
Understanding Age-Related Longitudinal Dynamics in Abundance and Diversity of Dominant Culturable Gut Lactic Acid Bacteria in Pastured Goats

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Husbandry
2.2. Fecal Sample Collection
2.3. Microbial Isolation and Identification of Lactic Acid Bacteria in Fecal Samples
2.4. Total DNA Extraction
2.5. LAB Abundance Quantification Using qPCR
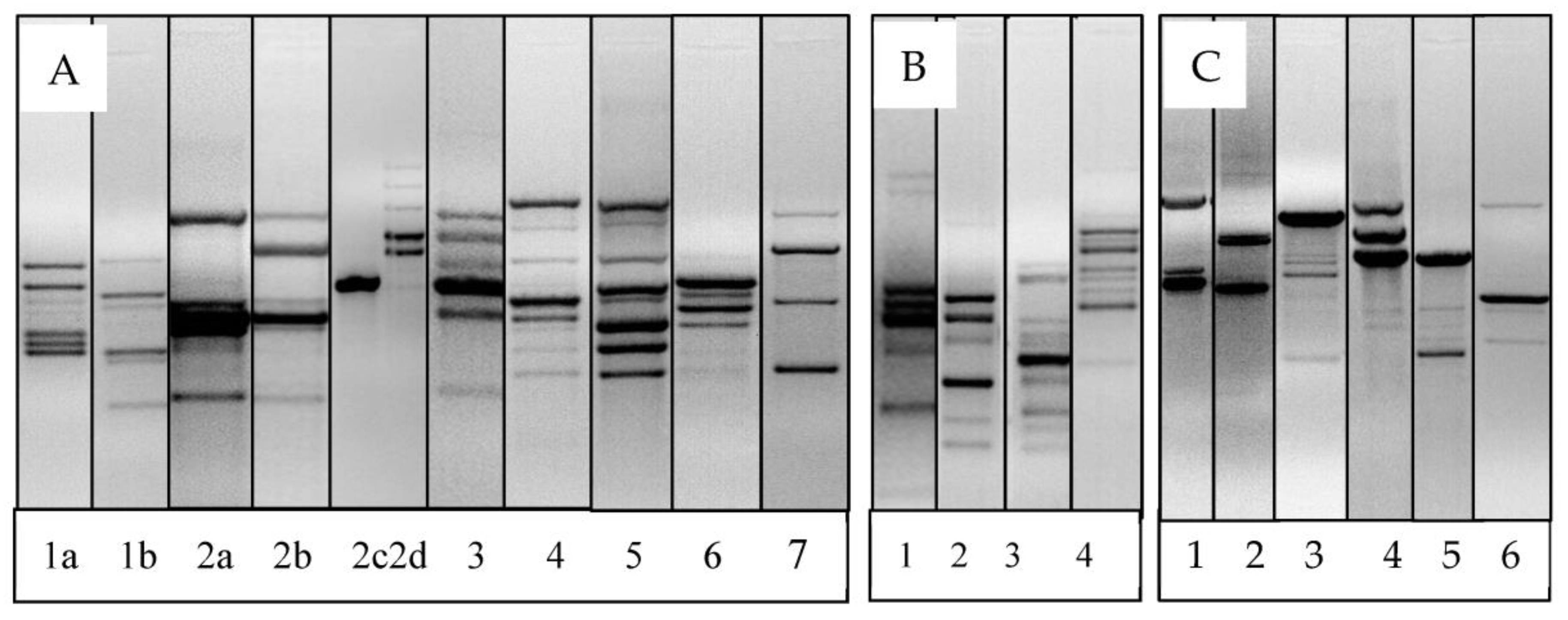
2.6. Microbial DNA Extraction, RAPD-PCR and Analysis of RAPD-PCR Fingerprints
2.7. PCR Amplification and DNA Sequencing of Lactic Acid Bacteria
2.8. Statistical Analysis
3. Results
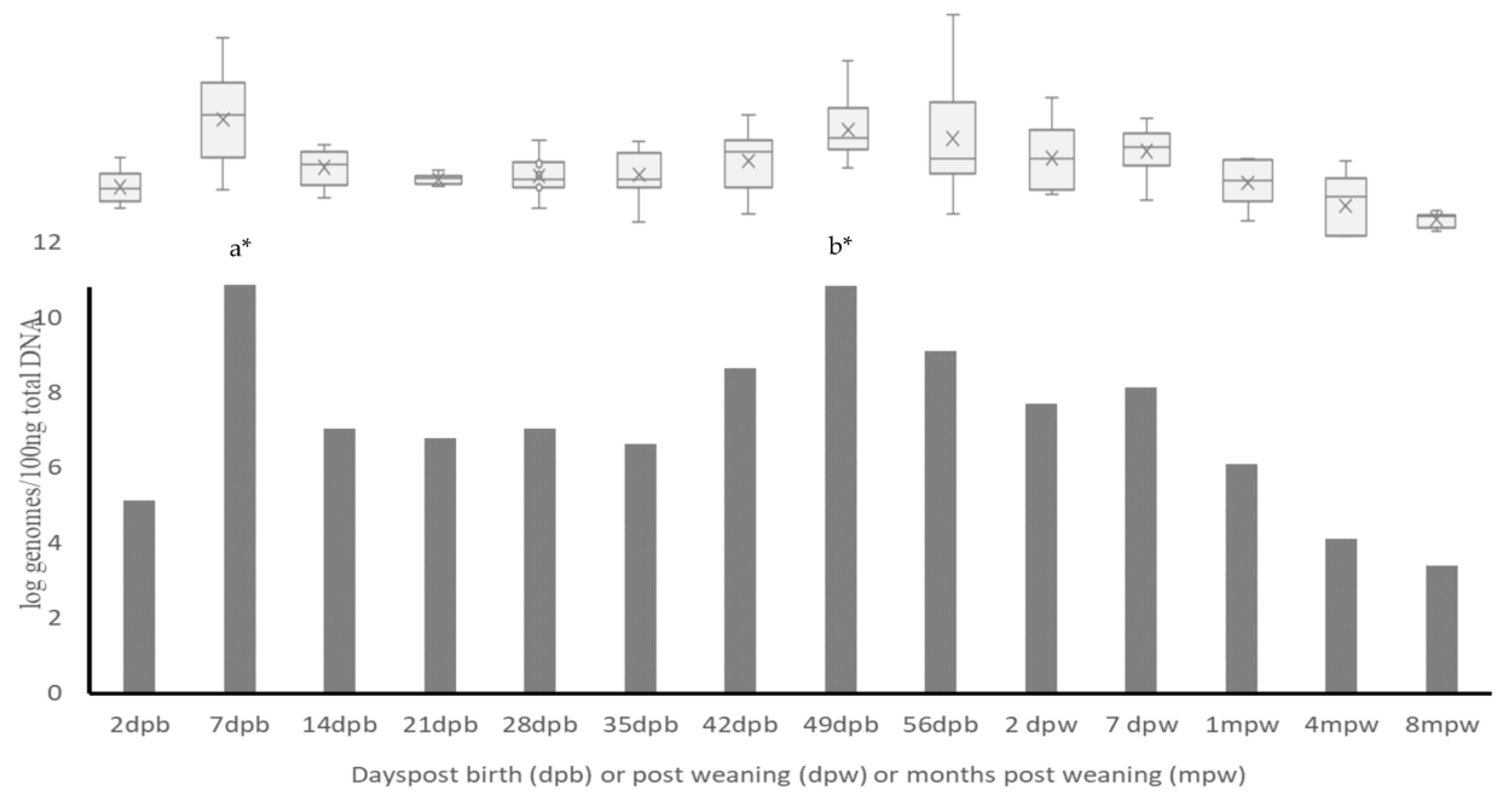
3.1. Changes in Abundance of LAB during Growing Period
3.2. Diversity and Prevalence of LAB at Different Age Groups of Goats
3.3. Frequency of Detection of LAB Strains in Individual Animals Following Repeated Sampling
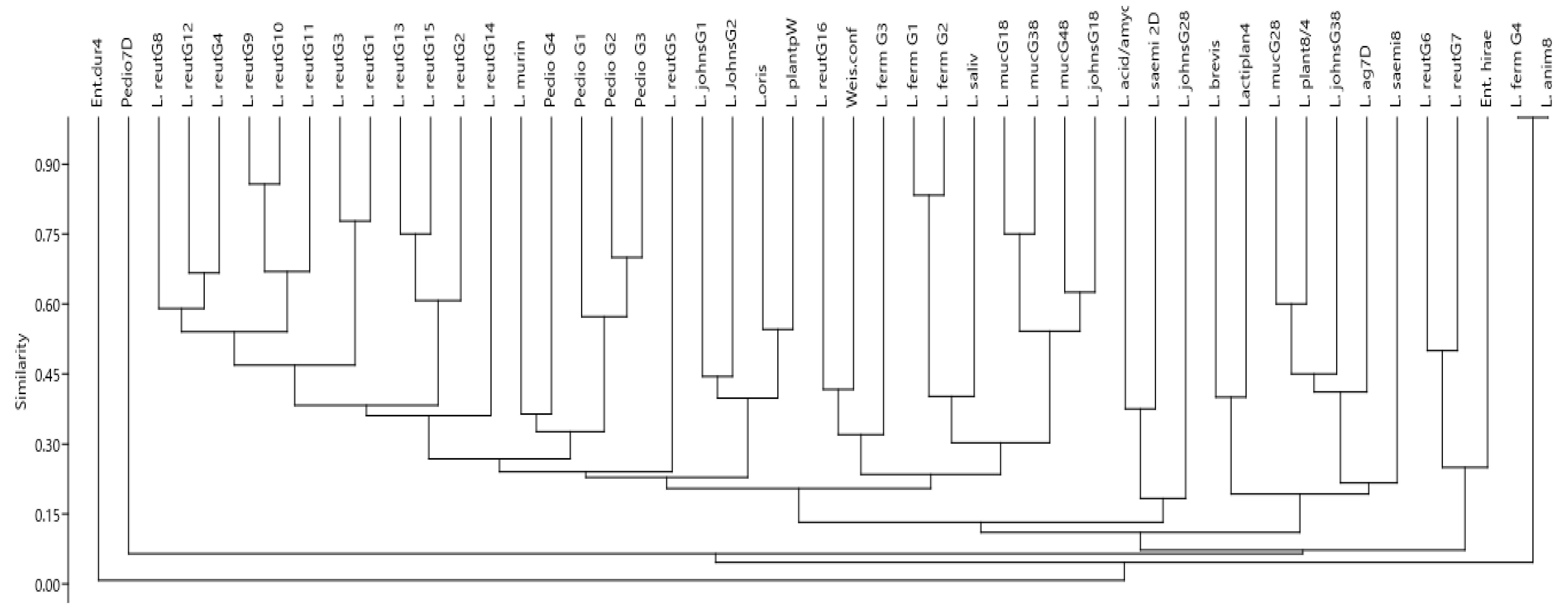
3.4. Genotypic Diversity of LAB from Goats as Evaluated with RAPD
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rattanachaikunsopon, P.; Phumkhachorn, P. Lactic acid bacteria: Their antimicrobial compounds and their uses in food production. Ann. Biol. Res. 2010, 1, 218–228. [Google Scholar]
- König, H.; Fröhlich, J. Lactic acid bacteria. In Biology of Microorganisms on Grapes, in Must and in Wine; Springer: Cham, Switzerland, 2017; pp. 3–41. [Google Scholar]
- Khalid, K. An overview of lactic acid bacteria. Int. J. Biosci. 2011, 1, 1–13. [Google Scholar]
- Masood, M.I.; Qadir, M.I.; Shirazi, J.H.; Khan, I.U. Beneficial effects of lactic acid bacteria on human beings. Crit. Rev. Microbiol. 2011, 37, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Ringø, E.; Hoseinifar, S.H.; Ghosh, K.; Doan, H.V.; Beck, B.R.; Song, S.K. Lactic acid bacteria in finfish—An update. Front. Microbiol. 2018, 9, 1818. [Google Scholar] [CrossRef]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Gancel, F.; Kempf, I.; Drider, D. Benefits and inputs from lactic acid bacteria and their bacteriocins as alternatives to antibiotic growth promoters during food-animal production. Front. Microbiol. 2019, 10, 57. [Google Scholar] [CrossRef]
- Laiño, J.; Villena, J.; Kanmani, P.; Kitazawa, H. Immunoregulatory effects triggered by lactic acid bacteria exopolysaccharides: New insights into molecular interactions with host cells. Microorganisms 2016, 4, 27. [Google Scholar] [CrossRef]
- Tojo, R.; Suárez, A.; Clemente, M.G.; de los Reyes-Gavilán, C.G.; Margolles, A.; Gueimonde, M.; Ruas-Madiedo, P. Intestinal microbiota in health and disease: Role of bifidobacteria in gut homeostasis. World J. Gastroenterol. WJG 2014, 20, 15163. [Google Scholar] [CrossRef]
- Tsuda, H.; Miyamoto, T. Guidelines for the evaluation of probiotics in food. Report of a joint FAO/WHO working group on drafting guidelines for the evaluation of probiotics in food Guidelines for the evaluation of probiotics in food. Report of a joint FAO/WHO working group on drafting guidelines for the evaluation of probiotics in food, 2002. Food Sci. Technol. Res. 2010, 16, 87–92. [Google Scholar]
- Balasingham, K.; Valli, C.; Radhakrishnan, L.; Balasuramanyam, D. Probiotic characterization of lactic acid bacteria isolated from swine intestine. Vet. World 2017, 10, 825. [Google Scholar] [CrossRef]
- Agazzi, A.; Tirloni, E.; Stella, S.; Maroccolo, S.; Ripamonti, B.; Bersani, C.; Caputo, J.M.; Dell’Orto, V.; Rota, N.; Savoini, G. Effects of species-specific probiotic addition to milk replacer on calf health and performance during the first month of life. Ann. Anim. Sci. 2014, 14, 101–115. [Google Scholar] [CrossRef]
- Timmerman, H.M.; Mulder, L.; Everts, H.; Van Espen, D.; Van Der Wal, E.; Klaassen, G.; Rouwers, S.; Hartemink, R.; Rombouts, F.; Beynen, A. Health and growth of veal calves fed milk replacers with or without probiotics. J. Dairy Sci. 2005, 88, 2154–2165. [Google Scholar] [CrossRef] [PubMed]
- Heilig, H.G.; Zoetendal, E.G.; Vaughan, E.E.; Marteau, P.; Akkermans, A.D.; de Vos, W.M. Molecular diversity of Lactobacillus spp. and other lactic acid bacteria in the human intestine as determined by specific amplification of 16S ribosomal DNA. Appl. Environ. Microbiol. 2002, 68, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zhang, K.; Li, C.; Wang, X.; Chen, Y.; Yang, Y. Characterization and Comparison of Microbiota in the Gastrointestinal Tracts of the Goat (Capra hircus) During Preweaning Development. Front. Microbiol. 2019, 10, 2125. [Google Scholar] [CrossRef]
- Guo, J.; Li, P.; Zhang, K.; Zhang, L.; Wang, X.; Li, L.; Zhang, H. Distinct Stage Changes in Early-Life Colonization and Acquisition of the Gut Microbiota and Its Correlations With Volatile Fatty Acids in Goat Kids. Front. Microbiol. 2020, 11, 584742. [Google Scholar] [CrossRef]
- Zhang, K.; Li, B.; Guo, M.; Liu, G.; Yang, Y.; Wang, X.; Chen, Y.; Zhang, E. Maturation of the Goat Rumen Microbiota Involves Three Stages of Microbial Colonization. Animals 2019, 9, 1028. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, H.; Zhu, L.; Xu, Y.; Liu, N.; Sun, X.; Hu, L.; Huang, H.; Wei, K.; Zhu, R. Dynamic Distribution of Gut Microbiota in Goats at Different Ages and Health States. Front. Microbiol. 2018, 9, 2509. [Google Scholar] [CrossRef] [PubMed]
- Emu, Q.; Guan, H.; Zhu, J.; Zhang, L.; Fan, J.; Ji, Y.; Lin, Y.; Li, C.; Dan, X.; Aguo, Y.; et al. Grazing and Supplementation of Dietary Yeast Probiotics Shape the Gut Microbiota and Improve the Immunity of Black Fattening Goats (Capra hircus). Front. Microbiol. 2021, 12, 666837. [Google Scholar] [CrossRef] [PubMed]
- McLoughlin, S.; Spillane, C.; Claffey, N.; Smith, P.E.; O’Rourke, T.; Diskin, M.G.; Waters, S.M. Rumen Microbiome Composition Is Altered in Sheep Divergent in Feed Efficiency. Front. Microbiol. 2020, 11, 1981. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Howe, S.; Wei, X.; Deng, F.; Tsai, T.; Chai, J.; Xiao, Y.; Yang, H.; Maxwell, C.V.; Li, Y. Comprehensive cultivation of the swine gut microbiome reveals high bacterial diversity and guides bacterial isolation in pigs. Msystems 2021, 6, e00477-21. [Google Scholar] [CrossRef]
- Allen-Vercoe, E. Bringing the gut microbiota into focus through microbial culture: Recent progress and future perspective. Curr. Opin. Microbiol. 2013, 16, 625–629. [Google Scholar] [CrossRef]
- Clavel, T.; Gomes-Neto, J.C.; Lagkouvardos, I.; Ramer-Tait, A.E. Deciphering interactions between the gut microbiota and the immune system via microbial cultivation and minimal microbiomes. Immunol. Rev. 2017, 279, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Fenske, G.J.; Ghimire, S.; Antony, L.; Christopher-Hennings, J.; Scaria, J. Integration of culture-dependent and independent methods provides a more coherent picture of the pig gut microbiome. FEMS Microbiol. Ecol. 2020, 96, fiaa022. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, N.; Gu, Q.; Li, P. Whole genome sequence analysis and in vitro probiotic characteristics of a Lactobacillus strain Lactobacillus paracasei ZFM54. J. Appl. Microbiol. 2020, 129, 422–433. [Google Scholar] [CrossRef]
- Sharma, A.; Lee, S.; Park, Y.-S. Molecular typing tools for identifying and characterizing lactic acid bacteria: A review. Food Sci. Biotechnol. 2020, 29, 1301–1318. [Google Scholar] [CrossRef] [PubMed]
- Cho, G.-S.; Cappello, C.; Schrader, K.; Fagbemigun, O.; Oguntoyinbo, F.A.; Csovcsics, C.; Rosch, N.; Kabisch, J.; Neve, H.; Bockelmann, W. Isolation and characterization of lactic acid bacteria from fermented goat milk in Tajikistan. J. Microbiol. Biotechnol. 2018, 28, 1834–1845. [Google Scholar] [CrossRef]
- Puntillo, M.; Gaggiotti, M.; Oteiza, J.M.; Binetti, A.; Massera, A.; Vinderola, G. Potential of lactic acid bacteria isolated from different forages as silage inoculants for improving fermentation quality and aerobic stability. Front. Microbiol. 2020, 11, 586716. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, P.; Seseña, S.; Palop, M.L. A comparative study of different PCR-based DNA fingerprinting techniques for typing of lactic acid bacteria. Eur. Food Res. Technol. 2014, 239, 87–98. [Google Scholar] [CrossRef]
- Plengvidhya, V.; Breidt, F., Jr.; Fleming, H. Use of RAPD-PCR as a method to follow the progress of starter cultures in sauerkraut fermentation. Int. J. Food Microbiol. 2004, 93, 287–296. [Google Scholar] [CrossRef]
- Randazzo, C.; Caggia, C.; Neviani, E. Application of molecular approaches to study lactic acid bacteria in artisanal cheeses. J. Microbiol. Methods 2009, 78, 1–9. [Google Scholar] [CrossRef]
- Rossetti, L.; Giraffa, G. Rapid identification of dairy lactic acid bacteria by M13-generated, RAPD-PCR fingerprint databases. J. Microbiol. Methods 2005, 63, 135–144. [Google Scholar] [CrossRef]
- Albesharat, R.; Ehrmann, M.A.; Korakli, M.; Yazaji, S.; Vogel, R.F. Phenotypic and genotypic analyses of lactic acid bacteria in local fermented food, breast milk and faeces of mothers and their babies. Syst. Appl. Microbiol. 2011, 34, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Ashmaig, A.; Hasan, A.; El Gaali, E. Identification of lactic acid bacteria isolated from traditional Sudanese fermented camels milk (Gariss). Afr. J. Microbiol. Res. 2009, 3, 451–457. [Google Scholar]
- Franciosi, E.; Settanni, L.; Cavazza, A.; Poznanski, E. Biodiversity and technological potential of wild lactic acid bacteria from raw cows’ milk. Int. Dairy J. 2009, 19, 3–11. [Google Scholar] [CrossRef]
- Ben Amor, K.; Vaughan, E.E.; de Vos, W.M. Advanced molecular tools for the identification of lactic acid bacteria. J. Nutr. 2007, 137, 741S–747S. [Google Scholar] [CrossRef]
- Rinttilä, T.; Kassinen, A.; Malinen, E.; Krogius, L.; Palva, A. Development of an extensive set of 16S rDNA-targeted primers for quantification of pathogenic and indigenous bacteria in faecal samples by real-time PCR. J. Appl. Microbiol. 2004, 97, 1166–1177. [Google Scholar] [CrossRef] [PubMed]
- Huey, B.; Hall, J. Hypervariable DNA fingerprinting in Escherichia coli: Minisatellite probe from bacteriophage M13. J. Bacteriol. 1989, 171, 2528–2532. [Google Scholar] [CrossRef]
- Giraffa, G.; Rossetti, L.; Neviani, E. An evaluation of chelex-based DNA purification protocols for the typing of lactic acid bacteria. J. Microbiol. Methods 2000, 42, 175–184. [Google Scholar] [CrossRef]
- Garcia-Vallvé, S.; Palau, J.; Romeu, A. Horizontal gene transfer in glycosyl hydrolases inferred from codon usage in Escherichia coli and Bacillus subtilis. Mol. Biol. Evol. 1999, 16, 1125–1134. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.; Ryan, P. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 4. [Google Scholar]
- Korhonen, J.; Sclivagnotis, Y.; Wright, A.V. Characterization of dominant cultivable lactobacilli and their antibiotic resistance profiles from faecal samples of weaning piglets. J. Appl. Microbiol. 2007, 103, 2496–2503. [Google Scholar] [CrossRef]
- Castro, B.G.; Souza, M.; Regua-Mangia, A.H.; Bittencourt, A.J. Genetic relationship between Escherichia coli strains isolated from dairy mastitis and from the stable fly Stomoxys calcitrans. Pesqui. Vet. Bras. 2016, 36, 479–484. [Google Scholar] [CrossRef]
- Chagnaud, P.; Machinis, K.; Coutte, L.c.A.; Marecat, A.; Mercenier, A. Rapid PCR-based procedure to identify lactic acid bacteria: Application to six common Lactobacillus species. J. Microbiol. Methods 2001, 44, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Huang, X.; Wang, P.; Yan, Z.; Sun, W.; Zhao, S.; Gun, S. Longitudinal development of the gut microbiota in healthy and diarrheic piglets induced by age-related dietary changes. Microbiologyopen 2019, 8, e923. [Google Scholar] [CrossRef] [PubMed]
- La-Ongkham, O.; Nakphaichit, M.; Nakayama, J.; Keawsompong, S.; Nitisinprasert, S. Age-related changes in the gut microbiota and the core gut microbiome of healthy Thai humans. 3 Biotech 2020, 10, 276. [Google Scholar] [CrossRef]
- Feng, L.; Zhu, C.; Xia, P.; Wang, H.; Su, Y.; Zhu, W. The development of intestinal lactic acid bacteria in piglets as determined by high-throughput sequencing. Anim. Biotechnol. 2021, 34, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhao, J.; Song, Y.; Zhang, J.; Yu, Z.; Zhang, H.; Sun, Z. Comparative genomics of the herbivore gut symbiont Lactobacillus reuteri reveals genetic diversity and lifestyle adaptation. Front. Microbiol. 2018, 9, 1151. [Google Scholar] [CrossRef]
- Maldonado, N.C.; de Ruiz, C.S.; Otero, M.C.; Sesma, F.; Nader-Macías, M.E. Lactic acid bacteria isolated from young calves–characterization and potential as probiotics. Res. Vet. Sci. 2012, 92, 342–349. [Google Scholar] [CrossRef]
- Brashears, M.; Jaroni, D.; Trimble, J. Isolation, selection, and characterization of lactic acid bacteria for a competitive exclusion product to reduce shedding of Escherichia coli O157: H7 in cattle. J. Food Prot. 2003, 66, 355–363. [Google Scholar] [CrossRef]
- Lin, W.-C.; Ptak, C.P.; Chang, C.-Y.; Ian, M.-K.; Chia, M.-Y.; Chen, T.-H.; Kuo, C.-J. Autochthonous lactic acid bacteria isolated from dairy cow feces exhibiting promising probiotic properties and in vitro antibacterial activity against foodborne pathogens in cattle. Front. Vet. Sci. 2020, 7, 239. [Google Scholar] [CrossRef]
- Mu, Q.; Tavella, V.J.; Luo, X.M. Role of Lactobacillus reuteri in human health and diseases. Front. Microbiol. 2018, 9, 757. [Google Scholar] [CrossRef]
- Garg, K.B.; Ganguli, I.; Das, R.; Talwar, G. Spectrum of Lactobacillus species present in healthy vagina of Indian women. Indian J. Med. Res. 2009, 129, 652–657. [Google Scholar] [PubMed]
- Kumar, S.; Varada, V.V.; Banakar, P.S.; Tyagi, N.; Chouraddi, R.; Hogarehalli Mallapa, R.; Tyagi, A.K. Screening and characterization of Sahiwal cattle calves-origin lactic acid bacteria based on desired probiotic attributes for potential application. Anim. Biotechnol. 2022, 34, 1612–1625. [Google Scholar] [CrossRef]
- Lähteinen, T.; Malinen, E.; Koort, J.M.; Mertaniemi-Hannus, U.; Hankimo, T.; Karikoski, N.; Pakkanen, S.; Laine, H.; Sillanpää, H.; Söderholm, H. Probiotic properties of Lactobacillus isolates originating from porcine intestine and feces. Anaerobe 2010, 16, 293–300. [Google Scholar] [CrossRef]
- Mishra, S.K.; Malik, R.; Manju, G.; Pandey, N.; Singroha, G.; Behare, P.; Kaushik, J. Characterization of a reuterin-producing Lactobacillus reuteri BPL-36 strain isolated from human infant fecal sample. Probiot. Antimicrob. Proteins 2012, 4, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Reuben, R.C.; Roy, P.C.; Sarkar, S.L.; Alam, R.-U.; Jahid, I.K. Isolation, characterization, and assessment of lactic acid bacteria toward their selection as poultry probiotics. BMC Microbiol. 2019, 19, 253. [Google Scholar] [CrossRef] [PubMed]
- Dudriková, M. Antimicrobially active lactobacilli from goats’ milk that do not produce biogenic amines. J. Food Nutr. Res. 2015, 54, 270–274. [Google Scholar]
- Kološta, M.; Slottová, A.; Drončovský, M.; Klapáčová, L.; Kmeť, V.; Bujňáková, D.; Lauková, A.; Greif, G.; Greifová, M.; Tomáška, M. Characterisation of lactobacilli from ewe’s and goat’s milk for their further processing re-utilisation. Slovak J. Food Sci. 2014, 8, 130–134. [Google Scholar]
- Reuben, R.C.; Elghandour, M.M.; Alqaisi, O.; Cone, J.W.; Márquez, O.; Salem, A.Z. Influence of microbial probiotics on ruminant health and nutrition: Sources, mode of action and implications. J. Sci. Food Agric. 2022, 102, 1319–1340. [Google Scholar] [CrossRef]
- Mikelsaar, M.; Zilmer, M. Lactobacillus fermentum ME-3–an antimicrobial and antioxidative probiotic. Microb. Ecol. Health Dis. 2009, 21, 1–27. [Google Scholar]
- Ramos, C.L.; Thorsen, L.; Schwan, R.F.; Jespersen, L. Strain-specific probiotics properties of Lactobacillus fermentum, Lactobacillus plantarum and Lactobacillus brevis isolates from Brazilian food products. Food Microbiol. 2013, 36, 22–29. [Google Scholar] [CrossRef]
- Naghmouchi, K.; Belguesmia, Y.; Bendali, F.; Spano, G.; Seal, B.S.; Drider, D. Lactobacillus fermentum: A bacterial species with potential for food preservation and biomedical applications. Crit. Rev. Food Sci. Nutr. 2020, 60, 3387–3399. [Google Scholar] [CrossRef] [PubMed]
- Olivares, M.; Díaz-Ropero, M.P.; Sierra, S.; Lara-Villoslada, F.; Fonollá, J.; Navas, M.; Rodríguez, J.M.; Xaus, J. Oral intake of Lactobacillus fermentum CECT5716 enhances the effects of influenza vaccination. Nutrition 2007, 23, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.-H.; Yu, B.; Jang, S.-H.; Tsen, H.-Y. Different probiotic properties for Lactobacillus fermentum strains isolated from swine and poultry. Anaerobe 2007, 13, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Mohammadabadi, T.; Bakhtiari, M.A.; Alimirzaei, P. Isolation and identification of lactate-producing and utilizing bacteria from the rumen of Najdi goats. Indian J. Small Rumin. 2018, 24, 276–280. [Google Scholar] [CrossRef]
- Ruiz, P.; Seseña, S.; Palop, M.L. Characterization of bacterial populations from Murciano-Granadina goat colostrum. Dairy Sci. Technol. 2014, 94, 549–560. [Google Scholar] [CrossRef]
- Islam, M.Z.; Uddin, M.E.; Rahman, M.T.; Islam, M.A.; Harun-ur-Rashid, M. Isolation and characterization of dominant lactic acid bacteria from raw goat milk: Assessment of probiotic potential and technological properties. Small Rumin. Res. 2021, 205, 106532. [Google Scholar] [CrossRef]
- Ayyanna, R.; Ankaiah, D.; Arul, V. Anti-inflammatory and antioxidant properties of probiotic bacterium Lactobacillus mucosae AN1 and Lactobacillus fermentum SNR1 in Wistar albino rats. Front. Microbiol. 2018, 9, 3063. [Google Scholar] [CrossRef]
- Rastogi, S.; Mittal, V.; Singh, A. In vitro evaluation of probiotic potential and safety assessment of Lactobacillus mucosae strains isolated from donkey’s lactation. Probiot. Antimicrob. Proteins 2020, 12, 1045–1056. [Google Scholar] [CrossRef]
- de Moraes, G.M.D.; de Abreu, L.R.; do Egito, A.S.; Salles, H.O.; da Silva, L.M.F.; Nero, L.A.; Todorov, S.D.; Dos Santos, K.M.O. Functional properties of Lactobacillus mucosae strains isolated from Brazilian goat milk. Probiot. Antimicrob. Proteins 2017, 9, 235–245. [Google Scholar] [CrossRef]
- Angelakis, E.; Bastelica, D.; Amara, A.B.; El Filali, A.; Dutour, A.; Mege, J.-L.; Alessi, M.-C.; Raoult, D. An evaluation of the effects of Lactobacillus ingluviei on body weight, the intestinal microbiome and metabolism in mice. Microb. Pathog. 2012, 52, 61–68. [Google Scholar] [CrossRef]
- Huang, C.-H.; Shen, C.-C.; Liang, Y.-C.; Jan, T.-R. The probiotic activity of Lactobacillus murinus against food allergy. J. Funct. Foods 2016, 25, 231–241. [Google Scholar] [CrossRef]
- Sarkar, S.L.; Hossain, M.I.; Monika, S.A.; Sanyal, S.K.; Roy, P.C.; Hossain, M.A.; Jahid, I.K. Probiotic potential of Pediococcus acidilactici and Enterococcus faecium isolated from indigenous yogurt and raw goat milk. Microbiol. Biotechnol. Lett. 2020, 48, 276–286. [Google Scholar] [CrossRef]
- Tserovska, L.; Stefanova, S.; Yordanova, T. Identification of Lactic Acid Bacteria Isolated from Katyk, Goat’s Milk and Cheese; National Bank for Industrial Microorganisms and Cell Cultures: Sofia, Bulgaria, 2002. [Google Scholar]
- Makete, G.; Aiyegoro, O.A.; Thantsha, M.S. Isolation, identification and screening of potential probiotic bacteria in milk from south african saanen goats. Probiot. Antimicrob. Proteins 2017, 9, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Fugaban, J.I.I.; Vazquez Bucheli, J.E.; Park, Y.J.; Suh, D.H.; Jung, E.S.; Franco, B.D.G.d.M.; Ivanova, I.V.; Holzapfel, W.H.; Todorov, S.D. Antimicrobial properties of Pediococcus acidilactici and Pediococcus pentosaceus isolated from silage. J. Appl. Microbiol. 2022, 132, 311–330. [Google Scholar] [CrossRef]
- Ladha, G.; Jeevaratnam, K. Probiotic potential of Pediococcus pentosaceus LJR1, a bacteriocinogenic strain isolated from rumen liquor of goat (Capra aegagrus hircus). Food Biotechnol. 2018, 32, 60–77. [Google Scholar] [CrossRef]
- Standen, B.; Rawling, M.; Davies, S.; Castex, M.; Foey, A.; Gioacchini, G.; Carnevali, O.; Merrifield, D. Probiotic Pediococcus acidilactici modulates both localised intestinal-and peripheral-immunity in tilapia (Oreochromis niloticus). Fish Shellfish. Immunol. 2013, 35, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, T.; Ikari, N.; Kouchi, T.; Kowatari, Y.; Kubota, Y.; Shimojo, N.; Tsuji, N.M. The molecular mechanism for activating IgA production by Pediococcus acidilactici K15 and the clinical impact in a randomized trial. Sci. Rep. 2018, 8, 5065. [Google Scholar] [CrossRef]
- Mikulski, D.; Jankowski, J.; Naczmanski, J.; Mikulska, M.; Demey, V. Effects of dietary probiotic (Pediococcus acidilactici) supplementation on performance, nutrient digestibility, egg traits, egg yolk cholesterol, and fatty acid profile in laying hens. Poult. Sci. 2012, 91, 2691–2700. [Google Scholar] [CrossRef]
- Porto, M.C.W.; Kuniyoshi, T.M.; Azevedo, P.; Vitolo, M.; Oliveira, R.S. Pediococcus spp.: An important genus of lactic acid bacteria and pediocin producers. Biotechnol. Adv. 2017, 35, 361–374. [Google Scholar] [CrossRef]
- Švec, P.; Franz, C.M. The genus Enterococcus. In Lactic Acid Bacteria: Biodiversity and Taxonomy; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2014; pp. 175–211. [Google Scholar]
- Öztürk, H.; Geniş, B.; Özden Tuncer, B.; Tuncer, Y. Bacteriocin production and technological properties of Enterococcus mundtii and Enterococcus faecium strains isolated from sheep and goat colostrum. Vet. Res. Commun. 2023. [Google Scholar] [CrossRef]
- Rajput, K.; Dubey, R.C.; Kumar, A. Probiotic potential and immunomodulatory properties in Enterococcus faecium GMB24 and Enterococcus hirae SMB16 isolated from goat and sheep milk. Arch. Microbiol. 2022, 204, 619. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LAB spp. | 2 Days (35) | 7 Days (28) | 14 Days (28) | 28 Days (31) | 56 Days (34) | 0 DPW (28) | 2 DPW (32) | 7 DPW (31) | 1 MPW (32) | 4 MPW (30) | 8 MPW (30) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| L. reuteri | 30 | 15 | 21 | 21 | 34 | 25 | 27 | 18 | - | - | 3 |
| L. fermentum | 9 | 4 | 7 | 3 | 3 | - | - | - | - | - | - |
| L. johnsonni | 7 | 6 | 2 | 3 | 5 | 3 | 1 | 1 | 3 | - | 6 |
| L. murinus | 3 | - | 4 | 1 | - | - | - | 2 | - | - | - |
| L. salivarius | - | 1 | - | 1 | - | - | 1 | 3 | - | - | - |
| L. plantarum | 1 | 1 | 2 | 1 | - | - | 1 | - | - | 10 | 2 |
| L. brevis | - | 1 | - | - | - | - | - | - | - | - | 1 |
| L. oris | - | - | - | 1 | - | - | - | - | - | - | - |
| L. acidophilus/amyolovorus | - | - | - | - | 1 | - | 2 | 1 | - | - | - |
| Pediococcus spp. | 26 | 16 | 22 | 20 | 15 | 2 | 3 | 9 | 8 | 3 | 1 |
| L. mucosae | - | - | - | - | - | 12 | 14 | 25 | 32 | 29 | 28 |
| L. animalis | - | - | - | - | - | - | - | - | - | 1 | 10 |
| Enterococcus spp. | - | - | - | - | - | - | - | - | - | 6 | 12 |
| L. agilis | - | - | - | - | - | 2 | - | 2 | - | - | - |
| Weisella confusa, W. cibaria | - | - | - | - | - | - | - | - | - | - | 2 |
| L. saerimneri | - | - | - | - | - | - | 1 | 1 | - | - | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
ElHadedy, D.E.; Kim, C.; Yousuf, A.B.; Wang, Z.; Ndegwa, E.N. Understanding Age-Related Longitudinal Dynamics in Abundance and Diversity of Dominant Culturable Gut Lactic Acid Bacteria in Pastured Goats. Animals 2023, 13, 2669. https://doi.org/10.3390/ani13162669
ElHadedy DE, Kim C, Yousuf AB, Wang Z, Ndegwa EN. Understanding Age-Related Longitudinal Dynamics in Abundance and Diversity of Dominant Culturable Gut Lactic Acid Bacteria in Pastured Goats. Animals. 2023; 13(16):2669. https://doi.org/10.3390/ani13162669
Chicago/Turabian StyleElHadedy, Doaa E., Chyer Kim, Adnan B. Yousuf, Zhenping Wang, and Eunice N. Ndegwa. 2023. "Understanding Age-Related Longitudinal Dynamics in Abundance and Diversity of Dominant Culturable Gut Lactic Acid Bacteria in Pastured Goats" Animals 13, no. 16: 2669. https://doi.org/10.3390/ani13162669