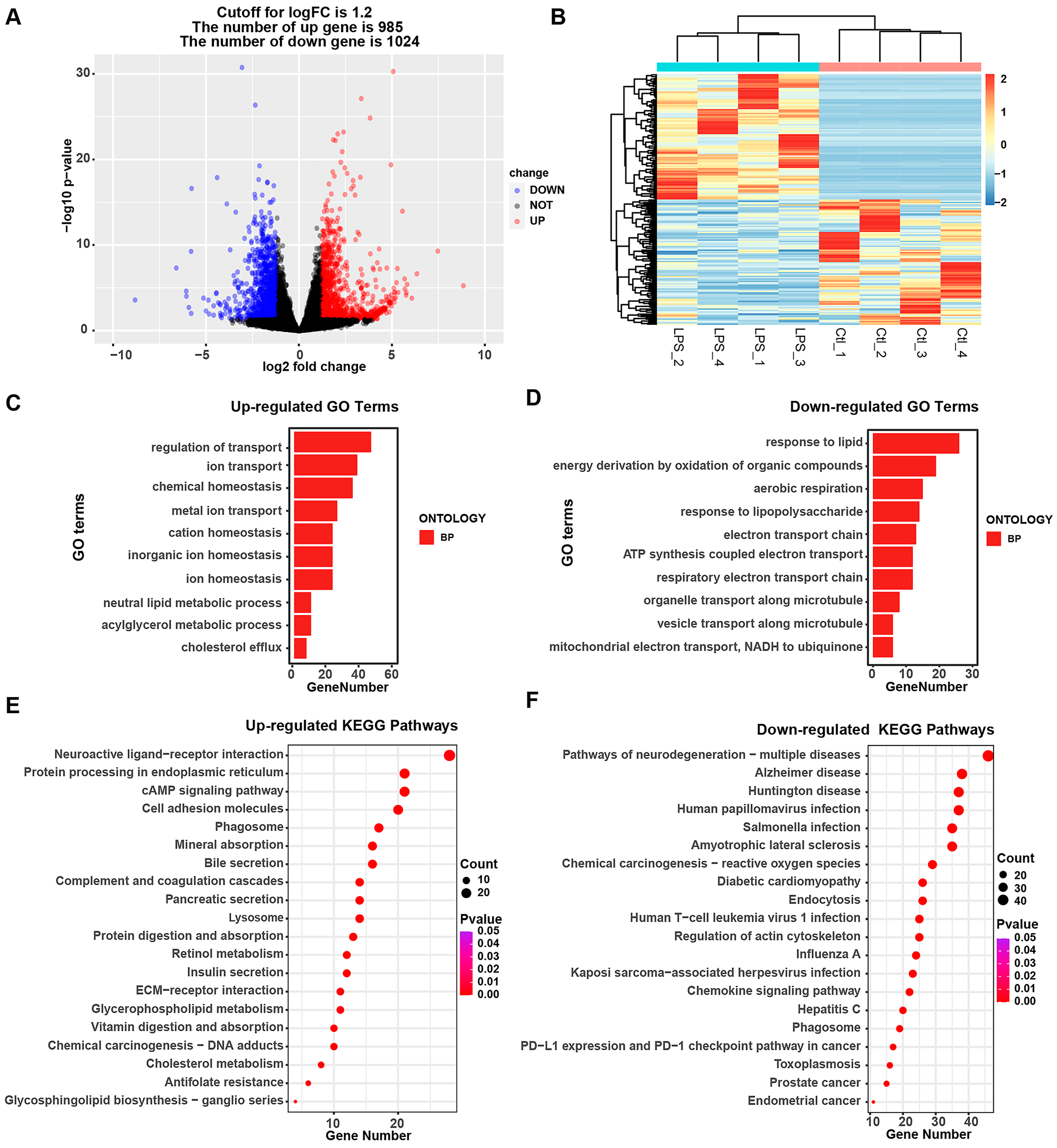
3.3. Alterations in the Transcriptomic and Biological Processes in IPEC-J2 Cells upon LPS Treatment
3.4. Integrative Analysis of the Transcriptomic Profile of IPEC-J2 Cells under Salmonella enterica and LPS Exposure
3.5. AQP8 (Aquaporin 8) Gene and Bile Secretion Pathway May Be Newly Important in IPEC-J2 Cells under Salmonella enterica or LPS Exposure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tian, Y.; Gu, D.; Wang, F.; Liu, B.; Li, J.; Kang, X.; Meng, C.; Jiao, X.; Pan, Z. Prevalence and Characteristics of Salmonella spp. from a Pig Farm in Shanghai, China. Foodborne Pathog. Dis. 2021, 18, 477–488. [Google Scholar] [CrossRef]
- Vinueza-Burgos, C.; Baquero, M.; Medina, J.; De Zutter, L. Occurrence, genotypes and antimicrobial susceptibility of Salmonella collected from the broiler production chain within an integrated poultry company. Int. J. Food Microbiol. 2019, 299, 1–7. [Google Scholar] [CrossRef]
- Smith, R.P.; May, H.E.; Burow, E.; Meester, M.; Tobias, T.J.; Sassu, E.-L.; Pavoni, E.; Di Bartolo, I.; Prigge, C.; Wasyl, D.; et al. Assessing pig farm biosecurity measures for the control of Salmonella on European farms. Epidemiol. Infect. 2023, 151, e130. [Google Scholar] [CrossRef] [PubMed]
- Fàbrega, A.; Vila, J. Salmonella enterica serovar Typhimurium skills to succeed in the host: Virulence and regulation. Clin. Microbiol. Rev. 2013, 26, 308–341. [Google Scholar] [CrossRef]
- National Institutes of Health. The European Union One Health 2020 Zoonoses Report. EFSA J. Eur. Food Saf. Auth. 2021, 19, e06971. [Google Scholar]
- Kumar, T.; Rajora, V.R.; Arora, N. Prevalence of Salmonella in pigs and broilers in the Tarai region of Uttarakhand, India. Indian J. Med. Microbiol. 2014, 32, 99. [Google Scholar] [CrossRef] [PubMed]
- Palhares, J.C.P.; Kich, J.D.; Bessa, M.C.; Biesus, L.L.; Berno, L.G.; Triques, N.J. Salmonella and antimicrobial resistance in an animal-based agriculture river system. Sci. Total Environ. 2014, 472, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Fairbrother, J.M.; Nadeau, E.; Gyles, C.L. Escherichia coli in postweaning diarrhea in pigs: An update on bacterial types, pathogenesis, and prevention strategies. Anim. Health Res. Rev. 2005, 6, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Doublet, B.; Lailler, R.; Meunier, D.; Brisabois, A.; Boyd, D.; Mulvey, M.R.; Chaslus-Dancla, E.; Cloeckaert, A. Variant Salmonella genomic island 1 antibiotic resistance gene cluster in Salmonella enterica serovar Albany. Emerg. Infect. Dis. 2003, 9, 585–591. [Google Scholar] [CrossRef]
- Narm, K.-E.; Kalafatis, M.; Slauch, J.M. HilD, HilC, and RtsA Form Homodimers and Heterodimers to Regulate Expression of the Salmonella Pathogenicity Island I Type III Secretion System. J. Bacteriol. 2020, 202, e00012-20. [Google Scholar] [CrossRef] [PubMed]
- Beshiru, A.; Igbinosa, I.H.; Igbinosa, E.O. Prevalence of Antimicrobial Resistance and Virulence Gene Elements of Salmonella Serovars From Ready-to-Eat (RTE) Shrimps. Front. Microbiol. 2019, 10, 1613. [Google Scholar] [CrossRef]
- Shi, J.; Zhao, Y.; Wang, Y.; Gao, W.; Ding, J.; Li, P.; Hu, L.; Shao, F. Inflammatory caspases are innate immune receptors for intracellular LPS. Nature 2014, 514, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Wei, G.; Wang, Y.; Ma, N.; Chang, G.; Shen, X. Sodium butyrate promotes lipopolysaccharide-induced innate immune responses by enhancing mitogen-activated protein kinase activation and histone acetylation in bovine mammary epithelial cells. J. Dairy Sci. 2020, 103, 11636–11652. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Sun, W.; Xia, D.; Wang, Y.; Li, J.; Yang, S. The m6A methyltransferase METTL3 promotes LPS-induced microglia inflammation through TRAF6/NF-κB pathway. Neuroreport 2022, 33, 243–251. [Google Scholar] [CrossRef]
- Sul, O.-J.; Ra, S.W. Quercetin Prevents LPS-Induced Oxidative Stress and Inflammation by Modulating NOX2/ROS/NF-kB in Lung Epithelial Cells. Molecules 2021, 26, 6949. [Google Scholar] [CrossRef]
- Meng, M.; Huo, R.; Wang, Y.; Ma, N.; Shi, X.; Shen, X.; Chang, G. Lentinan inhibits oxidative stress and alleviates LPS-induced inflammation and apoptosis of BMECs by activating the Nrf2 signaling pathway. Int. J. Biol. Macromol. 2022, 222, 2375–2391. [Google Scholar] [CrossRef]
- Qin, W.; Ren, Z.; Xu, C.; Cao, Y.-N.; Sun, M.-A.; Huang, R.; Bao, W. Chromatin Accessibility and Transcriptional Landscape during Inhibition of Salmonella enterica by Lactobacillus reuteri in IPEC-J2 Cells. Cells 2023, 12, 968. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.; Wang, L.; Xiong, Y.; Wang, Z.; Qiu, Y.; Wen, X.; Jiang, Z.; Yang, X.; Ma, X. Lactobacillus reuteri LR1 Improved Expression of Genes of Tight Junction Proteins via the MLCK Pathway in IPEC-1 Cells during Infection with Enterotoxigenic Escherichia coli K88. Mediat. Inflamm. 2018, 2018, 6434910. [Google Scholar] [CrossRef]
- Ibler, A.E.M.; ElGhazaly, M.; Naylor, K.L.; Bulgakova, N.A.; FEl-Khamisy, S.; Humphreys, D. Typhoid toxin exhausts the RPA response to DNA replication stress driving senescence and Salmonella infection. Nat. Commun. 2019, 10, 4040. [Google Scholar] [CrossRef]
- Draghici, S.; Khatri, P.; Tarca, A.L.; Amin, K.; Done, A.; Voichita, C.; Georgescu, C.; Romero, R. A systems biology approach for pathway level analysis. Genome Res. 2007, 17, 1537–1545. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆CT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Simoni, C.; de Campos Ausani, T.; Laviniki, V.; Lopes, G.V.; de Itapema Cardoso, M.R. Salmonella Derby from pig production chain over a 10-year period: Antimicrobial resistance, biofilm formation, and genetic relatedness. Braz. J. Microbiol. 2022, 53, 2185–2194. [Google Scholar] [CrossRef]
- Fabà, L.; Litjens, R.; Allaart, J.; van den Hil, P.R. Feed additive blends fed to nursery pigs challenged with Salmonella. J. Anim. Sci. 2020, 98, skz382. [Google Scholar] [CrossRef] [PubMed]
- Barba-Vidal, E.; Roll, V.F.B.; Castillejos, L.; Guerra-Ordaz, A.A.; Manteca, X.; Mallo, J.J.; Martín-Orúe, S.M. Response to a Salmonella Typhimurium challenge in piglets supplemented with protected sodium butyrate or Bacillus licheniformis: Effects on performance, intestinal health and behavior. Transl. Anim. Sci. 2017, 1, 186–200. [Google Scholar] [CrossRef]
- Huang, T.; Huang, X.; Shi, B.; Wang, F.; Feng, W.; Yao, M. Regulators of Salmonella-host interaction identified by peripheral blood transcriptome profiling: Roles of TGFB1 and TRP53 in intracellular Salmonella replication in pigs. Vet. Res. 2018, 49, 121. [Google Scholar] [CrossRef]
- Zhou, M.; Liu, X.; Yu, H.; Gong, J. Lactobacillus Regulates Caenorhabditis elegans Cell Signaling to Combat Salmonella Infection. Front. Immunol. 2021, 12, 653205. [Google Scholar] [CrossRef] [PubMed]
- Pitt, H.A.; Nakeeb, A. Chapter 8-Bile secretion and pathophysiology of biliary tract obstruction. In Blumgart’s Surgery of the Liver, Biliary Tract and Pancreas, 2-Volume Set, 6th ed.; Jarnagin, W.R., Ed.; Elsevier: Philadelphia, PA, USA, 2017; pp. 123–132.e1. [Google Scholar]
- Washabau, R.J. Chapter 1-Integration of Gastrointestinal Function. In Canine and Feline Gastroenterology; Washabau, R.J., Day, M.J., Eds.; W.B. Saunders: St. Louis, MO, USA, 2013; pp. 1–31. [Google Scholar]
- Song, X.; Sun, X.; Oh, S.F.; Wu, M.; Zhang, Y.; Zheng, W.; Geva-Zatorsky, N.; Jupp, R.; Mathis, D.; Benoist, C.; et al. Microbial bile acid metabolites modulate gut RORγ + regulatory T cell homeostasis. Nature 2020, 577, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Hang, S.; Paik, D.; Yao, L.; Kim, E.; Trinath, J.; Lu, J.; Ha, S.; Nelson, B.N.; Kelly, S.P.; Wu, L.; et al. Bile acid metabolites control TH17 and Treg cell differentiation. Nature 2019, 576, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Laforenza, U. Water channel proteins in the gastrointestinal tract. Mol. Asp. Med. 2012, 33, 642–650. [Google Scholar] [CrossRef]
- Xie, Y.; Zhan, X.; Tu, J.; Xu, K.; Sun, X.; Liu, C.; Ke, C.; Cao, G.; Zhou, Z.; Liu, Y. Atractylodes oil alleviates diarrhea-predominant irritable bowel syndrome by regulating intestinal inflammation and intestinal barrier via SCF/c-kit and MLCK/MLC2 pathways. J. Ethnopharmacol. 2021, 272, 113925. [Google Scholar] [CrossRef]
- Soria, L.R.; Gradilone, S.A.; Larocca, M.C.; Marinelli, R.A. Glucagon induces the gene expression of aquaporin-8 but not that of aquaporin-9 water channels in the rat hepatocyte. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R1274–R1281. [Google Scholar] [CrossRef] [PubMed]
- Calamita, G.; Ferri, D.; Bazzini, C.; Mazzone, A.; Bottà, G.; Liquori, G.E.; Paulmichl, M.; Portincasa, P.; Meyer, G.; Svelto, M. Expression and subcellular localization of the AQP8 and AQP1 water channels in the mouse gall-bladder epithelium. Biol. Cell 2005, 97, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Carreras, F.I.; Gradilone, S.A.; Mazzone, A.; García, F.; Huang, B.Q.; Ochoa, J.E.; Tietz, P.S.; Larusso, N.F.; Calamita, G.; Marinelli, R.A. Rat hepatocyte aquaporin-8 water channels are down-regulated in extrahepatic cholestasis. Hepatology 2003, 37, 1026–1033. [Google Scholar] [CrossRef] [PubMed]
- Day, R.E.; Kitchen, P.; Owen, D.S.; Bland, C.; Marshall, L.; Conner, A.C.; Bill, R.M.; Conner, M.T. Human aquaporins: Regulators of transcellular water flow. Biochim. Biophys. Acta 2014, 1840, 1492–1506. [Google Scholar] [CrossRef] [PubMed]
- Masyuk, A.I.; Marinelli, R.A.; LaRusso, N.F. Water transport by epithelia of the digestive tract. Gastroenterology 2002, 122, 545–562. [Google Scholar] [CrossRef]
- Shi, K.; Qu, L.; Lin, X.; Xie, Y.; Tu, J.; Liu, X.; Zhou, Z.; Cao, G.; Li, S.; Liu, Y. Deep-Fried Atractylodis Rhizoma Protects against Spleen Deficiency-Induced Diarrhea through Regulating Intestinal Inflammatory Response and Gut Microbiota. Int. J. Mol. Sci. 2019, 21, 124. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, J.; Chen, X.; Xu, C.; Zhu, X.; Wang, H.; Wu, S.; Cai, D.; Fan, H. The Metabolic Pathway of Bile Secretion Is Vulnerable to Salmonella enterica Exposure in Porcine Intestinal Epithelial Cells. Animals 2024, 14, 789. https://doi.org/10.3390/ani14050789
Cai J, Chen X, Xu C, Zhu X, Wang H, Wu S, Cai D, Fan H. The Metabolic Pathway of Bile Secretion Is Vulnerable to Salmonella enterica Exposure in Porcine Intestinal Epithelial Cells. Animals. 2024; 14(5):789. https://doi.org/10.3390/ani14050789
Chicago/Turabian StyleCai, Jiajia, Xiaolei Chen, Chao Xu, Xiaoyang Zhu, Haifei Wang, Shenglong Wu, Demin Cai, and Hairui Fan. 2024. "The Metabolic Pathway of Bile Secretion Is Vulnerable to Salmonella enterica Exposure in Porcine Intestinal Epithelial Cells" Animals 14, no. 5: 789. https://doi.org/10.3390/ani14050789