

Comprehensive Gene Expression Profiling Analysis of Adipose Tissue in Male Individuals from Fat- and Thin-Tailed Sheep Breeds
 ,
,  ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
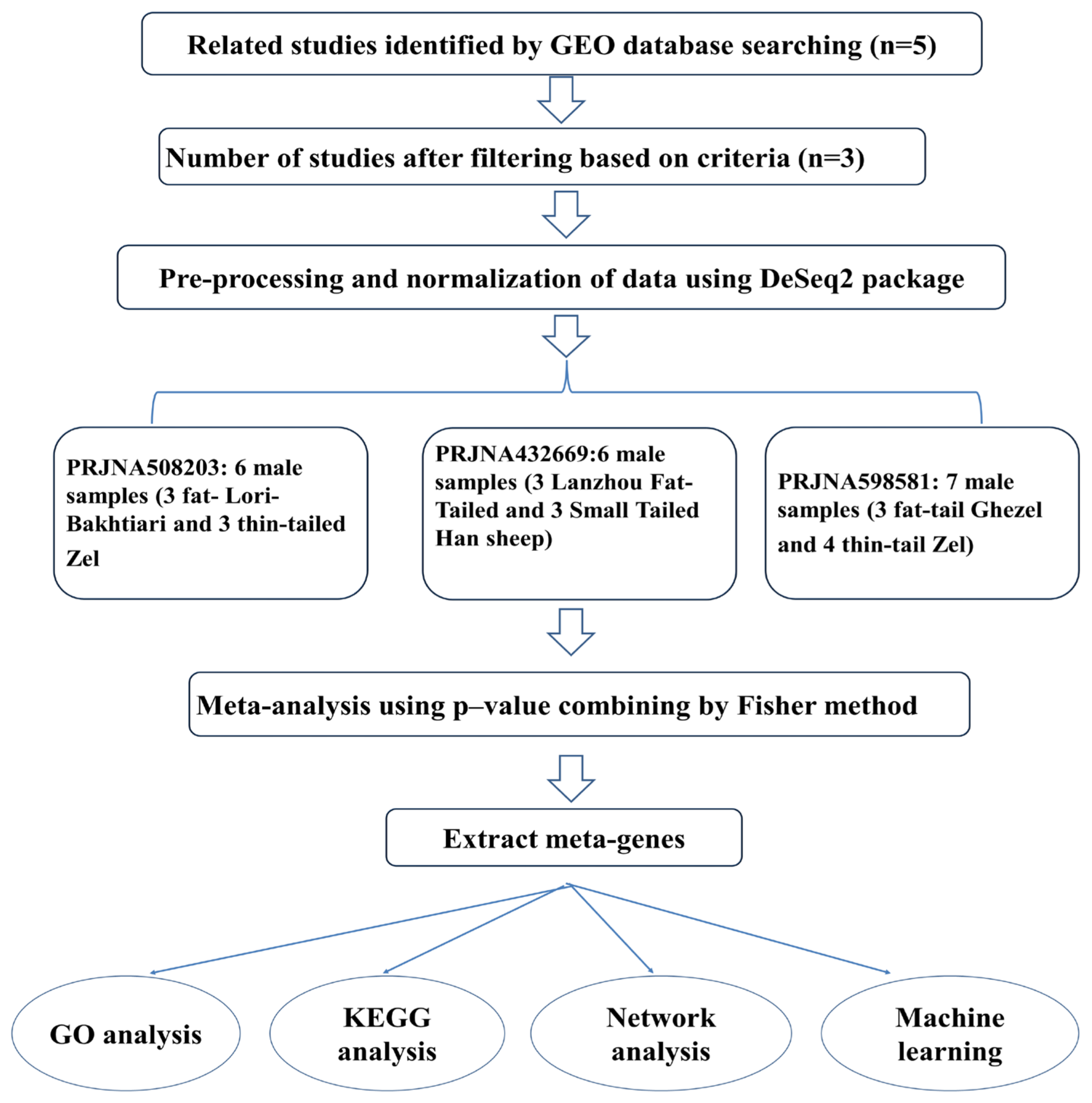
2. Materials and Methods
2.1. Dataset Collection
2.2. Quality Control, Mapping, and Differential Gene Expression Analysis
2.3. Meta-Analysis
2.4. GO Classification and KEGG Pathway Analysis
2.5. Protein–Protein Network and Module Analysis
2.6. Validation of Hub Genes Using Machine Learning Algorithms
3. Results
3.1. Sequencing Data Collection
3.2. Meta-Analysis of RNA-Seq Data
3.3. Functional Enrichment Analysis of Meta-Genes
3.4. Protein–Protein Interaction (PPI) Network and Module Analysis
3.5. Feature Selection for Machine Learning
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maggiolino, A.; Bragaglio, A.; Salzano, A.; Rufrano, D.; Claps, S.; Sepe, L.; Damiano, S.; Ciarcia, R.; Dinardo, F.R.; Hopkins, D.L.; et al. Dietary supplementation of suckling lambs with anthocyanins: Effects on growth, carcass, oxidative and meat quality traits. Anim. Feed Sci. Technol. 2021, 276, 114925. [Google Scholar] [CrossRef]
- De Palo, P.; Maggiolino, A.; Centoducati, P.; Calzaretti, G.; Ceci, E.; Tateo, A. An assessment of sire-breed effects on carcass and meat quality traits of lambs at the ages of 40 and 100 days from Comisana ewes crossed with Suffolk or Bergamasca rams. J. Anim. Prod. Sci. 2018, 58, 1794–1801. [Google Scholar] [CrossRef]
- Mohapatra, A.; Shinde, A. Fat-tailed sheep-an important sheep genetic resource for meat production in tropical countries: An overview. Indian. J. Small Rumin. 2018, 24, 1–17. [Google Scholar] [CrossRef]
- Zhou, G.; Wang, X.; Yuan, C.; Kang, D.; Xu, X.; Zhou, J.; Geng, R.; Yang, Y.; Yang, Z.; Chen, Y. Integrating miRNA and mRNA expression profiling uncovers miRNAs underlying fat deposition in sheep. BioMed Res. Int. 2017, 2017, 1857580. [Google Scholar] [CrossRef] [PubMed]
- Ryder, M.L. Sheep and Man; Gerald Duckworth & Co., Ltd.: London, UK, 1983. [Google Scholar]
- Vieira, C.; Fernández, A.M. Effect of ageing time on suckling lamb meat quality resulting from different carcass chilling regimes. Meat Sci. 2014, 96, 682–687. [Google Scholar] [CrossRef]
- Moradi, M.H.; Nejati-Javaremi, A.; Moradi-Shahrbabak, M.; Dodds, K.G.; McEwan, J.C. Genomic scan of selective sweeps in thin and fat tail sheep breeds for identifying of candidate regions associated with fat deposition. BMC Genet. 2012, 13, 10. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.-S.; Ren, X.; Yang, G.-L.; Xie, X.-L.; Zhao, Y.-X.; Zhang, M.; Shen, Z.-Q.; Ren, Y.-L.; Gao, L.; Shen, M.; et al. Genome-wide association analysis identifies the genetic basis of fat deposition in the tails of sheep (Ovis aries). Anim. Genet. 2017, 48, 560–569. [Google Scholar] [CrossRef] [PubMed]
- Mwacharo, J.M.; Kim, E.-S.; Elbeltagy, A.R.; Aboul-Naga, A.M.; Rischkowsky, B.A.; Rothschild, M.F. Genomic footprints of dryland stress adaptation in Egyptian fat-tail sheep and their divergence from East African and western Asia cohorts. Sci. Rep. 2017, 7, 17647. [Google Scholar] [CrossRef]
- Braissant, O.; Foufelle, F.; Scotto, C.; Dauça, M.; Wahli, W. Differential expression of peroxisome proliferator-activated receptors (PPARs): Tissue distribution of PPAR-alpha,-beta, and-gamma in the adult rat. Endocrinology 1996, 137, 354–366. [Google Scholar] [CrossRef]
- Li, B.; Qiao, L.; An, L.; Wang, W.; Liu, J.; Ren, Y.; Pan, Y.; Jing, J.; Liu, W. Transcriptome analysis of adipose tissues from two fat-tailed sheep breeds reveals key genes involved in fat deposition. BMC Genom. 2018, 19, 338. [Google Scholar] [CrossRef]
- Quiñones, J.; Maggiolino, A.; Bravo, S.; Muñoz, E.; Lorenzo, J.M.; Cancino, D.; Díaz, R.; Saenz, C.; Sepúlveda, N.; De Palo, P. Effect of canola oil on meat quality and fatty acid profile of Araucano creole lambs during fattening period. Anim. Feed Sci. Technol. 2019, 248, 20–26. [Google Scholar] [CrossRef]
- De Palo, P.; Tateo, A.; Maggiolino, A.; Marino, R.; Ceci, E.; Nisi, A.; Lorenzo, J.M. Martina Franca donkey meat quality: Influence of slaughter age and suckling technique. Meat Sci. 2017, 134, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Tateo, A.; Maggiolino, A.; Domínguez, R.; Lorenzo, J.M.; Dinardo, F.R.; Ceci, E.; Marino, R.; Della Malva, A.; Bragaglio, A.; De Palo, P. Volatile Organic Compounds, Oxidative and Sensory Patterns of Vacuum Aged Foal Meat. Animals 2020, 10, 1495. [Google Scholar] [CrossRef] [PubMed]
- Serrano, M.P.; Maggiolino, A.; Lorenzo, J.M.; De Palo, P.; García, A.; Landete-Castillejos, T.; Gambín, P.; Cappelli, J.; Domínguez, R.; Pérez-Barbería, F.J.; et al. Meat quality of farmed red deer fed a balanced diet: Effects of supplementation with copper bolus on different muscles. Animal 2019, 13, 888–896. [Google Scholar] [CrossRef]
- Dong, L.; Jin, Y.; Cui, H.; Yu, L.; Luo, Y.; Wang, S.; Wang, H. Effects of diet supplementation with rumen-protected betaine on carcass characteristics and fat deposition in growing lambs. Meat Sci. 2020, 166, 108154. [Google Scholar] [CrossRef] [PubMed]
- Pourlis, A.F. A review of morphological characteristics relating to the production and reproduction of fat-tailed sheep breeds. Trop. Anim. Health Prod. 2011, 43, 1267–1287. [Google Scholar] [CrossRef]
- Kalds, P.; Luo, Q.; Sun, K.; Zhou, S.; Chen, Y.; Wang, X. Trends towards revealing the genetic architecture of sheep tail patterning: Promising genes and investigatory pathways. Anim. Genet. 2021, 52, 799–812. [Google Scholar] [CrossRef]
- Ornaghi, M.G.; Guerrero, A.; Vital, A.C.P.; de Souza, K.A.; Passetti, R.A.C.; Mottin, C.; de Araújo Castilho, R.; Sañudo, C.; do Prado, I.N. Improvements in the quality of meat from beef cattle fed natural additives. Meat Sci. 2020, 163, 108059. [Google Scholar] [CrossRef]
- Luo, R.; Zhang, X.; Wang, L.; Zhang, L.; Li, G.; Zheng, Z. GLIS1, a potential candidate gene affect fat deposition in sheep tail. Mol. Biol. Rep. 2021, 48, 4925–4931. [Google Scholar] [CrossRef]
- Shao, J.; He, S.; Pan, X.; Yang, Z.; Nanaei, H.A.; Chen, L.; Li, R.; Wang, Y.; Gao, S.; Xu, H. Allele-specific expression reveals the phenotypic differences between thin-and fat-tailed sheep. J. Genet. Genom. 2020, 49, 583–586. [Google Scholar]
- Moioli, B.; Pilla, F.; Ciani, E. Signatures of selection identify loci associated with fat tail in sheep. J. Anim. Sci. 2015, 93, 4660–4669. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Wang, H.; Liu, G.; Wu, M.; Cao, J.; Liu, Z.; Liu, R.; Zhao, F.; Zhang, L.; Lu, J. Genome-wide analysis reveals population structure and selection in Chinese indigenous sheep breeds. BMC Genom. 2015, 16, 194. [Google Scholar] [CrossRef]
- Dong, K.; Yang, M.; Han, J.; Ma, Q.; Han, J.; Song, Z.; Luosang, C.; Gorkhali, N.A.; Yang, B.; He, X. Genomic analysis of worldwide sheep breeds reveals PDGFD as a major target of fat-tail selection in sheep. BMC Genom. 2020, 21, 800. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Li, S.; Liu, Q.; Wang, Z.; Zhou, Z.; Di, R.; An, X.; Miao, B.; Wang, X.; Hu, W. Rapid evolution of a retro-transposable hotspot of ovine genome underlies the alteration of BMP2 expression and development of fat tails. BMC Genom. 2019, 20, 261. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhou, G.; Xu, X.; Geng, R.; Zhou, J.; Yang, Y.; Yang, Z.; Chen, Y. Transcriptome profile analysis of adipose tissues from fat and short-tailed sheep. Gene 2014, 549, 252–257. [Google Scholar] [CrossRef]
- Ma, L.; Li, Z.; Cai, Y.; Xu, H.; Yang, R.; Lan, X. Genetic variants in fat-and short-tailed sheep from high-throughput RNA-sequencing data. Anim. Genet. 2018, 49, 483–487. [Google Scholar] [CrossRef]
- Bakhtiarizadeh, M.R.; Alamouti, A.A. RNA-Seq based genetic variant discovery provides new insights into controlling fat deposition in the tail of sheep. Sci. Rep. 2020, 10, 13525. [Google Scholar] [CrossRef]
- Farhadi, S.; Shodja Ghias, J.; Hasanpur, K.; Mohammadi, S.A.; Ebrahimie, E. Molecular mechanisms of fat deposition: IL-6 is a hub gene in fat lipolysis, comparing thin-tailed with fat-tailed sheep breeds. Arch. Anim. Breed. 2021, 64, 53–68. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, M.; Wang, J.; Wang, S.; Wang, X.; Yang, J.; Gao, L.; Gan, S. Comparative transcriptome analysis of key genes and pathways activated in response to fat deposition in two sheep breeds with distinct tail phenotype. Front. Genet. 2021, 12, 639030. [Google Scholar] [CrossRef]
- Ibrahim, A.; Baliarti, E.; Budisatria, I.; Artama, W.T.; Widayanti, R.; Maharani, D.; Tavares, L.; Margawati, E.T. Genetic diversity and relationship among Indonesian local sheep breeds on Java Island based on mitochondrial cytochrome b gene sequences. J. Genet. Eng. Biotechnol. 2023, 21, 34. [Google Scholar] [CrossRef]
- Deribe, B.; Beyene, D.; Dagne, K.; Getachew, T.; Gizaw, S.; Abebe, A. Morphological diversity of northeastern fat-tailed and northwestern thin-tailed indigenous sheep breeds of Ethiopia. Heliyon 2021, 7, e07472. [Google Scholar] [CrossRef]
- Yazdani, H.; Gholizadeh, M.; Farhadi, A.; Moradi, M.H. Study of the copy number variation on the sex chromosome in some Iranian sheep breeds. Iran. J. Anim. Sci. 2023, 54, 253–266. [Google Scholar]
- Xu, Y.-X.; Wang, B.; Jing, J.-N.; Ma, R.; Luo, Y.-H.; Li, X.; Yan, Z.; Liu, Y.-J.; Gao, L.; Ren, Y.-L. Whole-body adipose tissue multi-omic analyses in sheep reveal molecular mechanisms underlying local adaptation to extreme environments. Commun. Biol. 2023, 6, 159. [Google Scholar] [CrossRef] [PubMed]
- Mohammadabadi, M.; Kord, M.; Nazari, M. Studying expression of leptin gene in different tissues of Kermani Sheep using Real Time PCR. Agric. Biotechnol. J. 2018, 10, 111–123. [Google Scholar]
- Krawczyńska, A.; Herman, A.P.; Antushevich, H.; Bochenek, J.; Wojtulewicz, K.; Zieba, D.A. The Effect of Leptin on the Blood Hormonal Profile (Cortisol, Insulin, Thyroid Hormones) of the Ewe in Acute Inflammation in Two Different Photoperiodical Conditions. Int. J. Mol. Sci. 2022, 23, 8109. [Google Scholar] [CrossRef]
- Picó, C.; Palou, M.; Pomar, C.A.; Rodríguez, A.M.; Palou, A. Leptin as a key regulator of the adipose organ. Rev. Endocr. Metab. Disord. 2022, 23, 13–30. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, C.; Kong, Y.; Li, F.; Yue, X. Effects of intramuscular fat on meat quality and its regulation mechanism in Tan sheep. Front. Nutr. 2022, 9, 908355. [Google Scholar] [CrossRef]
- Ji, K.; Jiao, D.; Yang, G.; Degen, A.A.; Zhou, J.; Liu, H.; Wang, W.; Cong, H. Transcriptome analysis revealed potential genes involved in thermogenesis in muscle tissue in cold-exposed lambs. Front. Genet. 2022, 13, 1017458. [Google Scholar] [CrossRef]
- Zhang, Y.-M.; Erdene, K.; Zhao, Y.-B.; Li, C.-Q.; Wang, L.; Tian, F.; Ao, C.-J.; Jin, H. Role of white adipose tissue browning in cold seasonal acclimation in grazing Mongolian sheep (Ovis aries). J. Therm. Biol. 2022, 109, 103333. [Google Scholar] [CrossRef]
- Aali, M.; Moradi-Shahrbabak, H.; Moradi-Shahrbabak, M.; Sadeghi, M.; Kohram, H. Polymorphism in the SCD gene is associated with meat quality and fatty acid composition in Iranian fat-and thin-tailed sheep breeds. Livest. Sci. 2016, 188, 81–90. [Google Scholar] [CrossRef]
- Aali, M.; Shahrbabak, M.M.; Shahrbabak, H.M.; Sadeghi, M. Identifying novel SNPs and allelic sequences of the stearoyl-Coa desaturase gene (SCD) in fat-tailed and thin-tailed sheep breeds. Biochem. Genet. 2014, 52, 153–158. [Google Scholar] [CrossRef]
- Bakhtiarizadeh, M.R.; Salehi, A.; Alamouti, A.A.; Abdollahi-Arpanahi, R.; Salami, S.A. Deep transcriptome analysis using RNA-Seq suggests novel insights into molecular aspects of fat-tail metabolism in sheep. Sci. Rep. 2019, 9, 9203. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A quality control tool for high throughput sequence data. Anal. Biochem. 2010, 548, 38–43. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T. Wolfgang Huber HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621. [Google Scholar] [CrossRef] [PubMed]
- Love, M.; Anders, S.; Huber, W. Differential analysis of count data–the DESeq2 package. Genome Biol. 2014, 15, 10–1186. [Google Scholar]
- Adèr, H.J.; Mellenbergh, G.J. Research Methodology in the Social, Behavioural and Life Sciences: Designs, Models and Methods; Sage: London, UK, 1999. [Google Scholar]
- Conn, V.S.; Ruppar, T.M.; Phillips, L.J.; Chase, J.-A.D. Using meta-analyses for comparative effectiveness research. Nurs. Outlook 2012, 60, 182–190. [Google Scholar] [CrossRef]
- Rau, A.; Marot, G.; Jaffrézic, F. Differential meta-analysis of RNA-seq data from multiple studies. BMC Bioinform. 2014, 15, 91. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P. STRING v10: Protein–protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P. The STRING database in 2017: Quality-controlled protein–protein association networks, made broadly accessible. Nucleic Acids Res. 2016, 45, 362–368. [Google Scholar] [CrossRef]
- Chin, C.-H.; Chen, S.-H.; Wu, H.-H.; Ho, C.-W.; Ko, M.-T.; Lin, C.-Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef]
- Le, V.-H.; Kim, S.-R. K-strings algorithm, a new approach based on Kmeans. In Proceedings of the 2015 Conference on Research in Adaptive and Convergent Systems, New York, NY, USA, 9–12 October 2015; pp. 15–20. [Google Scholar]
- Vabalas, A.; Gowen, E.; Poliakoff, E.; Casson, A.J. Machine learning algorithm validation with a limited sample size. PLoS ONE 2019, 14, e0224365. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimie, E.; Ebrahimi, M.; Sarvestani, N.R.; Ebrahimi, M. Protein attributes contribute to halo-stability, bioinformatics approach. Saline Syst. 2011, 7, 1. [Google Scholar] [CrossRef]
- Farhadian, M.; Rafat, S.A.; Panahi, B.; Mayack, C. Weighted gene co-expression network analysis identifies modules and functionally enriched pathways in the lactation process. Sci. Rep. 2021, 11, 2367. [Google Scholar]
- Ikoma-Seki, K.; Nakamura, K.; Morishita, S.; Ono, T.; Sugiyama, K.; Nishino, H.; Hirano, H.; Murakoshi, M. Role of LRP1 and ERK and cAMP signaling pathways in lactoferrin-induced lipolysis in mature rat adipocytes. PLoS ONE 2015, 10, e0141378. [Google Scholar] [CrossRef] [PubMed]
- Carmen, G.-Y.; Víctor, S.-M. Signalling mechanisms regulating lipolysis. Cell. Signal. 2006, 18, 401–408. [Google Scholar] [CrossRef]
- Robciuc, M.R.; Kivelä, R.; Williams, I.M.; de Boer, J.F.; van Dijk, T.H.; Elamaa, H.; Tigistu-Sahle, F.; Molotkov, D.; Leppänen, V.-M.; Käkelä, R. VEGFB/VEGFR1-induced expansion of adipose vasculature counteracts obesity and related metabolic complications. Cell Metab. 2016, 23, 712–724. [Google Scholar] [CrossRef]
- Steelman, A.J.; Li, J. Astrocyte galectin-9 potentiates microglial TNF secretion. J. Neuroinflammation 2014, 11, 144. [Google Scholar] [CrossRef]
- Rodriguez-Nunez, I.; Caluag, T.; Kirby, K.; Rudick, C.N.; Dziarski, R.; Gupta, D. Nod2 and Nod2-regulated microbiota protect BALB/c mice from diet-induced obesity and metabolic dysfunction. Sci. Rep. 2017, 7, 548. [Google Scholar] [CrossRef]
- Matsuki, T.; Horai, R.; Sudo, K.; Iwakura, Y. IL-1 plays an important role in lipid metabolism by regulating insulin levels under physiological conditions. J. Exp. Med. 2003, 198, 877–888. [Google Scholar] [CrossRef]
- Bruun, J.M.; Pedersen, S.B.; Richelsen, B. Regulation of interleukin 8 production and gene expression in human adipose tissue in vitro. J. Clin. Endocrinol. Metab. 2001, 86, 1267–1273. [Google Scholar] [CrossRef]
- Acosta, J.R.; Tavira, B.; Douagi, I.; Kulyte, A.; Arner, P.; Ryden, M.; Laurencikiene, J. Human-specific function of IL-10 in adipose tissue linked to insulin resistance. J. Clin. Endocrinol. Metab. 2019, 104, 4552–4562. [Google Scholar] [CrossRef]
- Sari, A.; Dogan, S.; Nibali, L.; Koseoglu, S. Evaluation of IL-23p19/Ebi3 (IL-39) gingival crevicular fluid levels in periodontal health, gingivitis, and periodontitis. Clin. Oral. Investig. 2022, 26, 7209–7218. [Google Scholar] [CrossRef]
- Jurga, L.; Elisabet, A.N.m.; Andrea, D.; Lennart, B.; Erik, N.s.; Dominique, L.; Peter, A.; Mikael, R.n. NF-κB is important for TNF-α-induced lipolysis in human adipocytes. J. Lipid Res. 2007, 48, 1069–1077. [Google Scholar] [CrossRef]
- Zhang, H.H.; Halbleib, M.; Ahmad, F.; Manganiello, V.C.; Greenberg, A.S. Tumor necrosis factor-α stimulates lipolysis in differentiated human adipocytes through activation of extracellular signal-related kinase and elevation of intracellular cAMP. Diabetes 2002, 51, 2929–2935. [Google Scholar] [CrossRef]
- Feingold, K.R.; Doerrler, W.; Dinarello, C.A.; Fiers, W.; Grunfeld, C. Stimulation of lipolysis in cultured fat cells by tumor necrosis factor, interleukin-1, and the interferons is blocked by inhibition of prostaglandin synthesis. Endocrinology 1992, 130, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Van Hall, G.; Steensberg, A.; Sacchetti, M.; Fischer, C.; Keller, C.; Schjerling, P.; Hiscock, N.; Møller, K.; Saltin, B.; Febbraio, M.A. Interleukin-6 stimulates lipolysis and fat oxidation in humans. J. Clin. Endocrinol. Metab. 2003, 88, 3005–3010. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhang, Y.; Ye, J. IL-6: A potential role in cardiac metabolic homeostasis. Int. J. Mol. Sci. 2018, 19, 2474. [Google Scholar] [CrossRef] [PubMed]
- Contreras, G.A.; Strieder-Barboza, C.; Raphael, W. Adipose tissue lipolysis and remodeling during the transition period of dairy cows. J. Anim. Sci. Biotechnol. 2017, 8, 41. [Google Scholar] [CrossRef]
- Nonogaki, K.; Fuller, G.M.; Fuentes, N.L.; Moser, A.H.; Staprans, I.; Grunfeld, C.; Feingold, K.R. Interleukin-6 stimulates hepatic triglyceride secretion in rats. Endocrinology 1995, 136, 2143–2149. [Google Scholar] [CrossRef]
- Ma, Y.; Gao, M.; Sun, H.; Liu, D. Interleukin-6 gene transfer reverses body weight gain and fatty liver in obese mice. Biochim. Biophys. Acta 2015, 1852, 1001–1011. [Google Scholar] [CrossRef]
- Han, J.; Meng, Q.; Shen, L.; Wu, G. Interleukin-6 induces fat loss in cancer cachexia by promoting white adipose tissue lipolysis and browning. Lipids Health Dis. 2018, 17, 14. [Google Scholar] [CrossRef]
- Mariman, E.C.; Wang, P. Adipocyte extracellular matrix composition, dynamics and role in obesity. Cell. Mol. Life Sci. 2010, 67, 1277–1292. [Google Scholar] [CrossRef]
- Adapala, V.J.; Adedokun, S.A.; Considine, R.V.; Ajuwon, K.M. Acute inflammation plays a limited role in the regulation of adipose tissue COL1A1 protein abundance. J. Nutr. Biochem. 2012, 23, 567–572. [Google Scholar] [CrossRef] [PubMed]
- San, J.; Du, Y.; Wu, G.; Xu, R.; Yang, J.; Hu, J. Transcriptome analysis identifies signaling pathways related to meat quality in broiler chickens–the extracellular matrix (ECM) receptor interaction signaling pathway. Poult. Sci. 2021, 100, 101135. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Jang, M.; Kim, H.; Kwak, W.; Park, W.; Hwang, J.Y.; Lee, C.-K.; Jang, G.W.; Park, M.N.; Kim, H.-C. Comparative transcriptome analysis of adipose tissues reveals that ECM-receptor interaction is involved in the depot-specific adipogenesis in cattle. PLoS ONE 2013, 8, e66267. [Google Scholar] [CrossRef]
- Rosso, F.; Giordano, A.; Barbarisi, M.; Barbarisi, A. From cell–ECM interactions to tissue engineering. J. Cell. Physiol. 2004, 199, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Franchini, M.; Monnais, E.; Seboek, D.; Radimerski, T.; Zini, E.; Kaufmann, K.; Lutz, T.; Reusch, C.; Ackermann, M.; Muller, B. Insulin resistance and increased lipolysis in bone marrow derived adipocytes stimulated with agonists of Toll-like receptors. Horm. Metab. Res. 2010, 42, 703–709. [Google Scholar] [CrossRef]
- Suganami, T.; Tanimoto-Koyama, K.; Nishida, J.; Itoh, M.; Yuan, X.; Mizuarai, S.; Kotani, H.; Yamaoka, S.; Miyake, K.; Aoe, S. Role of the Toll-like receptor 4/NF-κB pathway in saturated fatty acid–induced inflammatory changes in the interaction between adipocytes and macrophages. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 84–91. [Google Scholar] [CrossRef]
- Zu, L.; He, J.; Jiang, H.; Xu, C.; Pu, S.; Xu, G. Bacterial endotoxin stimulates adipose lipolysis via toll-like receptor 4 and extracellular signal-regulated kinase pathway. J. Biol. Chem. 2009, 284, 5915–5926. [Google Scholar] [CrossRef] [PubMed]
- Purohit, J.S.; Hu, P.; Chen, G.; Whelan, J.; Moustaid-Moussa, N.; Zhao, L. Activation of nucleotide oligomerization domain containing protein 1 induces lipolysis through NF-κB and the lipolytic PKA activation in 3T3-L1 adipocytes. Biochem. Cell Biol. 2013, 91, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.; Ding, Y.-F.; Jia, W.-H.; Liu, X.-L.; Feng, J.-Y.; Zhu, Q.; Cai, S.-L.; Yang, Y.-S.; Lu, Q.-Y.; Huang, X.-T. SET domain–containing protein 4 epigenetically controls breast cancer stem cell quiescence. Cancer Res. 2019, 79, 4729–4743. [Google Scholar] [CrossRef] [PubMed]
- Graja, A.; Garcia-Carrizo, F.; Jank, A.M.; Gohlke, S.; Ambrosi, T.H.; Jonas, W.; Ussar, S.; Kern, M.; Schürmann, A.; Aleksandrova, K. Loss of periostin occurs in aging adipose tissue of mice and its genetic ablation impairs adipose tissue lipid metabolism. Aging Cell 2018, 17, e12810. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GEO Accession Number | Number of Samples | Tissue Sample | Age of Slaughter (Month) | Read Length | Reference | |
|---|---|---|---|---|---|---|
| Thin-Tailed | Fat-Tailed | |||||
| PRJNA432669 | 3 | 3 | Tail | 6 | 150 bp | [11] |
| PRJNA508203 | 3 | 3 | Tail | 6 | 150 bp | [43] |
| PRJNA598581 | 4 | 3 | Tail | 6 | 150 bp | [29] |
| Biological Process Terms | Adjusted p-Value |
|---|---|
| Positive regulation of T cell cytokine production | 0.002 |
| Extracellular matrix organization | 0.004 |
| Stress-activated MAPK cascade | 0.008 |
| ERK1 and ERK2 cascade | 0.008 |
| Positive regulation of interleukin-10 production | 0.009 |
| Positive regulation of interleukin-1 secretion | 0.016 |
| Positive regulation of interleukin-6 production | 0.034 |
| Positive regulation of interleukin-8 production | 0.036 |
| Positive regulation of interferon-gamma (IFN)secretion | 0.04 |
| Positive regulation of tumor necrosis factor (TNF) production | 0.04 |
| Attribute | Weight_Info Gain Ratio | Weight_Rule | Weight_Chi Squared | Weight_Gini Index | Weight_Uncertainty | Weight_Relief | Weight_Info Gain | Average_Weight |
|---|---|---|---|---|---|---|---|---|
| POSTN | 1 | 1 | 0.5 | 0.8 | 0.6 | 0.6 | 0.8 | 0.8 |
| K35 | 0.8 | 0.9 | 0.6 | 0.6 | 0.7 | 1 | 0.6 | 0.8 |
| SETD4 | 0.8 | 1.0 | 0.7 | 0.7 | 0.6 | 0.9 | 0.6 | 0.8 |
| USP29 | 0.7 | 0.8 | 0.8 | 0.5 | 0.7 | 1.0 | 0.5 | 0.7 |
| ANKRD37 | 0.7 | 0.9 | 0.8 | 0.5 | 0.7 | 0.9 | 0.5 | 0.7 |
| ENSOARG00000001454 | 0.8 | 1 | 0.5 | 0.6 | 0.4 | 0.7 | 0.6 | 0.7 |
| RTN2 | 0.7 | 1 | 0.6 | 0.5 | 0.6 | 0.8 | 0.5 | 0.7 |
| PRG4 | 0.8 | 1 | 0.3 | 0.6 | 0.4 | 0.9 | 0.6 | 0.7 |
| LRRC4C | 0.7 | 0.8 | 0.6 | 0.5 | 0.5 | 1 | 0.5 | 0.7 |
| Model | Accuracy |
|---|---|
| Random Forest with accuracy criterion | 90% +/− 22.36% |
| Random Forest with gain_ratio criterion | 85% +/− 13.69% |
| Decision Tree with gain_ratio criterion | 58.33% +/− 37.27% |
| Decision Tree with accuracy criterion | 75% +/− 35.36% |
| Deep Learning with Tanh parameter | 85% +/− 22.36% |
| Deep Learning with Rectifier parameter | 75% +/− 25.00% |
| Deep Learning with Maxout parameter | 56.67% +/− 18.07% |
| Naïve Bayes | 78.33% +/− 21.73% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farhadi, S.; Hasanpur, K.; Ghias, J.S.; Palangi, V.; Maggiolino, A.; Landi, V. Comprehensive Gene Expression Profiling Analysis of Adipose Tissue in Male Individuals from Fat- and Thin-Tailed Sheep Breeds. Animals 2023, 13, 3475. https://doi.org/10.3390/ani13223475
Farhadi S, Hasanpur K, Ghias JS, Palangi V, Maggiolino A, Landi V. Comprehensive Gene Expression Profiling Analysis of Adipose Tissue in Male Individuals from Fat- and Thin-Tailed Sheep Breeds. Animals. 2023; 13(22):3475. https://doi.org/10.3390/ani13223475
Chicago/Turabian StyleFarhadi, Sana, Karim Hasanpur, Jalil Shodja Ghias, Valiollah Palangi, Aristide Maggiolino, and Vincenzo Landi. 2023. "Comprehensive Gene Expression Profiling Analysis of Adipose Tissue in Male Individuals from Fat- and Thin-Tailed Sheep Breeds" Animals 13, no. 22: 3475. https://doi.org/10.3390/ani13223475