
Black Soldier Fly Larva Oil in Diets with Roughage to Concentrate Ratios on Fermentation Characteristics, Degradability, and Methane Generation


Tropical Feed Resources Research and Development Center (TROFREC), Department of Animal Science, Faculty of Agriculture, Khon Kaen University, Khon Kaen 40002, Thailand
*
Author to whom correspondence should be addressed.
Animals 2023, 13(15), 2416; https://doi.org/10.3390/ani13152416
Submission received: 25 June 2023
/
Revised: 19 July 2023
/
Accepted: 24 July 2023
/
Published: 26 July 2023
(This article belongs to the Section Animal Nutrition)
Abstract
:Simple Summary
Animal husbandry costs are being driven up by a scarcity of high-quality, expensive animal feed. Researchers are investigating alternative feed resources derived from black soldier fly larva, including the utilization of oil from black fly larvae as a byproduct of the industry. A study using in vitro gas production techniques investigated the influence of black soldier fly larva oils and the proportion of roughage-to-concentrate ratios on in vitro fermentation. Results showed that adding 4% black soldier fly larva oils (BSFO) in concentration at different roughage (R)-to-concentrate (C) ratios increased propionate levels, decreased methane (CH4) emission, and preserved dry matter (DM) degradability.
Abstract
Currently, the scarcity of high-quality, expensive animal feed is a primary factor driving up the cost of animal husbandry. As a result, most researchers have focused on improving the potential of using alternative feed resources derived from the black soldier fly larva. In particular, the utilization of oil from black fly larvae is a byproduct of the industry. The aim of this study was to investigate the influence of black soldier fly larva oils and the proportion of roughage-to-concentrate ratios on gas kinetics, rumen characteristics, degradability, and mitigate CH4 production by using in vitro gas production techniques. The in vitro investigation used a completely randomized design (CRD) with a 2 × 4 factorial arrangement. The level of R:C ratios (60:40 and 40:60) were factor A, while BSFO levels (0, 2, 4, and 6% of DM) were factor B. Under this investigation, the combined impact of R:C ratio and BSFO on the kinetics of gas and accumulative gas production was found to be significant (p < 0.01). After 4 h of incubation, the pH and ammonia-nitrogen (NH3-N) concentration were found to be impacted by the inclusion of BSFO levels at different R:C-ratios (p < 0.01). Moreover, after 4 and 8 h of incubation, supplementing the BSFO at 4% with the level of R:C ratio at 40:60 resulted in a significant reduction in the amount of CH4 in the rumen (p < 0.05). However, the inclusion of BSFO levels at different R:C ratios had no effect on the degradability of DM after 12 and 24 h of incubation (p > 0.05), whereas increasing the concentration of BSFO in concentrate at 6% reduced the DM degradability after 24 h of incubation (p < 0.05). Furthermore, adding BSFO to the diet at various R:C ratios enhanced the propionate (C3) concentration, with the highest level observed with the level of R:C ratio at 40:60 and 4% BSFO inclusion (p < 0.05). To summarize, the addition of BSFO at 4% with a 40:60 of R:C ratio increased C3 levels, decreased CH4 emission, and preserved DM degradability. A R:C ratio of 40:60 could improve the total volatile fatty acids and digestibility. Moreover, the inclusion of 6% BSFO at different R:C ratios lowered the in vitro dry matter digestibility, in vitro organic matter digestibility, NH3-N, and protozoal populations.
1. Introduction
A greenhouse gas (GHG) released by livestock is methane (CH4), which accounts for 19% of all emissions [1], 45% of which are related to intestinal fermentation [2]. Methanogenic archaea use hydrogen present in the rumen to reduce CO2 to CH4 through methanogenesis; this causes the gross energy (GE) absorbed in the meal to decrease by 2 to 12% [3]. The rumen’s anaerobic breakdown of digested organic compound (OM) is the primary pathway for producing CH4 as the end product. Thus, the goals of CH4 mitigation measures in ruminants have been to gain economic and anti-global warming benefits [4].
Several methods have been attempted to reduce emitting CH4 in animals, such as defaunation, the use of chemical inhibitors, and the use of ionophores, which can either indirectly or directly suppress methanogenesis in the rumen; however, no consistently effective approaches have been identified for practical use. One of the alternative interesting topics is the addition of lipids in the diets of cattle. Typically, lipid supplementation is utilized to boost the ration’s energy density [5]. There is significant proof that enteric CH4 production is reduced by additional oil-rich meals [6]. By lowering fermentation and CH4 production, switching ruminant diets from carbs to lipids has been demonstrated to have an effect on gas generation in the rumen [7,8]. Studies have shown that adding 2 to 6% of lipids to the diets of ruminants can lead to reductions in CH4 emissions ranging from 7.2% to 21.4% [9,10]. Numerous causes, including the substitution of rumen fermentable organic compounds in feed, a decline in ruminal protozoa and methanogen, and the biohydrogenation of unsaturated fatty acids, are credited with this decrease [11].
Oil is a substance that is primarily abundant in insects. For ruminants, insects have lately been proposed as a sufficient, sustainable, and novel source of protein and fat [12], with no detrimental effects on rumen characteristics or animal performance [13]. It has been discovered that the concentration of ether extract in black soldier fly larvae oils (BSFO) ranges from 15.0 to 34.8% dry matter [14]. Several writers have investigated and reported on the fatty acid composition of oils derived from BSFO [15,16]. To our knowledge, there is currently no research available on the influence of insect oils on rumen properties, degradability, and the reduction of CH4 production.
The purpose of the present study was to investigate the effect of black soldier fly larvae oils and the roughage-to-concentrate ratio on gas patterns, rumen characteristics, degradability, and CH4 reduction utilizing in vitro gas production techniques.
2. Materials and Methods
2.1. Black Soldier Fly Larvae Oil Preparation and Design
To maintain the nutritional and chemical quality, the clean and dried black soldier fly larvae (BSF) were subjected to press-defatting at 45 °C to 48 °C using an NF-80 cold press (Karaerler, Ankara, Turkey), and the process was carried out by BSFLY Company Ltd., Udonthani, Thailand.
The current experiment was conducted using a gas generation method over a range of incubation times. Using a completely randomized design with three replication runs (CRD), a 2 × 4 factorial experiment was conducted. The experimental diets consisted of two different roughage-to-concentrate (R:C) ratios (60:40 and 40:60) and four different levels of BSFO concentration (0, 2, 4, and 6% DM). As a roughage source, rice straw was used. Table 1 lists the components and nutrients found in both the concentrates and rice straw.
2.2. Animals and Rumen Fluid Inoculum
For rumen fluid supply, Thai native steers (2.0–2.3 years old) weighing 350 ± 10 kg were employed. For 14 days, at 7:30 and 15:30, the cattle were fed with concentrate mixture (14.0% crude protein (CP) and 75% total digestible nutrient) at 1% of their weight and rice straw was fed on an ad libitum basis. Each animal was housed in a separate corral and had unrestricted access to fresh, clean water as well as mineral blocks. Before morning feeding, rumen fluids from each cow were also collected using a pump connected to a stomach tube. The fluids were transferred to thermos flasks that had been preheated and had an O2-free headspace before being transported anaerobically to the lab at 39 °C and used as inoculum after passing through four layers of cheesecloth.
2.3. Fermentation Substrates In Vitro
Artificial saliva was prepared on the day of conduction, prior to the collection of rumen fluids. A two-set inoculum mixture in thermos flasks containing artificial saliva and rumen fluid at a ratio of 2:1 was performed. The rice straw and concentration samples, weighing 0.5 g each, were placed into 50 mL serum vials. Four replications with five blanks were created for each treatment. For the in vitro gas generation test, the bottles were closed with rubber disclosures and crimp caps, which were then incubated for 96 h at 39 °C. Every three hours, the bottles were gently shaken during the incubation phase. Three 50 mL glass bottles, one in quadruplicate and one in blank, each holding 0.5 g of substrate on a dry matter basis, were set up, flushed with carbon dioxide, and preheated to 39 °C. The bottles were divided into three sets: the first set (4 bottles × 8 treatments × 4 bottles of blank) was used for gas kinetics and gas production measurement (incubated for 96 h), the second set (4 bottles × 8 treatments × 2 observation times at 4 and 8 h) for the measurement of ruminal parameters including pH, ruminal ammonia-nitrogen (NH3-N), volatile fatty acids, and protozoa, and the third set (4 bottles × 8 treatments × 2 observation times at 12 and 24 h × 4 bottles of blank) was used for the determination of nutrient digestibility.
2.4. Measurements and Chemical Analysis
The samples of rice straw and concentrate were put through a 1-mm screen on a Cyclotech Mill from Tecator in Hoganas, Sweden, dried for 72 h at 60 °C, then analyzed in vitro for chemical composition and gas generation. According to the AOAC International Method [17], the following quantities were measured and conveyed with any leftover ash included: dry matter (DM), ether extract (EE), organic matter (OM), and acid detergent fiber (ADF). The α-amylase and sodium sulfite and were used to compute the neutral detergent fiber (NDF) in accordance with the methodology of Van Soest et al. [18] (Sigma no. A3306, Sigma Chemical Co., St. Louis, MO, USA). According to the AOAC [17], the amount of CP in the feed samples was measured using a Leco combustion nitrogen analyzer (Leco CN628 Carbon/Nitrogen Analyzer, Leco Instruments Inc., St. Joseph, MI, USA). The fatty acid content of BSFO was assessed using gas chromatography with a flame ionization detector (GC-FID) (Thermo Scientific Trace GC Ultra) and a WCOT fused silica column (100 m, 0.25 mm i.d., 0.2 m f.t., Coating Select Fame, Varian, Houten, The Netherlands). Table 1 shows the results as a percentage of total fatty acids. A pressure transducer and calibrated syringe were used at many moments in time (0.5, 1, 2, 4, 6, 8, 12, 18, 24, 36, 48, 72, and 96 h) to assess gas generation right after incubation. After inserting a fitted syringe with needle No. 24 into the rubber stopper of fermentation bottles, the bottles were placed in an incubator. The pressure transducer’s metering system was made up of a 3-way stopcock valve, a mechanical pressure gauge, a glass syringe, and a needle. The pressure within the serum bottles was monitored through the transducer linked to the 3-way valve, while the amount of gas produced was measured with the glass syringe. The third port of the valve was connected to a hose with a needle for puncturing the butyl rubber stopper plug on the injection bottle.
The pH levels of 64 bottles of fermented inoculum were determined at 4 and 8 h after inoculation using 32 samples taken at each time. The liquid was then filtered through Grade 40 Cheesecloth. The frozen rumen fluid was thawed and centrifuged at 16,000 g for 10 min to determine NH3-N and the volatile fatty acid contents (VFAs). The VFA proportion was determined using gas chromatography, as outlined in the approach by So et al. [19] using an altered technique of Osaki et al. [20]. According to the Fawcett and Scott method [21], 40 microliters of centrifuged rumen liquid, 2500 microliters of phenol color reagent, and 2000 microliters of alkaline hypochloride reagent were combined in a 15 mL test tube using a vortex mixer. A blue reaction was produced when the mixture was vortexed and then incubated for 10 min at 37 °C in a shaking water bath. A second 630 nm UV/Vis spectrophotometric analysis of the mixture followed. To assess the rumen’s microorganisms, a 1 mL sample of the serum was mixed thoroughly with 9 mL of formaldehyde after being incubated for 4 and 8 h. Ruminal protozoa was quantified using a hemocytometer (Boeco, Hamburg, Germany) and a manual counting technique [22]. A leak-proof syringe was used to extract 10 mL of gas from the fermented bottle’s headspace, and gas chromatography was used to determine the amount of CH4 in the sample at 4 and 8 h after incubation (Shimadzu Corporation, Kyoto, Japan).
After filtering the contents through a pre-weighed glass filter crucible of 50 mL, the material was incubated for 12 and 24 h (4 samples per treatment) before determining its in vitro dry matter digestibility (IVDMD) [23]. The NDF concentration of the indigestible residues in the test bottles was determined in order to compute the in vitro true digestibility (IVTD) [18] using the following equations.
where NDF = neutral detergent fiber (% of DM), IVTD = in vitro true digestibility (% of DM), and NDFD = neutral detergent fiber digestibility (% of NDF).
IVTD = 100 − [(100 − NDFD) × (NDF/100)],
To determine the amount of organic matter (OM) and the percentage of OM digestibility (IVOMD), the glass filter crucible was heated at 550 °C for 6 h. The residue data were then used to make these determinations.
2.5. Statistical Analysis
The cumulative gas production curve was calculated using the Schofield [24] model.
where b is the final asymptotic gas volume corresponding to fully digested substrate (mL/g DM), t is the incubation time (h), c is a rate constant (units time 1), and L is a discontinuous lag term (h).
Gas production = b × [1 − exp−c(t − L)],
The PROC GLM of SAS [25] was used to evaluate the in vitro study’s data in accordance with a 2 × 4 factorial in a completely randomized system. The model is:
where Yijk are the response variances; µ is the overall mean, ai are the R:C ratio levels at 60:40 and 40:60 (i, 1–2), bj are the BSFO levels at 0, 2, 4, and 6% of DM (j, 1–4), abij are the interaction effects, and εijk are the residues. The means of the response variances were also presented, along with the standard error of the mean. Mean values of each individual run were used as the experimental unit. With a significance level of p < 0.05, the least significant difference (lsd) was utilized to examine statistical differences between treatment means.
Yijk = µ + ai + bj + abij + εijk,
3. Result
3.1. Nutritional Composition of Diet
Table 1 lists the ingredients and chemical composition of concentrate diet and rice straw. The protein level of the concentrate diets, which ranged from 15.05 to 15.45% DM in each group, was almost the same, and urea was given to modify the CP content, and rice straw was used as a source of roughage in a substrate that contained CP at a concentration of 2.54% DM. The EE contents of the inclusion of 0, 2, 4, and 6% BSFO in the experimental diets were 2.05, 4.02, 6.03, and 7.96, respectively, which progressively increase when BSFO is added from 0 to 6%. Additionally, adding 0, 2, 4, and 6% BSFO to the feed increased the amount of GE, from 16.98 to 17.67, favoring gross energy density.
3.2. Kinetics of Gas Production
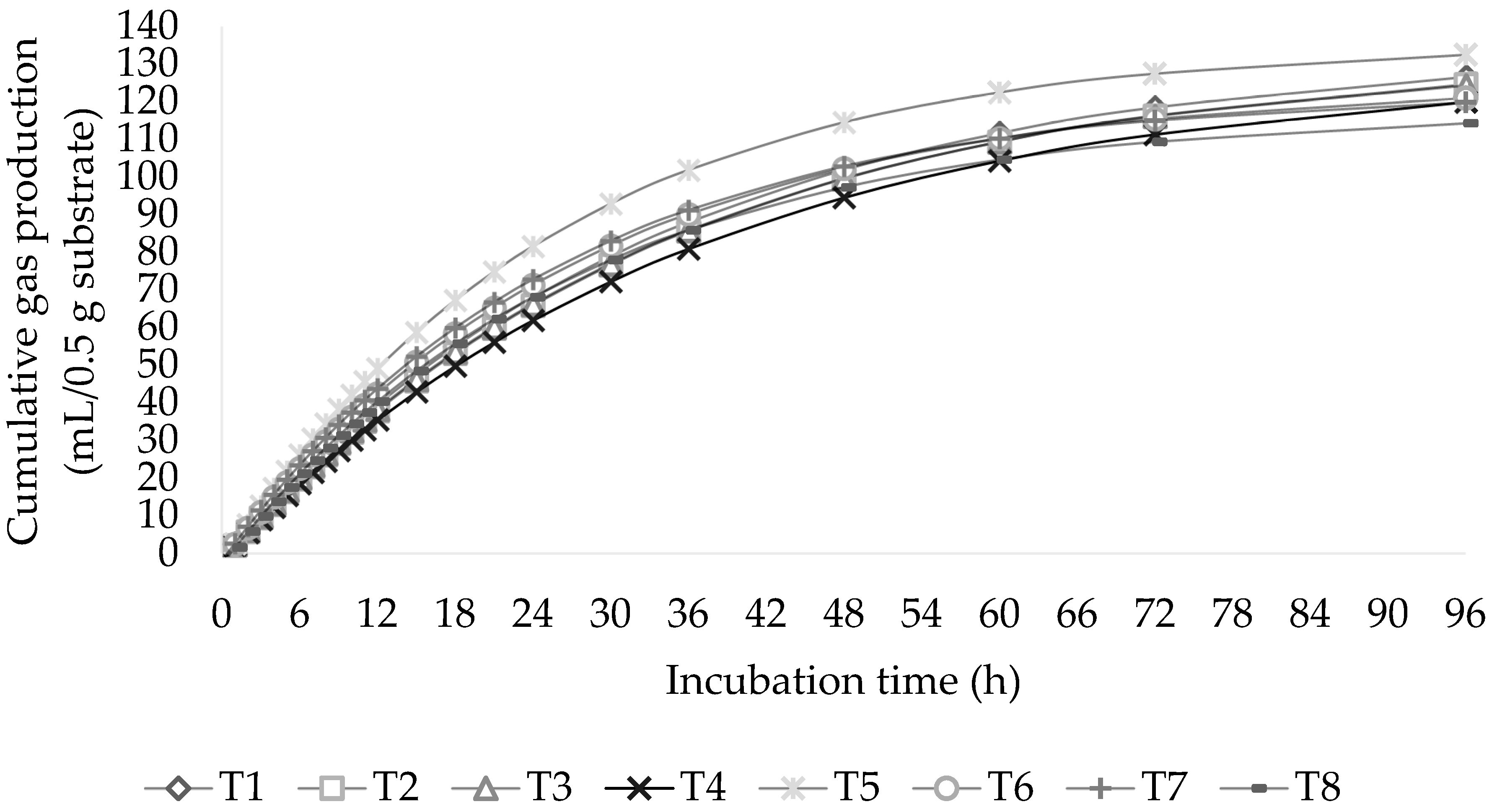
The calculated parameters for the tested substrates based on the kinetics of producing gases models are shown in Table 2. At 96 h of incubation, a relationship among the R:C ratio and increasing the BSFO for asymptotic gas production (b) and gas accumulation was observed (Figure 1; p < 0.01). According to the findings, the asymptotic gas production and overall gas production were at their peak in the R:C ratio (40:60) with BSFO included at 0%. The rate of gas generation (c) and the discrete lag time (L) before gas production did not interact at the R:C ratio and BSFO levels (p > 0.05). However, the discrete lag time was reduced (p < 0.01) when a significant amount of concentrate diet was administered.
3.3. In Vitro Ruminal Fermentation and CH4 Concentration
Table 3 illustrates the influence of R:C ratio and BSFO level on NH3-N concentration, pH, CH4 output, and protozoal population after 4 and 8 h of incubation using an in vitro gas technique assay. After 4 h of incubation, there was a significant difference detected (p < 0.01) among the inclusion of BSFO levels at different R:C-ratios for pH and NH3-N concentration, and the values ranged from 6.95 to 7.03 and 18.42 to 22.41 mg/dL, respectively. The inclusion of BSFO levels at different R:C ratios did not have an effect on NH3-N and pH during 8 h of incubation (p > 0.05). However, the level of R:C ratio at 40:60 dramatically reduced pH, whereas increasing the amount of BSFO in the diet up to 2% significantly decreased the impact (p < 0.01) on ruminal NH3-N, with the lowest values of 17.28 mg/dL for the inclusion of BSFO at 6%. The concentration of CH4 was significantly reduced by enhancing doses of BSFO in various roughage-to-concentrate ratios. The amount of CH4 was at its lowest (2.45 mL/1 g) at 4 h after incubation when the level of R:C ratio at 60:40 in 4% BSFO inclusion (p < 0.01), and at its lowest (6.30 mL/1 g) at 8 h after incubation when the level of R:C ratio was at 60:40 in 4% BSFO inclusion (p < 0.05). For protozoal populations at 4 and 8 h after incubation, there was no relationship between the level of R:C ratio and BSFO inclusion (p > 0.05). Nonetheless, adding up to 2% BSFO considerably reduced the impact (p < 0.01) on protozoal numbers at 4 and 8 h, with the lowest values being 2.50 × 106 and 3.00 × 106 cells/mL, respectively.
3.4. In Vitro Degradability
Table 4 lists the impact of the roughage-to-concentrate ratio and the BSFO level on the in vitro digestibility of DM, OM, and IVTD. The in vitro digestibility was unaffected by the level of R:C ratio and BSFO inclusion (p > 0.05), except that IVOMD at 12 h and IVTD at 24 h after incubation have the lowest values at 36.42% and 76.51% DM, respectively, when the level of R:C ratio was 60:40 in BSFO inclusion at 6% (p < 0.05). The IVDMD, IVOMD, and IVTD at various hours of incubation times were increased when a high quantity of concentrate diet was provided (p < 0.05). In addition, the presence of BSFO at 6% decreased the values of the in vitro digestibility of DM and OM at 24 h, which were 39.53 and 46.70% DM, respectively (p < 0.01).
3.5. Ruminal Volatile Fatty Acid Concentration
Table 5 illustrates the influence of the R:C ratio and BSFO on total VFAs, acetate (C2), propionate (C3), and butyrate (C4) concentrations. No changes (p > 0.05) existed between the inclusion of BSFO levels at different R:C ratios on total VFAs and profiles at 4 h. Nevertheless, increasing the amount of concentrate diet from 40% to 60% enhanced total VFAs by 61.08 mmol/L (p < 0.01). After 8 h of incubation, the level of R:C ratio at 40:60 caused a decrease of C2 in rumen fluids of approximately 2.28% when compared with the level of R:C ratio at 60:40, whereas increasing the amount of BSFO in the diet up to 2% significantly decreased the effect (p < 0.01) on the concentration of C2 with the highest values of 68.33 and 68.85% for inclusion of BSFO at 4 and 6%, respectively. At 8 h, there were relationships (p < 0.05) among the level of R:C ratio and BSFO on C3 and C4. The level of R:C ratio at 40:60 in BSFO inclusion at 4 and 6% shows the highest C3 at approximately 21.10 and 20.67%, respectively. In addition, the C4 concentration was lowest (11.47%) when the level of R:C ratio at 40:60 with 2% BSFO inclusion (p < 0.05).
4. Discussion
4.1. Production of Gas and Kinetics
There is evidence that the inclusion of BSFO in a ruminants diet affects the kinetics of gas in the rumen [26]. The addition of oil tends to decrease the gas generation rate and increase the duration of the lag time, or the time it takes for gas to start being produced after feeding. In this study, higher amounts of BSFO in the various R:C ratio levels reduced asymptotic gas production and total gas. This might be a result of the concentrate diet’s high oil content, which slows the rate of feed fermentation in the rumen and reduces gas output. Oil can also affect the physical properties of the rumen, such as the viscosity and flow rate, which can influence the rate and extent of digestion [27]. The combination of garlic powder and coconut oil in in vitro studies, according to Kongmun et al. [28], results in gas generation. In comparison to the control group, adding coconut oil at various doses reduced gas accumulation. Similarly, Kang et al. [29] discovered that using krabok seed oil as a rumen stimulant in a gas production system significantly reduced gas values and kinetics.
4.2. In Vitro Ruminal Fermentation and CH4 Concentration
Ruminal pH is an important metric because it indicates the internal balance of the rumen ecosystem. Ruminants typically maintain well-balanced conditions in order to maintain a ruminal pH ranging from 6.5 to 7 [30]. The current investigation found that, when concentrate intake increased, the ruminal pH dropped. The rumen’s pH was found to be at its lowest when the level of R:C ratio was 40:60 and BSFO was excluded. The observed decrease in pH could be attributed to an elevated concentration of fermentable carbohydrates in the concentrate diet, which undergo rapid fermentation. According to Ramos et al. [31], consuming feed high in concentrated substances caused the ruminal pH to significantly drop, the rate of metabolism to slow down, and the activity of cellulolytic bacteria to decrease. Subacute ruminal acidosis (SARA) and the control of bacterial growth are both influenced by the daily variation of ruminal pH. The level of pH in the rumen shows that stable pH is necessary for healthy rumen ecology, metabolism, and microbial development.
Microbial protein synthesis is mostly nitrogen-dependent, and ammonia can be the primary supply of nitrogen for bacterial development. At 4 h after incubation, the NH3-N content in the present study varied between 18.42 and 22.41 mg/dL. The maximum NH3-N concentration was reported when the diet contained 0% to 4% BSFO in the level of R:C ratio at 40:60. When compared to a low-concentrate diet, a high-concentration feed ratio with a higher CP may result in higher amounts of NH3-N due to increased microbial breakdown. Maintaining an NH3-N concentration in the 15–30 mg/dL range can improve rumen ecology in areas such as feed intake, bacterial protein production, and digestibility [32]. In contrast, a lack of NH3-N can slow bacterial development. The high content of ruminal NH3-N in the high-concentrate diet is explained by a dynamic balance among NH3-N production and consumption by rumen bacteria [33,34]. The observation of large amounts of NH3-N in the stomach of ruminants indicates that the ruminants diet contains an adequate amount of accessible nitrogen, which is most likely due to the diet’s high concentrate concentration. According to Musco et al. [33], the higher concentration of ruminal NH3-N in the high-concentrate diet could also be due to the higher presence of rumen-degradable protein. In addition, the findings imply that increasing the amount of oil in the diet may reduce NH3-N concentrations in the rumen, probably due to a decrease in protozoa populations. This finding is consistent with a prior study that found that adding plant oil reduced rumen ammonia concentrations. Ammonia is typically formed by the degradation of bacterial proteins, which is frequently related to the presence of protozoa. Several studies show that using essential oils can reduce ammonia concentrations in in vitro systems. According to Castillejos et al. [35], essential oils can interact with the bacterial cell membrane, preventing the growth of specific strains and ultimately resulting in a drop in ammonia concentration [36].
The amount and type of dietary lipids, as well as the mix of the diet, all influence the changes that take place in the rumen ecology [37]. The amount and type of dietary lipids, as well as the food mix, all influence the changes that occur in the rumen ecology, which explains why the lipid’s influence on the quantity and/or function of the microorganisms varies [11]. The current study’s protozoa population changed as a result of the inclusion of feeding BSFO. This is due to BSFO’s high lauric acid concentration, which has significant antiprotozoal characteristics and, as a result, has a variety of impacts on the development of fermentation end products and CH4 reduction in the rumen. Due to the impact of medium-chain fatty acids (MCFAs), prior study has revealed that a large amount of vegetable oil supplementation could decrease the rumen protozoal population in ruminants [11]. In addition, lauric (C12) and myristic (C14) acids in olive oil have a detrimental effect on protozoal membranes, which can reduce their fibrolytic activity, similar to what was found in Matsumoto et al. [38].
Reduced protozoa numbers are frequently accompanied by a decrease in CH4 generation as a result of increased dietary fat concentrations. In this experiment, increasing the BSFO inclusion reduced CH4 output. It has been demonstrated that adding oil or fat from a variety of sources can reduce the release of CH4 from cattle [11]. By decreasing the number or activity of methanogens, partially eliminating protozoa (which serve as the host for some methanogens), and reducing nutrient breakdown and the process of fermentation (which both promote hydrogen creation and, in the case of fatty acids that are unsaturated, act as substitute hydrogen sinks), oil can be used to inhibit methanogenesis [39]. The results of the study show that medium-chain fatty acids, long-chain fatty acids, saturated fatty acids, and unsaturated fatty acids are all effective at reducing the gastrocolic emissions of CH4 [40].
4.3. In Vitro Degradability
The digestibility of DM, OM, and true digestibility (TD) were enhanced as the percentage of concentrate was increased. This may be due to the fact that a concentrate diet might provide additional nutrients for the growth of rumen bacteria, thereby improving rumen fermentation. The improvement in DMD, OMD, and TD caused by a higher proportion of concentrate in the diet is a clear indication of concentrate components’ improved digestibility compared to treated straw [41]. These findings are comparable with those of Chen et al. [42], who found that cows fed a high concentrate diet had a considerable increase in nutritional digestibility. In contrast, Calabro et al. [43] compared several feeds and found higher OM in vitro digestibility as oil content increased. Moreover, oil supplementation in ruminant diets has been shown to have both positive and negative effects on rumen fermentation and digestion. In the current experiment, the digestibility of DM decreased with increasing BSFO levels. The reduction in the digestibility of fiber promoted by vegetable oil is attributed to two main effects: the physical effect of coating the feed particles and the toxic effect on the rumen microorganisms. These findings are consistent with those of Arcos-lvarez et al. [44], who found a decrease in digestible NDF intake at the two highest levels of olive oil inclusion (4 and 6%). Van-Cleef et al. [45] discovered that a 6% inclusion level of vegetable oils reduced the digestion of DM and fiber. Similarly, Sutton et al. [46] showed a 76% reduction in digestion in the forestomach and a 30% reduction in the overall tract when 40 g coconut oil was supplemented per sheep per day. However, in the present work, it seems that the slightly low in vitro DM and OM digestibilities might be attributed to the diet’s slightly higher fiber content, which may have limited digestibility [47]. Concentrate diets including NDF and ADF range from 19.9 to 22.48% and 12.37 to 14.33% DM, respectively, whereas rice straw contained 72% NDF and 50% ADF. Furthermore, fat addition may contribute to somewhat worse feed digestion.
4.4. Ruminal Volatile Fatty Acid Concentration
Increasing the quantity of concentrate in the diet from 40% to 60% resulted in a 13.21% increase in total volatile fatty acids. This could be owing to the presence of highly degradable carbohydrates, particularly starch, in the concentrate, which is consistent with the findings of Cherdthong et al. [48]. Furthermore, Phesatcha et al. [41] found that raising the concentrate ratio in the diet to 60% enhanced VFAs and C3, while decreasing C2 and the acetate to propionate (C2:C3) ratio.
Additionally, the propionate molar proportion constantly increased as BSFO increased, increasing the ratio of a concentrate diet to 60%. According to a previous study, adding fish oil raises the molar proportions of C3, C4, and isobutyric acids [11,49]. While our findings are largely compatible with these studies, the maximum concentration of C3 was reported with a 6% level of BSFO, which could be attributed to BSFO lipolysis-induced glycerol release in the rumen. Glycerol is quickly fermented and transformed into C3, accounting for 35% to 69% of its production [36,44]. Also, the C2:C3 ratio decreases as glycerol is converted to C3. The lipolysis process may also release unsaturated fatty acids, sometimes in excess of their biohydrogenation capacity, without affecting the fermentability of the rumen [37,45]. This causes a rise in the production of C3, which competes with metabolic hydrogen, a byproduct of fermentation, in the body. Similar to this, Mapato et al. [50] showed that supplementing with 6% sunflower oil decreased the ratio of C2:C3 while increasing the molar proportion of ruminal acetate and increasing the molar proportion of C3.
5. Conclusions and Recommendation
In conclusion, supplementation of BSFO at 4% at different R:C ratios enhanced C3 concentration and reduced CH4 production. An R:C ratio of 40:60 may be beneficial for digestibility and total volatile fatty acids; however, the R:C ratio of 60:40 was ineffective. In addition, the inclusion of 6% BSFO at different R:C ratios lowered the DMD, OMD, NH3-N, and protozoal populations. However, to examine how the R:C ratio and BSFO impact actual feeding regimens, and in vivo study is required.
Author Contributions
Planning and design of the study, R.P. and A.C.; conducting and sampling, R.P. and A.C.; sample analysis, R.P.; statistical analysis, R.P. and A.C.; manuscript drafting, R.P.; manuscript editing and finalizing, R.P. and A.C.; All authors have read and agreed to the published version of the manuscript.
Funding
The authors express their sincerest gratitude to the Fundamental Fund of Khon Kaen University, which has received funding support from the National Science, Research and Innovation Fund (NSRF) for providing financial support. The Research Program on the Research and Development of Winged Bean Root Utilization as Ruminant Feed, and Graduate Studies, Khon Kaen University also partially supported this work.
Institutional Review Board Statement
The study was conducted under approval record No. U1-02494-2559 by Animal Ethics Committee of Khon Kaen University with the guidelines of the National Research Council of Thailand for the ethical treatment of animals in research.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to express our sincere thanks to the Tropical Feed Resources Research and Development Center (TROFREC), Department of Animal Science, Faculty of Agriculture, Khon Kaen University for the use of the research facilities.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Olivier, J.G.J.; Peters, J.A.H.W. Trends in Global CO2 and Total Greenhouse Gas Emissions: 2019 Report; PBL Neth Environ Assess Agency: Hague, The Netherlands, 2020; p. 70. [Google Scholar]
- Eugène, M.; Sauvant, D.; Noziere, P.; Viallard, D.; Oueslati, K.; Lherm, M.; Mathias, E.; Doreau, M. A new Tier 3 method to calculate methane emission inventory for ruminants. J. Environ. Manag. 2019, 231, 982–988. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.A.; Johnson, D.E. Methane emissions from cattle. J. Anim. Sci. 1995, 73, 2483–2492. [Google Scholar] [CrossRef] [PubMed]
- Gunun, P.; Wanapat, M.; Anantasook, N.; Cherdthong, A. Effects of condensed tannins in Mao (Antidesma thwaitesianum Muell. Arg.) seed meal on rumen fermentation characteristics and nitrogen utilization in goats. Asian-Australas. J. Anim. Sci. 2016, 29, 1111–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmquist, D.; Jenkins, T. A 100-Year Review: Fat feeding of dairy cows. J. Dairy Sci. 2017, 100, 10061–10077. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Hess, P.; Williams, S.; Jacobs, J.; Hannah, M.; Beauchemin, K.; Eckard, R.; Wales, W.; Morris, G.; Moate, P. Effect of dietary fat supplementation on methane emissions from dairy cows fed wheat or corn. J. Dairy Sci. 2019, 102, 2714–2723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haque, M.N. Dietary manipulation: A sustainable way to mitigate methane emissions from ruminants. J. Anim. Sci. Technol. 2018, 60, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.; Lee, S.-S. Advanced estimation and mitigation strategies: A cumulative approach to enteric methane abatement from ruminants. J. Anim. Sci. Technol. 2019, 61, 122. [Google Scholar] [CrossRef] [Green Version]
- Beck, M.; Thompson, L.; Williams, G.; Place, S.; Gunter, S.; Reuter, R. Fat supplements differing in physical form improve performance but divergently influence methane emissions of grazing beef cattle. Anim. Feed Sci. Technol. 2019, 254, 114210. [Google Scholar] [CrossRef]
- Drehmel, O.; Brown-Brandl, T.; Judy, J.; Fernando, S.C.; Miller, P.S.; Hales, K.; Kononoff, P.J. The influence of fat and hemicellulose on methane production and energy utilization in lactating Jersey cattle. J. Dairy Sci. 2018, 101, 7892–7906. [Google Scholar] [CrossRef]
- Patra, A.K. The effect of dietary fats on methane emissions, and its other effects on digestibility, rumen fermentation and lactation performance in cattle: A meta-analysis. Livest. Sci. 2013, 155, 244–254. [Google Scholar] [CrossRef]
- Meneguz, M.; Schiavone, A.; Gai, F.; Dama, A.; Lussiana, C.; Renna, M.; Gasco, L. Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia illucens) larvae. J. Sci. Food Agric. 2018, 98, 5776–5784. [Google Scholar] [CrossRef]
- Ahmed, E.; Fukuma, N.; Hanada, M.; Nishida, T. Insects as novel ruminant feed and a potential mitigation strategy for methane emissions. Animals 2021, 11, 2648. [Google Scholar] [CrossRef] [PubMed]
- Makkar, H.P.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Belforti, M.; Gai, F.; Lussiana, C.; Renna, M.; Malfatto, V.; Rotolo, L.; De Marco, M.; Dabbou, S.; Schiavone, A.; Zoccarato, I. Tenebrio molitor meal in rainbow trout (Oncorhynchus mykiss) diets: Effects on animal performance, nutrient digestibility and chemical composition of fillets. Ital. J. Anim. Sci. 2015, 14, 4170. [Google Scholar] [CrossRef] [Green Version]
- Spranghers, T.; Ottoboni, M.; Klootwijk, C.; Ovyn, A.; Deboosere, S.; De Meulenaer, B.; Michiels, J.; Eeckhout, M.; De Clercq, P.; De Smet, S. Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. J. Sci. Food Agric. 2017, 97, 2594–2600. [Google Scholar] [CrossRef] [PubMed]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1998; Volume 2. [Google Scholar]
- Van Soest, P.V.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- So, S.; Cherdthong, A.; Wanapat, M. Growth performances, nutrient digestibility, ruminal fermentation and energy partition of Thai native steers fed exclusive rice straw and fermented sugarcane bagasse with Lactobacillus, cellulase and molasses. J. Anim. Physiol. Anim. Nutr. 2022, 106, 45–54. [Google Scholar] [CrossRef]
- Yamamoto-Osaki, T.; Kamiya, S.; Sawamura, S.; Kai, M.; Ozawa, A. Growth inhibition of Clostridium difficile by intestinal flora of infant faeces in continuous flow culture. J. Med. Microbiol. 1994, 40, 179–187. [Google Scholar] [CrossRef]
- Fawcett, J.; Scott, J. A rapid and precise method for the determination of urea. J. Clin. Pathol. 1960, 13, 156–159. [Google Scholar] [CrossRef] [Green Version]
- Galyean, M. Laboratory Procedure in Animal Nutrition Research; Department of Animal and Life Science New Mexico State University: Las Cruces, NM, USA, 1989; Volume 188. [Google Scholar]
- Tilley, J.; Terry, D.R. A two-stage technique for the in vitro digestion of forage crops. Grass Forage Sci. 1963, 18, 104–111. [Google Scholar] [CrossRef]
- Schofield, P. Gas production methods. Chapter 10. In Farm Animal Metabolism and Nutrition; D’Mello, J.P.F., Ed.; CABI Publishing: Ithaca, NY, USA, 2000; pp. 209–232. [Google Scholar]
- Statistical Analysis Systems. SAS/STAT User’s Guide: Version 6.12, 4th ed.; SAS Inc.: Cary, NC, USA, 1996. [Google Scholar]
- Flores-Santiago, E.d.J.; González-Garduño, R.; Vaquera-Huerta, H.; Calzada-Marín, J.M.; Cadena-Villegas, S.; Arceo-Castillo, J.I.; Vázquez-Mendoza, P.; Ku-Vera, J.C. Reduction of enteric methane emissions in heifers fed tropical grass-based rations supplemented with palm oil. Fermentation 2022, 8, 349. [Google Scholar] [CrossRef]
- Thao, N.T.; Wanapat, M.; Kang, S.; Cherdthong, A. effects of supplementation of eucalyptus (E. camaldulensis) leaf meal on feed intake and rumen fermentation efficiency in swamp buffaloes. Asian-Australas. J. Anim. Sci. 2015, 28, 951–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kongmun, P.; Wanapat, M.; Pakdee, P.; Navanukraw, C. Effect of coconut oil and garlic powder on in vitro fermentation using gas production technique. Livest. Sci. 2010, 127, 38–44. [Google Scholar] [CrossRef]
- Kang, S.; Wanapat, M.; Phesatcha, K.; Norrapoke, T.; Foiklang, S.; Ampapon, T.; Phesatcha, B. Using krabok (Irvingia malayana) seed oil and Flemingia macrophylla leaf meal as a rumen enhancer in an in vitro gas production system. Anim. Prod. Sci. 2016, 57, 327–333. [Google Scholar] [CrossRef]
- Prachumchai, R.; Cherdthong, A.; Wanapat, M. Screening of cyanide-utilizing bacteria from rumen and in vitro evaluation of fresh cassava root utilization with pellet containing high sulfur diet. Vet. Sci. 2021, 8, 10. [Google Scholar] [CrossRef]
- Ramos, S.C.; Jeong, C.D.; Mamuad, L.L.; Kim, S.H.; Kang, S.H.; Kim, E.T.; Cho, Y.I.; Lee, S.S.; Lee, S.S. Diet transition from high-forage to high-concentrate alters rumen bacterial community composition, epithelial transcriptomes and ruminal fermentation parameters in dairy cows. Animals 2021, 11, 838. [Google Scholar] [CrossRef]
- Prachumchai, R.; Cherdthong, A.; Wanapat, M.; So, S.; Polyorach, S. Fresh cassava root replacing cassava chip could enhance milk production of lactating dairy cows fed diets based on high sulfur-containing pellet. Sci. Rep. 2022, 12, 3809. [Google Scholar] [CrossRef] [PubMed]
- Calabrò, S.; Tudisco, R.; Balestrieri, A.; Piccolo, G.; Infascelli, F.; Cutrignelli, M.I. Fermentation characteristics of different grain legumes cultivars with the in vitro gas production technique. Ital. J. Anim. Sci. 2009, 8, 280. [Google Scholar] [CrossRef] [Green Version]
- Cherdthong, A.; Prachumchai, R.; Supapong, C.; Khonkhaeng, B.; Wanapat, M.; Foiklang, S.; Milintawisamai, N.; Gunun, N.; Gunun, P.; Chanjula, P. Inclusion of yeast waste as a protein source to replace soybean meal in concentrate mixture on ruminal fermentation and gas kinetics using in vitro gas production technique. Anim. Prod. Sci. 2018, 59, 1682–1688. [Google Scholar] [CrossRef]
- Castillejos, L.; Calsamiglia, S.; Martín-Tereso, J.; Ter Wijlen, H. In vitro evaluation of effects of ten essential oils at three doses on ruminal fermentation of high concentrate feedlot-type diets. Anim. Feed Sci. Technol. 2008, 145, 259–270. [Google Scholar] [CrossRef]
- Castillejos, L.; Calsamiglia, S.; Ferret, A.; Losa, R. Effects of dose and adaptation time of a specific blend of essential oil compounds on rumen fermentation. Anim. Feed Sci. Technol. 2007, 132, 186–201. [Google Scholar] [CrossRef]
- Ueda, K.; Ferlay, A.; Chabrot, J.; Loor, J.; Chilliard, Y.; Doreau, M. Effect of linseed oil supplementation on ruminal digestion in dairy cows fed diets with different forage: Concentrate ratios. J. Dairy Sci. 2003, 86, 3999–4007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, M.; Kobayashi, T.; Itabashi, H. Effects of the absence of rumen ciliate protozoa on urinary allantoin excretion in goats. Anim. Sci. Technol. 1991, 62, 939–946. [Google Scholar]
- Beauchemin, K.; McGinn, S.; Benchaar, C.; Holtshausen, L. Crushed sunflower, flax, or canola seeds in lactating dairy cow diets: Effects on methane production, rumen fermentation, and milk production. J. Dairy Sci. 2009, 92, 2118–2127. [Google Scholar] [CrossRef]
- Machmüller, A. Medium-chain fatty acids and their potential to reduce methanogenesis in domestic ruminants. Agric. Ecosyst. Environ. 2006, 112, 107–114. [Google Scholar] [CrossRef]
- Phesatcha, K.; Phesatcha, B.; Wanapat, M.; Cherdthong, A. The effect of yeast and roughage concentrate ratio on ruminal pH and protozoal population in Thai native beef cattle. Animals 2021, 12, 53. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, C.; Huasai, S.; Chen, A. Effects of dietary forage to concentrate ratio on nutrient digestibility, ruminal fermentation and rumen bacterial composition in Angus cows. Sci. Rep. 2021, 11, 17023. [Google Scholar] [CrossRef] [PubMed]
- Musco, N.; Tudisco, R.; Grossi, M.; Mastellone, V.; Morittu, V.M.; Pero, M.E.; Wanapat, M.; Trinchese, G.; Cavaliere, G.; Mollica, M.P.; et al. Effect of a high forage : concentrate ratio on milk yield, blood parameters and oxidative status in lactating cows. Anim. Prod. Sci. 2020, 60, 1531–1538. [Google Scholar] [CrossRef]
- Arcos-Álvarez, D.N.; Aguilar-Urquizo, E.; Sanginés-García, J.R.; Chay-Canul, A.J.; Molina-Botero, I.; Tzec-Gamboa, M.; Vargas-Bello-Pérez, E.; Piñeiro-Vázquez, Á.T. Effect of adding extra virgin olive oil to hair sheep lambs’ diets on productive performance, ruminal fermentation kinetics and rumen ciliate protozoa. Animals 2022, 12, 2588. [Google Scholar] [CrossRef]
- van Cleef, F.D.O.S.; Ezequiel, J.M.B.; D’Aurea, A.P.; Almeida, M.T.C.; Perez, H.L.; van Cleef, E.H.C.B. Feeding behavior, nutrient digestibility, feedlot performance, carcass traits, and meat characteristics of crossbred lambs fed high levels of yellow grease or soybean oil. Small Rumin. Res. 2016, 137, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Sutton, J.; Knight, R.; McAllan, A.; Smith, R. Digestion and synthesis in the rumen of sheep given diets supplemented with free and protected oils. Br. J. Nutr. 1983, 49, 419–432. [Google Scholar] [CrossRef]
- Supapong, C.; Cherdthong, A.; Seankamsorn, A.; Khonkhaeng, B.; Wanapat, M.; Uriyapongson, S.; Gunun, N.; Gunun, P.; Chanjula, P.; Polyorach, S. In vitro fermentation, digestibility and methane production as influenced by Delonix regia seed meal containing tannins and saponins. J. Anim. Feed Sci. 2017, 26, 123–130. [Google Scholar] [CrossRef]
- Cherdthong, A.; Prachumchai, R.; Wanapat, M. In vitro evaluations of pellets containing Delonix regia seed meal for ruminants. Trop. Anim. Health Prod. 2019, 51, 2003–2010. [Google Scholar] [CrossRef] [PubMed]
- Costa, D.; Quigley, S.; Isherwood, P.; McLennan, S.; Sun, X.; Gibbs, S.; Poppi, D. The inclusion of low quantities of lipids in the diet of ruminants fed low quality forages has little effect on rumen function. Anim. Feed Sci. Technol. 2017, 234, 20–28. [Google Scholar] [CrossRef]
- Mapato, C.; Wanapat, M.; Cherdthong, A. Effects of urea treatment of straw and dietary level of vegetable oil on lactating dairy cows. Trop. Anim. Health Prod. 2010, 42, 1635–1642. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effect of roughage-to-concentrate (R:C) ratio level combined with black soldier fly oil (BSFO) level on cumulative gas during incubation times.
Figure 1.
Effect of roughage-to-concentrate (R:C) ratio level combined with black soldier fly oil (BSFO) level on cumulative gas during incubation times.


{kind=link}
Table 1.
Ingredient and chemical composition of concentrate diet and rice straw (%DM).
| Item | BSFO 0% | BSFO 2% | BSFO 4% | BSFO 6% | Rice Straw |
|---|---|---|---|---|---|
| Ingredients, % DM | |||||
| Cassava chips | 41.0 | 40.0 | 40.0 | 39.0 | |
| Corn meal | 15.0 | 14.0 | 14.0 | 14.0 | |
| Rice bran | 10.0 | 10.0 | 9.0 | 9.0 | |
| Soybean meal (SBM) | 7.0 | 7.0 | 7.0 | 7.0 | |
| Dried brewers’ grains | 11.0 | 11.0 | 10.5 | 10.0 | |
| Palm kernel meal | 11.0 | 11.0 | 10.4 | 9.8 | |
| Black soldier fly oil (BSFO) 1 | 0.0 | 2.0 | 4.0 | 6.0 | |
| Vitamin and mineral premix 2 | 1.0 | 1.0 | 1.0 | 1.0 | |
| Urea | 1.0 | 1.0 | 1.1 | 1.2 | |
| Sodium chloride | 1.0 | 1.0 | 1.0 | 1.0 | |
| Molasses | 2.0 | 2.0 | 2.0 | 2.0 | |
| Chemical composition | |||||
| Dry matter, % | 88.39 | 88.34 | 88.59 | 88.11 | 88.57 |
| Organic matter, % DM | 94.57 | 94.77 | 95.05 | 95.14 | 89.56 |
| Crude protein, % DM | 15.21 | 15.45 | 15.05 | 15.05 | 2.54 |
| Ether extract, % DM | 2.05 | 4.02 | 6.03 | 7.96 | 0.56 |
| Neutral detergent fiber, % DM | 21.23 | 22.48 | 21.61 | 19.90 | 72.89 |
| Acid detergent fiber, % DM | 13.36 | 14.33 | 13.33 | 12.37 | 50.14 |
| Gross energy (MJ) | 16.98 | 17.44 | 17.51 | 17.67 | 14.67 |
1 Black soldier fly larvae oil contains the following fatty acid composition: 40.1% Lauric acid (C12: 0), 16.3% Oleic acid (C18:1 n-9), 15.0% Linoleic acid (C18:2 n-6), 13.2% Myristic acid (C14:0), and 13.7% Palmitic acid (C16:0) of the total fatty acids, respectively. 2 Contains per kilogram premix: 10,000,000 IU vitamin A; 70,000 IU vitamin E; 1,600,000 IU vitamin D; 50 g iron; 40 g zinc; 40 g manganese; 0.1 g cobalt; 10 g copper; 0.1 g selenium; 0.5 g iodine.
Table 2.
Effect of roughage-to-concentrate (R:C) ratio level combined with black soldier fly oil (BSFO) level on gas kinetics and cumulative gas at 96 h after incubation.
Table 2.
Effect of roughage-to-concentrate (R:C) ratio level combined with black soldier fly oil (BSFO) level on gas kinetics and cumulative gas at 96 h after incubation.
| Treatment | R:C Ratio | BSFO (%) | Gas Kinetics (mL/0.5 g) | Cumulative Gas (mL/g DM Basis) | ||
|---|---|---|---|---|---|---|
| b | c | L | ||||
| T1 | 60:40 | 0 | 134.47 a | 0.03 | 0.333 | 126.57 b |
| T2 | 60:40 | 2 | 132.83 ab | 0.03 | 0.467 | 124.60 bc |
| T3 | 60:40 | 4 | 134.60 a | 0.03 | 0.367 | 126.12 b |
| T4 | 60:40 | 6 | 129.17 bc | 0.03 | 0.367 | 120.00 c |
| T5 | 40:60 | 0 | 135.33 a | 0.13 | 0.533 | 132.11 a |
| T6 | 40:60 | 2 | 128.37 c | 0.04 | 0.533 | 124.56 bc |
| T7 | 40:60 | 4 | 123.13 d | 0.04 | 0.433 | 119.83 c |
| T8 | 40:60 | 6 | 117.75 e | 0.04 | 0.650 | 114.26 d |
| SEM | 1.359 | 0.032 | 0.072 | 1.510 | ||
| Main effects | ||||||
| R:C Ratio | 60:40 | 132.77 a | 0.03 | 0.54 b | 124.32 | |
| 40:60 | 126.15 b | 0.06 | 0.38 a | 122.69 | ||
| BSFO (%) | 0 | 134.90 a | 0.08 | 0.51 | 129.34 a | |
| 2 | 130.60 b | 0.03 | 0.50 | 124.58 b | ||
| 4 | 128.87 b | 0.03 | 0.43 | 122.97 b | ||
| 6 | 123.46 c | 0.03 | 0.40 | 117.13 c | ||
| Significance of main effect and interaction | ||||||
| R:C Ratio | <0.01 | 0.1826 | <0.01 | 0.1457 | ||
| BSFO (%) | <0.01 | 0.3875 | 0.3982 | <0.01 | ||
| Interaction | <0.01 | 0.4100 | 0.3842 | <0.01 | ||
a–e Means in the same column with different lowercase letters differ (p < 0.05); b = the final asymptotic gas volume corresponding to fully digested substrate (mL/g DM); c = a rate constant (units time 1); L = a discontinuous lag term (h).
Table 3.
Effect of roughage-to-concentrate (R:C) ratio level combined with black soldier fly oil (BSFO) level on ruminal pH, ammonia-nitrogen (NH3-N) concentration, protozoal population, and methane production.
Table 3.
Effect of roughage-to-concentrate (R:C) ratio level combined with black soldier fly oil (BSFO) level on ruminal pH, ammonia-nitrogen (NH3-N) concentration, protozoal population, and methane production.
| Treatment | R:C Ratio | BSFO (%) | pH | NH3-N (mg/dL) | Methane (mL/1 g Dry Matter Substrate) | Protozoa (×106 Cell/mL) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| 4 h | 8 h | 4 h | 8 h | 4 h | 8 h | 4 h | 8 h | |||
| T1 | 60:40 | 0 | 7.03 a | 6.94 | 19.84 b | 20.40 | 4.50 a | 9.45 b | 8.00 | 8.00 |
| T2 | 60:40 | 2 | 6.97 bc | 6.94 | 19.14 b | 20.25 | 4.60 a | 8.65 bc | 4.00 | 4.00 |
| T3 | 60:40 | 4 | 6.97 bc | 6.92 | 18.42 b | 17.53 | 2.45 c | 9.00 c | 3.00 | 6.00 |
| T4 | 60:40 | 6 | 6.98 bc | 6.96 | 18.96 b | 17.01 | 3.30 b | 7.30 bcd | 3.00 | 4.00 |
| T5 | 40:60 | 0 | 6.95 c | 6.89 | 22.41 a | 20.67 | 4.55 a | 11.70 a | 9.00 | 10.00 |
| T6 | 40:60 | 2 | 6.97 bc | 6.87 | 21.95 a | 18.18 | 4.55 a | 7.30 bcd | 5.00 | 6.00 |
| T7 | 40:60 | 4 | 7.00 abc | 6.88 | 21.52 a | 17.22 | 3.60 b | 6.30 d | 2.00 | 4.00 |
| T8 | 40:60 | 6 | 7.02 abc | 6.87 | 18.96 b | 17.01 | 2.65 c | 6.40 d | 3.00 | 2.00 |
| SEM | 0.016 | 0.015 | 0.389 | 0.582 | 0.279 | 0.659 | 1.061 | 1.173 | ||
| Main effects | ||||||||||
| R:C Ratio | 60:40 | 6.99 | 6.94 a | 19.19 b | 18.80 | 3.71 | 8.60 | 4.50 | 5.50 | |
| 40:60 | 6.98 | 6.88 b | 21.19 a | 18.20 | 3.84 | 7.93 | 4.75 | 5.50 | ||
| BSFO (%) | 0 | 6.99 | 6.92 | 21.13 a | 20.51 a | 4.53 a | 10.58 a | 8.50 a | 9.00 a | |
| 2 | 6.97 | 6.91 | 20.54 a | 19.73 a | 4.58 a | 7.98 b | 4.50 b | 5.00 b | ||
| 4 | 6.98 | 6.90 | 20.28 a | 17.38 b | 3.03 b | 7.65 b | 2.50 b | 5.00 b | ||
| 6 | 7.00 | 6.92 | 18.91 b | 17.28 b | 2.98 b | 6.85 b | 3.00 b | 3.00 b | ||
| Significance of main effect and interaction | ||||||||||
| R:C Ratio | 0.6650 | <0.01 | <0.01 | 0.2025 | 0.3555 | 0.1855 | 0.7475 | 1.00 | ||
| BSFO (%) | 0.2900 | 0.7099 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | ||
| Interaction | <0.01 | 0.4157 | <0.01 | 0.591 | <0.01 | <0.05 | 0.7520 | 0.2018 | ||
a–d Means in the same column with different lowercase letters differ (p < 0.05).
Table 4.
Effect of roughage-to-concentrate (R:C) ratio level combined with black soldier fly oil (BSFO) level on the in vitro degradability of nutrients.
Table 4.
Effect of roughage-to-concentrate (R:C) ratio level combined with black soldier fly oil (BSFO) level on the in vitro degradability of nutrients.
| Treatment | R:C Ratio | BSFO (%) | IVDMD (% DM) | IVOMD (% DM) | IVTD (% DM) | |||
|---|---|---|---|---|---|---|---|---|
| 12 h | 24 h | 12 h | 24 h | 12 h | 24 h | |||
| T1 | 60:40 | 0 | 31.08 | 37.34 | 39.90 bc | 45.69 | 73.39 | 79.59 b |
| T2 | 60:40 | 2 | 29.63 | 36.39 | 38.29 dc | 44.40 | 73.45 | 79.48 b |
| T3 | 60:40 | 4 | 28.59 | 37.09 | 37.26 d | 45.19 | 73.55 | 78.70 b |
| T4 | 60:40 | 6 | 27.90 | 35.61 | 36.42 d | 43.52 | 74.14 | 76.51 c |
| T5 | 40:60 | 0 | 35.00 | 45.74 | 41.93 ab | 52.27 | 83.72 | 84.75 a |
| T6 | 40:60 | 2 | 34.10 | 45.24 | 40.85 ab | 51.79 | 83.81 | 84.16 a |
| T7 | 40:60 | 4 | 35.14 | 45.49 | 42.12 ab | 51.80 | 84.12 | 84.26 a |
| T8 | 40:60 | 6 | 35.74 | 43.46 | 43.00 a | 49.89 | 83.98 | 85.45 a |
| SEM | 0.684 | 0.264 | 0.704 | 0.286 | 0.606 | 0.482 | ||
| Main effects | ||||||||
| R:C Ratio | 60:40 | 29.30 b | 36.61 b | 37.97 b | 44.70 b | 73.63 b | 78.57 b | |
| 40:60 | 35.12 a | 44.98 a | 41.97 a | 51.44 a | 83.93 a | 84.66 a | ||
| BSFO (%) | 0 | 33.04 | 41.54 a | 40.92 | 48.98 a | 77.53 | 82.17 | |
| 2 | 31.12 | 40.82 b | 39.57 | 48.09 b | 78.63 | 81.82 | ||
| 4 | 31.87 | 41.29 ab | 39.69 | 48.49 ab | 78.83 | 81.48 | ||
| 6 | 31.82 | 39.53 c | 39.71 | 46.70 c | 79.06 | 80.98 | ||
| Significance of main effect and interaction | ||||||||
| R:C Ratio | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | ||
| BSFO (%) | 0.2621 | <0.01 | 0.2605 | <0.01 | 0.1279 | 0.1256 | ||
| Interaction | 0.0604 | 0.3689 | <0.05 | 0.3674 | 1.00 | <0.01 | ||
a–d Means in the same column with different lowercase letters differ (p < 0.05); IVDMD = in vitro dry matter digestibility; IVOMD = in vitro organic matter digestibility; IVTD = in vitro true digestibility.
Table 5.
Effect of roughage-to-concentrate (R:C) ratio level combined with black soldier fly oil (BSFO) level on volatile fatty acid (VFA) concentrations.
Table 5.
Effect of roughage-to-concentrate (R:C) ratio level combined with black soldier fly oil (BSFO) level on volatile fatty acid (VFA) concentrations.
| Treatment | R:C Ratio | BSFO (%) | Total VFAs (mmol/L) | Acetate (%) | Propionate (%) | Butyrate (%) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| 4 h | 8 h | 4 h | 8 h | 4 h | 8 h | 4 h | 8 h | |||
| T1 | 60:40 | 0 | 42.55 | 52.85 | 72.63 | 72.53 | 14.92 | 15.54 d | 12.45 | 11.93 ab |
| T2 | 60:40 | 2 | 41.03 | 53.54 | 72.54 | 70.35 | 14.78 | 16.95 c | 12.68 | 12.70 a |
| T3 | 60:40 | 4 | 42.77 | 51.69 | 72.76 | 69.30 | 14.86 | 17.95 bc | 12.68 | 12.76 a |
| T4 | 60:40 | 6 | 41.80 | 53.97 | 72.31 | 70.07 | 15.12 | 18.38 b | 12.59 | 11.55 b |
| T5 | 40:60 | 0 | 40.17 | 58.76 | 71.36 | 70.57 | 15.15 | 16.97 c | 13.22 | 12.46 ab |
| T6 | 40:60 | 2 | 41.76 | 61.53 | 72.54 | 70.39 | 15.06 | 18.14 b | 12.67 | 11.47 b |
| T7 | 40:60 | 4 | 41.99 | 62.49 | 72.04 | 67.36 | 15.19 | 21.10 a | 12.88 | 11.54 b |
| T8 | 40:60 | 6 | 40.30 | 61.55 | 72.26 | 67.63 | 15.03 | 20.67 a | 12.83 | 11.71 ab |
| SEM | 1.235 | 2.105 | 0.455 | 0.547 | 0.303 | 0.350 | 0.234 | 0.336 | ||
| Main effects | ||||||||||
| R:C ratio | 60:40 | 41.99 | 53.01 b | 72.56 | 70.56 a | 14.92 | 17.20 b | 12.61 | 12.23 | |
| 40:60 | 41.13 | 61.08 a | 72.05 | 68.99 b | 15.10 | 19.22 a | 12.90 | 11.79 | ||
| BSFO (%) | 0 | 41.36 | 55.80 | 71.87 | 71.55 a | 15.03 | 16.26 c | 12.91 | 12.19 | |
| 2 | 41.39 | 57.54 | 72.54 | 70.37 b | 14.92 | 17.55 b | 12.68 | 12.09 | ||
| 4 | 42.46 | 57.09 | 72.40 | 68.33 c | 14.99 | 19.52 a | 12.80 | 12.15 | ||
| 6 | 41.20 | 57.76 | 72.29 | 68.85 c | 15.09 | 19.52 a | 12.74 | 11.63 | ||
| Significance of main effect and interaction | ||||||||||
| R:C ratio | 0.3886 | <0.01 | 0.1459 | <0.01 | 0.4556 | <0.01 | 0.1308 | 0.0823 | ||
| BSFO (%) | 0.7822 | 0.7920 | 0.5599 | <0.01 | 0.9584 | <0.01 | 0.8077 | 0.3391 | ||
| Interaction | 0.7184 | 0.7098 | 0.5835 | 0.1481 | 0.9066 | <0.05 | 0.5100 | <0.05 | ||
a–d Means in the same column with different lowercase letters differ (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Prachumchai, R.; Cherdthong, A. Black Soldier Fly Larva Oil in Diets with Roughage to Concentrate Ratios on Fermentation Characteristics, Degradability, and Methane Generation. Animals 2023, 13, 2416. https://doi.org/10.3390/ani13152416
AMA Style
Prachumchai R, Cherdthong A. Black Soldier Fly Larva Oil in Diets with Roughage to Concentrate Ratios on Fermentation Characteristics, Degradability, and Methane Generation. Animals. 2023; 13(15):2416. https://doi.org/10.3390/ani13152416
Chicago/Turabian StylePrachumchai, Rittikeard, and Anusorn Cherdthong. 2023. "Black Soldier Fly Larva Oil in Diets with Roughage to Concentrate Ratios on Fermentation Characteristics, Degradability, and Methane Generation" Animals 13, no. 15: 2416. https://doi.org/10.3390/ani13152416
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.