


The Exploitation of Toxic Fish from the Terminal Pleistocene in Maritime Southeast Asia: A Case Study from the Mindoro Archaeological Sites, Philippines

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
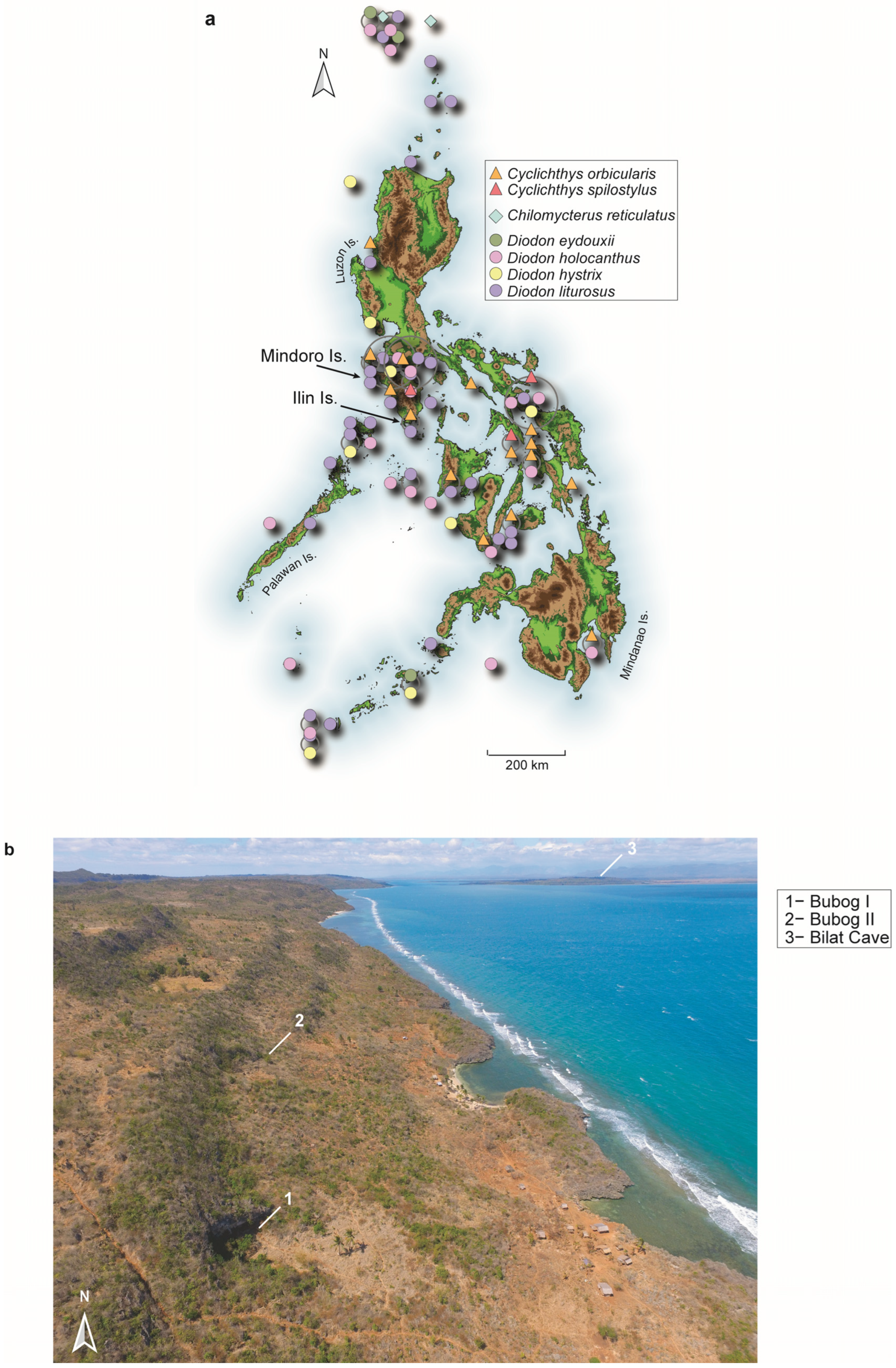
2.1. Study Sites
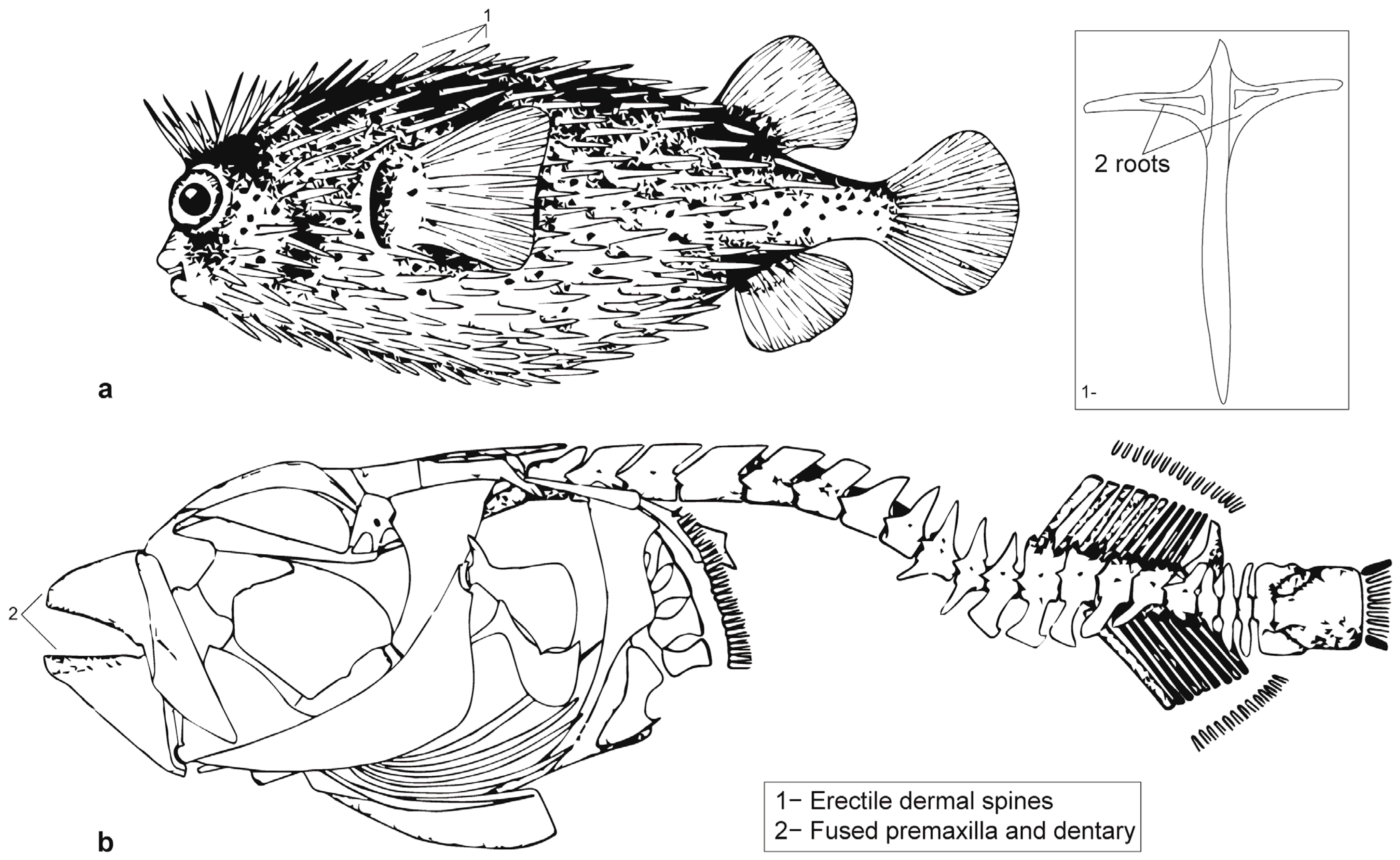
2.2. Taxonomy
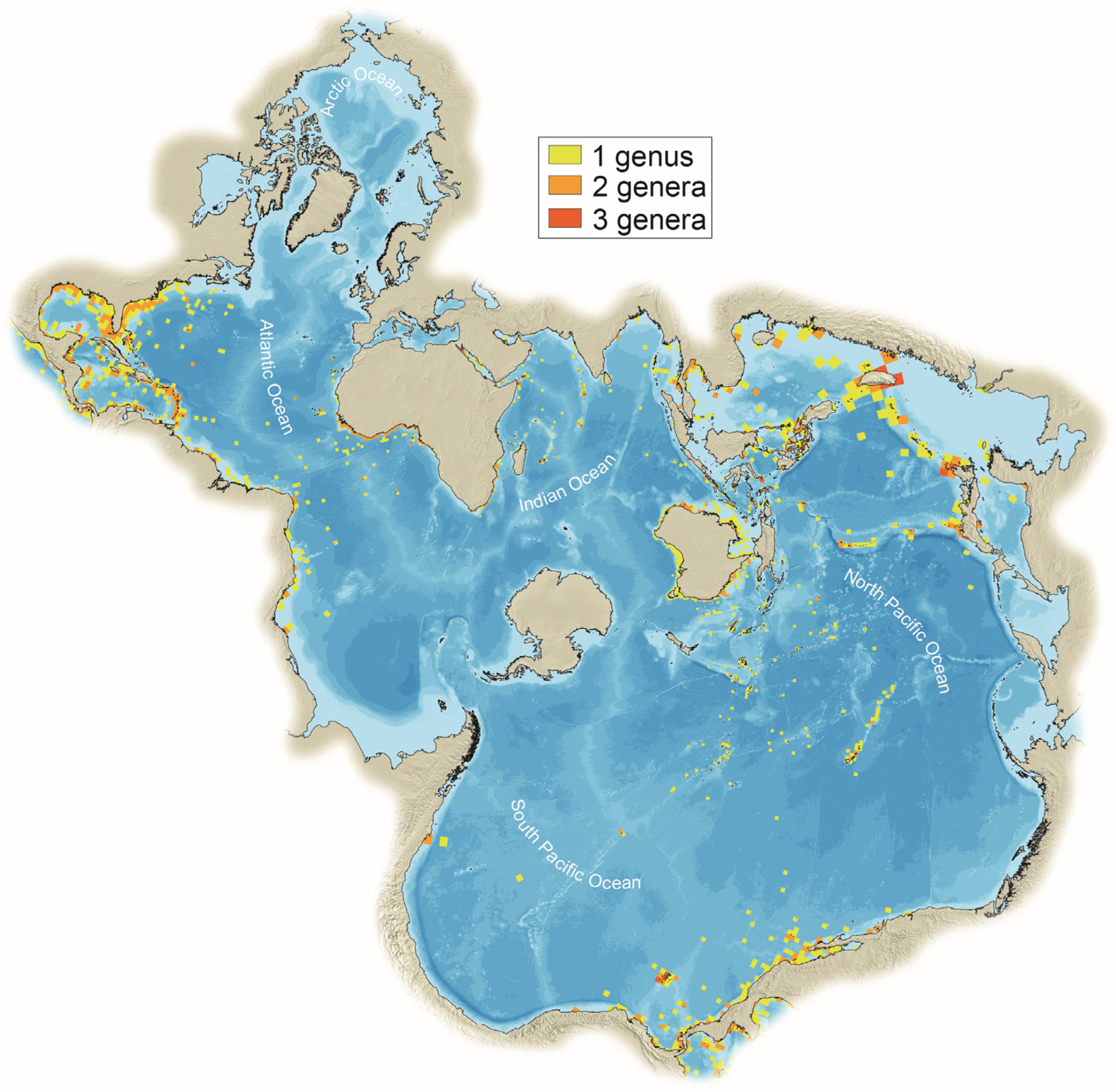
2.3. Ecology of Tetraodontoidei Fishes
2.4. Zooarchaeology
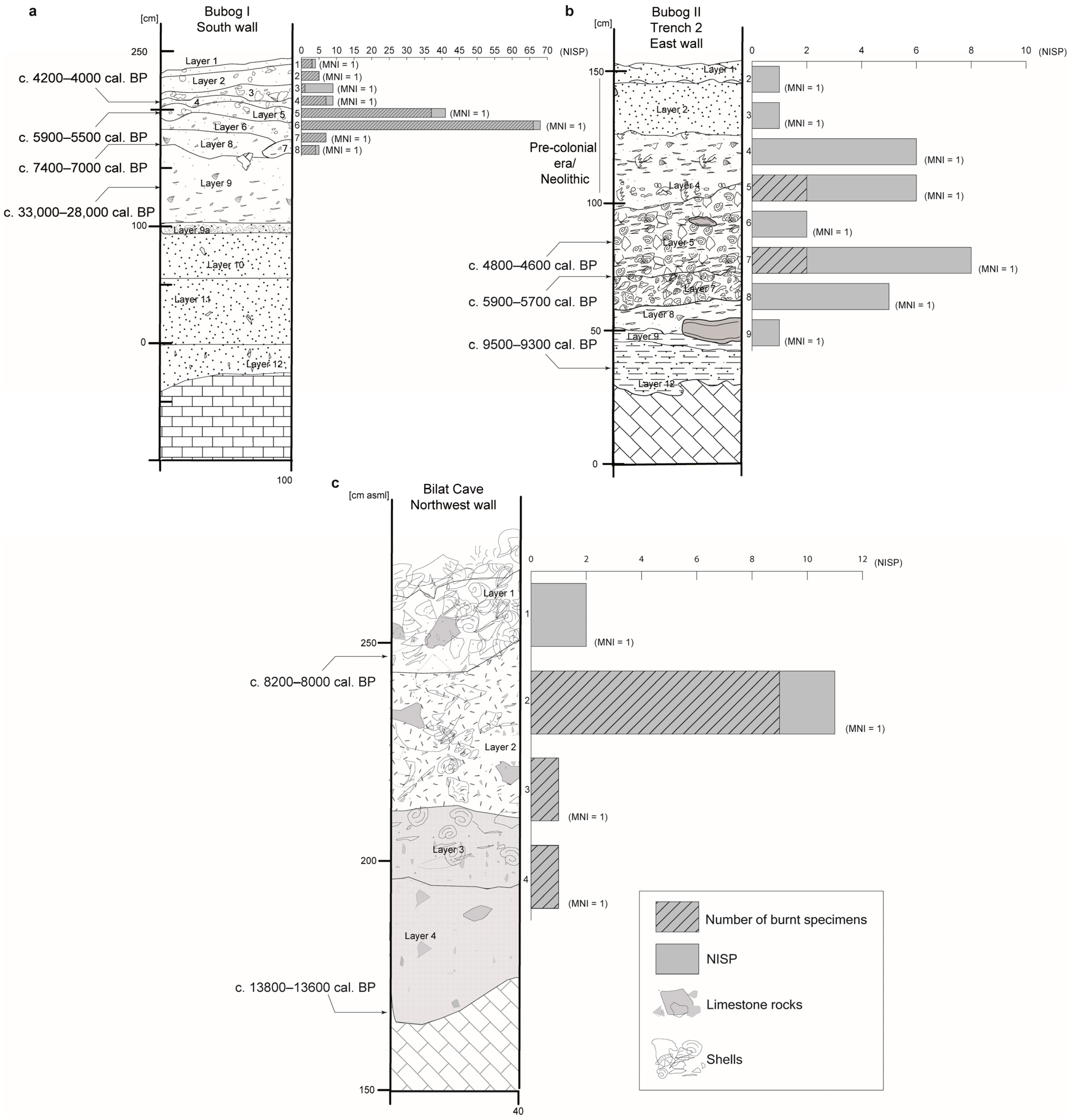
3. Results
3.1. Bubog I
3.2. Bubog II
3.3. Bilat Cave
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borgia, V. Chapter 1. The Prehistory of Poison Arrows. In Toxicology in Antiquity, 2nd ed.; Wexler, P., Ed.; History of Toxicology and Environmental Health; Academic Press: Cambridge, MA, USA, 2019; pp. 1–10. ISBN 978-0-12-815339-0. [Google Scholar]
- Perrot, E.; Vogt, E. Poisons de Flèches et Poisons d’Epreuve; Vigot Frères: Paris, France, 1913. [Google Scholar]
- Bae, C.J.; Douka, K.; Petraglia, M.D. On the Origin of Modern Humans: Asian Perspectives. Science 2017, 358, eaai9067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borgia, V.; Carlin, M.G.; Crezzini, J. Poison, Plants and Palaeolithic Hunters. An Analytical Method to Investigate the Presence of Plant Poison on Archaeological Artefacts. Quat. Int. 2017, 427, 94–103. [Google Scholar] [CrossRef]
- D’Errico, F.; Backwell, L.; Villa, P.; Degano, I.; Lucejko, J.J.; Bamford, M.K.; Higham, T.F.G.; Colombini, M.P.; Beaumont, P.B. Early Evidence of San Material Culture Represented by Organic Artifacts from Border Cave, South Africa. Proc. Natl. Acad. Sci. USA 2012, 109, 13214–13219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, A.A. Arrow Poisons in the Palaeolithic? Proc. Natl. Acad. Sci. USA 2012, 109, E3290. [Google Scholar] [CrossRef] [Green Version]
- Erlandson, J.M.; Fitzpatrick, S.M. Oceans, Islands, and Coasts: Current Perspectives on the Role of the Sea in Human Prehistory. J. Isl. Coast. Archaeol. 2006, 1, 5–32. [Google Scholar] [CrossRef]
- Clarkson, C.; Jacobs, Z.; Marwick, B.; Fullagar, R.; Wallis, L.; Smith, M.; Roberts, R.G.; Hayes, E.; Lowe, K.; Carah, X.; et al. Human Occupation of Northern Australia by 65,000 Years Ago. Nature 2017, 547, 306. [Google Scholar] [CrossRef] [Green Version]
- Kealy, S.; Louys, J.; O’Connor, S. Least-Cost Pathway Models Indicate Northern Human Dispersal from Sunda to Sahul. J. Hum. Evol. 2018, 125, 59–70. [Google Scholar] [CrossRef]
- Walter, R.C.; Buffler, R.T.; Bruggemann, J.H.; Guillaume, M.M.M.; Berhe, S.M.; Negassi, B.; Libsekal, Y.; Cheng, H.; Edwards, R.L.; von Cosel, R.; et al. Early Human Occupation of the Red Sea Coast of Eritrea during the Last Interglacial. Nature 2000, 405, 65–69. [Google Scholar] [CrossRef]
- Henshilwood, C.S.; Marean, C.W. The Origin of Modern Human Behavior: Critique of the Models and Their Test Implications. Curr. Anthropol. 2003, 44, 627–651. [Google Scholar] [CrossRef]
- Marean, C.W.; Bar-Matthews, M.; Bernatchez, J.; Fisher, E.; Goldberg, P.; Herries, A.I.R.; Jacobs, Z.; Jerardino, A.; Karkanas, P.; Minichillo, T.; et al. Early Human Use of Marine Resources and Pigment in South Africa during the Middle Pleistocene. Nature 2007, 449, 905–908. [Google Scholar] [CrossRef]
- D’Errico, F. The Invisible Frontier. A Multiple Species Model for the Origin of Behavioral Modernity. Evol. Anthropol. Issues News Rev. 2003, 12, 188–202. [Google Scholar] [CrossRef]
- Habgood, P.J.; Franklin, N.R. The Revolution That Didn’t Arrive: A Review of Pleistocene Sahul. J. Hum. Evol. 2008, 55, 187–222. [Google Scholar] [CrossRef] [PubMed]
- Roberts, P.; Stewart, B.A. Defining the ‘Generalist Specialist’ Niche for Pleistocene Homo Sapiens. Nat. Hum. Behav. 2018, 2, 542. [Google Scholar] [CrossRef] [PubMed]
- Rabett, R.J.; Piper, P.J. The Emergence of Bone Technologies at the End of the Pleistocene in Southeast Asia: Regional and Evolutionary Implications. Camb. Archaeol. J. 2012, 22, 37–56. [Google Scholar] [CrossRef]
- Wedage, O.; Amano, N.; Langley, M.C.; Douka, K.; Blinkhorn, J.; Crowther, A.; Deraniyagala, S.; Kourampas, N.; Simpson, I.; Perera, N.; et al. Specialized Rainforest Hunting by Homo Sapiens ~45,000 Years Ago. Nat. Commun. 2019, 10, 739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingicco, T.; Amano, N.; Setiagama, K.; Moigne, A.M.; Budiman; Sémah, A.M.; Simanjuntak, T.; Sémah, F. From Food to Grave Good: Nonhuman Primate Exploitation in Early to Mid-Holocene Eastern Java (Indonesia). Curr. Anthropol. 2020, 61, 264–277. [Google Scholar] [CrossRef]
- D’Errico, F.; Backwell, L.; Villa, P.; Degano, I.; Lucejko, J.J.; Bamford, M.K.; Higham, T.F.G.; Colombini, M.P.; Beaumont, P.B. Reply to Evans: Use of Poison Remains the Most Parsimonious Explanation for Border Cave Castor Bean Extract. Proc. Natl. Acad. Sci. USA 2012, 109, E3291–E3292. [Google Scholar] [CrossRef] [Green Version]
- Pawlik, A.F. Technology, Adaptation, and Mobility in Maritime Environments in the Philippines from the Late Pleistocene to Early/Mid-Holocene. Quat. Int. 2021, 596, 109–123. [Google Scholar] [CrossRef]
- Bronk Ramsey, C. Bayesian Analysis of Radiocarbon Dates. Radiocarbon 2009, 51, 337–360. [Google Scholar] [CrossRef] [Green Version]
- Heaton, T.J.; Köhler, P.; Butzin, M.; Bard, E.; Reimer, R.W.; Austin, W.E.N.; Ramsey, C.B.; Grootes, P.M.; Hughen, K.A.; Kromer, B.; et al. Marine20—The Marine Radiocarbon Age Calibration Curve (0–55,000 Cal BP). Radiocarbon 2020, 62, 779–820. [Google Scholar] [CrossRef]
- Reimer, P.J.; Austin, W.E.N.; Bard, E.; Bayliss, A.; Blackwell, P.G.; Ramsey, C.B.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 Cal KBP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- Pawlik, A.F.; Piper, P.J.; Faylona, M.G.P.G.; Padilla, S.G.; Carlos, J.; Mijares, A.S.B.; Vallejo, B.; Reyes, M.; Amano, N.; Ingicco, T.; et al. Adaptation and Foraging from the Terminal Pleistocene to the Early Holocene: Excavation at Bubog on Ilin Island, Philippines. J. Field Archaeol. 2014, 39, 230–247. [Google Scholar] [CrossRef]
- Boulanger, C.; Ingicco, T.; Piper, P.J.; Amano, N.; Grouard, S.; Ono, R.; Hawkins, S.; Pawlik, A.F. Coastal Subsistence Strategies and Mangrove Swamp Evolution at Bubog I Rockshelter (Ilin Island, Mindoro, Philippines) from the Late Pleistocene to the Mid-Holocene. J. Isl. Coast. Archaeol. 2019, 14, 584–604. [Google Scholar] [CrossRef]
- Pawlik, A.F.; Piper, P.J. The Philippines from c. 14,000 to 4,000 Cal. BP in Regional Context. Camb. Archaeol. J. 2019, 29, 1–22. [Google Scholar] [CrossRef]
- Boulanger, C. Etude des Comportements de Subsistance d’un Site Australo-Mélanésien: Bubog I (île d’Ilin, Mindoro, Philippines), ca. 11000–4000 ans BP Ichtyofaune, Crustacés Décapodes et Grands Mammifères. Master’s Thesis, Muséum National d’Histoire Naturelle, Paris, France, 2015. [Google Scholar]
- Boulanger, C. Aquatic Resources Exploitation and Adaptation of Anatomically Modern Human in Island Southeast Asia: Palaeoenvironmental and Cultural Implications. Ph.D. Thesis, Muséum National d’Histoire Naturelle, Paris, France, Australian National University, Canberra, Australia, 2021. [Google Scholar]
- Reyes, M.C.; Ingicco, T.; Piper, P.J.; Amano, N.; Pawlik, A.F. First Fossil Evidence of the Extinct Philippine Cloud Rat Crateromys Paulus (Muridae: Murinae: Phloeomyini) from Ilin Island, Mindoro, and Insights into Its Holocene Abundance. Proc. Biol. Soc. Wash. 2017, 130, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.K.A. Coastal Adaptation of Prehistoric Mindoro: The Archaeomalacology of Bubog 1, Ilin Island from the Terminal Pleistocene to the Mid Holocene. Master’s Thesis, University of the Philippines Diliman, Quezon City, Philippines, 2016. [Google Scholar]
- Pawlik, A.F.; Piper, P.J.; Wood, R.E.; Lim, K.K.A.; Faylona, M.G.P.G.; Mijares, A.S.B.; Porr, M. Shell Tool Technology in Island Southeast Asia: An Early Middle Holocene Tridacna Adze from Ilin Island, Mindoro, Philippines. Antiquity 2015, 89, 292–308. [Google Scholar] [CrossRef]
- Neri, L.A.M.; Pawlik, A.F.; Reepmeyer, C.; Mijares, A.S.B.; Paz, V.J. Mobility of Early Islanders in the Philippines during the Terminal Pleistocene/Early Holocene Boundary: PXRF-Analysis of Obsidian Artefacts. J. Archaeol. Sci. 2015, 61, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Voris, H.K. Maps of Pleistocene Sea Levels in Southeast Asia: Shorelines, River Systems and Time Durations. J. Biogeogr. 2000, 27, 1153–1167. [Google Scholar] [CrossRef] [Green Version]
- Sathiamurthy, E.; Voris, H.K. Maps of Holocene Sea Level Transgression and Submerged Lakes on the Sunda Shelf. Trop. Nat. Hist. 2006, 2, 1–44. [Google Scholar]
- Pawlik, A.F. Field Report on the Archaeological Excavations on Ilin Island and Sta Teresa, Mindoro Occidental, Philippines—Update on the Campaign from 21 May–4 June 2017; University of the Philippines Diliman: Quezon City, Philippines, 2017; p. 29. [Google Scholar]
- Shono, T.; Thiery, A.P.; Cooper, R.L.; Kurokawa, D.; Britz, R.; Okabe, M.; Fraser, G.J. Evolution and Developmental Diversity of Skin Spines in Pufferfishes. iScience 2019, 19, 1248–1259. [Google Scholar] [CrossRef] [Green Version]
- Brainerd, E.L. Pufferfish Inflation: Functional Morphology of Postcranial Structures in Diodon holocanthus (Tetraodontiformes). J. Morphol. 1994, 220, 243–261. [Google Scholar] [CrossRef] [PubMed]
- Panão, I.; Carrascosa, C.; Jaber, J.R.; Raposo, A. Puffer Fish and Its Consumption: To Eat or Not to Eat? Food Rev. Int. 2016, 32, 305–322. [Google Scholar] [CrossRef] [Green Version]
- Froese, R.; Pauly, D. FishBase. Available online: www.fishbase.org (accessed on 25 November 2022).
- Ahasan, H.A.M.N.; Mamun, A.A.; Karim, S.R.; Bakar, M.A.; Gazi, E.A.; Bala, C.S. Paralytic Complications of Puffer Fish (Tetrodotoxin) Poisoning. Singap. Med. J. 2004, 45, 73–74. [Google Scholar]
- Nelson, J.S. Fishes of the World; Wiley: New York, NY, USA, 2006; ISBN 978-0-471-54713-6. [Google Scholar]
- Dye, T.; Longenecker, K.R. Manual of Hawaiian Fish Remains Identification Based on the Skeletal Reference Collection of Alan C. Ziegler and Including Otoliths; Society for Hawaiian Archaeology: Honolulu, HI, USA, 2004. [Google Scholar]
- Matsuura, K. Pufferfishes and Their Allies of Japan; Tokai University Press: Tokyo, Japan, 2017. [Google Scholar]
- Kirch, P.V.; Dye, T.S. Ethno-Archaeology and the Development of Polynesian Fishing Strategies. J. Polyn. Soc. 1979, 88, 53–76. [Google Scholar]
- Lambrides, A.; Weisler, M.; Charleux, M. New Taxonomic Records and Regional Trends for the Marquesan Prehistoric Marine Fishery, Eiao Island, Polynesia. J. Pac. Archaeol. 2018, 9, 44–62. [Google Scholar]
- Carpenter, K.E.; Niem, V.H. (Eds.) The Living Marine Resources of the Western Central Pacific; FAO: Rome, Italy, 1998; ISBN 978-92-5-104051-5. [Google Scholar]
- Butler, V.L. Fish Feeding Behaviour and Fish Capture: The Case for Variation in Lapita Fishing Strategies. Archaeol. Ocean. 1994, 29, 81–90. [Google Scholar] [CrossRef]
- Densmore, N. An Archaeological Assessment of Fisheries in Vava’U, Tonga. Master’s Thesis, McMaster University, Hamilton, OH, USA, 2010. [Google Scholar]
- Campos, F.Z. The Ichtyoarchaeology of Batanes Islands, Northern Philippines. Master’s Thesis, University of the Philippines Diliman, Quezon City, Philippines, 2009. [Google Scholar]
- Bouffandeau, L.; Béarez, P.; Bedford, S.; Valentin, F.; Spriggs, M.; Nolet, É. Fishing at Arapus-Mangaasi, Efate, Vanuatu (2800–2200 BP): New Methodological Approaches and Results. J. Archaeol. Sci. Rep. 2018, 18, 356–369. [Google Scholar] [CrossRef]
- Tyler, J.C. Osteology, Phylogeny, and Higher Classification of the Fishes of the Order Plectognathi (Tetraodontiformes); U.S. Dept. of Commerce, National Oceanic and Atmospheric Administration, National Marine Fisheries Service: Seattle, WA, USA, 1980.
- Fernandez-Jalvo, Y.; Andrews, P. Atlas of Taphonomic Identifications; Springer: Dordrecht, The Netherlands, 2017. [Google Scholar]
- Lebon, M. Caractérisation par Spectroscopie Infrarouge à Transformée de Fourier des Ossements Chauffés en Contexte Archéologique. Comparaison entre Référentiel Moderne et Matériel Fossile, Implications Diagénétiques. Ph.D. Thesis, Muséum National d’Histoire Naturelle, Paris, France, 2008. [Google Scholar]
- Willis, L.M.; Boehm, A.R. Fish Bones, Cut Marks, and Burial: Implications for Taphonomy and Faunal Analysis. J. Archaeol. Sci. 2014, 45, 20–25. [Google Scholar] [CrossRef]
- Ono, R. Ethno-Archaeology and Early Austronesian Fishing Strategies in near-Shore Environments. J. Polyn. Soc. 2010, 119, 269–314. [Google Scholar]
- Noguchi, T.; Arakawa, O. Tetrodotoxin—Distribution and Accumulation in Aquatic Organisms, and Cases of Human Intoxication. Mar. Drugs 2008, 6, 220–242. [Google Scholar] [CrossRef] [Green Version]
- Malpezzi, E.L.A.; De Freitas, J.; Rantin, F.T. Occurrence of Toxins, Other than Paralysing Type, in the Skin of Tetraodontiformes Fish. Toxicon 1997, 35, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Azman, A.M.N.; Samsur, M.; Othman, M. Distribution of Tetrodotoxin among Tissues of Pufferfish from Sabah and Sarawak Waters (Taburan Tetrodotoksin Antara Tisu Di Dalam Ikan Buntal Dari Perairan Sabah Dan Sarawak). Sains Malays. 2014, 43, 1003–1011. [Google Scholar]
- Elshaer, F.M. Comparative Histopathological Studies on Kidney and Liver of Rat Treated by Tetrodotoxin (TTX) Extracted from Gonads and Muscles of Porcupine Fish Species. Int. J. Fish. Aquat. Stud. 2016, 4, 355–360. [Google Scholar]
- Arakawa, O.; Hwang, D.F.; Taniyama, S.; Takani, T.H. Toxins of Pufferfish That Cause Human Intoxications. In Coastal Environmental and Ecosystem Issues of the East China Sea; Ishimatsu, A., Lie, H.-J., Eds.; Nagasaki University: Nagasaki, Japan, 2010; pp. 227–244. [Google Scholar]
- Farrag, M.M.S.; El-Far, A.M.; Aly, W.; AbouelFadl, K.Y.; Nasr-Allah, A.M.; El-Geddawy, M.-A.M.A.; Charo-Karisa, H. Nutritional Values vs. Toxicity Assessment of Pufferfish, Tetraodon Lineatus (Linnaeus, 1758), from Lake Nasser, Egypt. Egypt. J. Aquat. Res. 2022, 48, 53–59. [Google Scholar] [CrossRef]
- Khora, S.S.; Isa, J.; Yasumoto, T. Toxicity of Puffers from Okinawa, Japan. Nippon. Suisan Gakkaisha 1991, 57, 163–167. [Google Scholar] [CrossRef]
- Dalzell, P. The Role of Archaeological and Cultural-Historical Records in Long-Range Coastal Fisheries Resources Management Strategies and Policies in the Pacific Islands. Ocean. Coast. Manag. 1998, 40, 237–252. [Google Scholar] [CrossRef] [Green Version]
- Henrich, J.; Henrich, N. The Evolution of Cultural Adaptations: Fijian Food Taboos Protect against Dangerous Marine Toxins. Proc. R. Soc. Lond. B Biol. Sci. 2010, 277, rspb20101191. [Google Scholar] [CrossRef]
- Yong, Y.S.; Quek, L.S.; Lim, E.K.; Ngo, A. A Case Report of Puffer Fish Poisoning in Singapore. Case Rep. Med. 2013, 2013, e206971. [Google Scholar] [CrossRef]
- Kumar, N.P. Distribution, Extraction and Characterization of Tetrodotoxin (TTX) in Puffer Fishes from the South Andaman Sea. Ph.D. Thesis, Pondicherry University, Pondicherry, India, 2017. [Google Scholar]
- Tamele, I.J.; Silva, M.; Vasconcelos, V. The Incidence of Tetrodotoxin and Its Analogs in the Indian Ocean and the Red Sea. Mar. Drugs 2019, 17, 28. [Google Scholar] [CrossRef] [Green Version]
- Bagnis, R.A. La Ciguatera Aux Îles Marquises: Aspects Cliniques et Épidémiologiques. Bull. World Health Organ. 1973, 49, 67–73. [Google Scholar]
- Jal, S.; Khora, S.S. An Overview on the Origin and Production of Tetrodotoxin, a Potent Neurotoxin. J. Appl. Microbiol. 2015, 119, 907–916. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Oliver, J.D.; Alam, M.; Ali, A.; Waldor, M.K.; Qadri, F.; Martinez-Urtaza, J. Vibrio Spp. Infections. Nat. Rev. Dis. Prim. 2018, 4, 1–19. [Google Scholar] [CrossRef]
- Corrales, R.A.; Maclean, J.L. Impacts of Harmful Algae on Seafarming in the Asia-Pacific Areas. J. Appl. Phycol. 1995, 7, 151–162. [Google Scholar] [CrossRef]
- Monsalud, R.G.; Magbanua, F.O.; Tapay, L.M.; Hedreyda, C.T.; Olympia, M.S.D.; Migo, V.P.; Kurahashi, M.; Yokota, A. Identification of Pathogenic and Non-Pathogenic Vibrio Strains from Shrimp and Shrimp Farms in the Philippines. J. Gen. Appl. Microbiol. 2003, 49, 309–314. [Google Scholar] [CrossRef] [Green Version]
- Tendencia, E.A.; Lavilla-Pitogo, C.R. Bacterial Diseases. In Diseases of Cultured Groupers; Nagasawa, K., Cruz-Lacierda, E.R., Eds.; Southeast Asian Fisheries Development Center, Aquaculture Department: Iloilo, Philippines, 2004; pp. 19–28. [Google Scholar]
- Sampang, A.G. The Calamian Tagbanwa Ancestral Domain (Coron Island, Palawan, Philippines): Evaluation of Traditional Fishing Practices towards Biodiversity Conservation and Sustainability; World Fish Center: Los Baños, Philippines, 2007. [Google Scholar]
- Jawad, L.A. Dangerous Fishes of the Eastern and Southern Arabian Peninsula; Springer International Publishing: Berlin/Heidelberg, Germany, 2018; ISBN 978-3-319-57926-9. [Google Scholar]
- Gimlette, J.D. Malay Poisons and Charm Cures, 2nd ed.; J. & A. Churchill: London, UK, 1923. [Google Scholar]
- Roggatz, C.C.; Fletcher, N.; Benoit, D.M.; Algar, A.C.; Doroff, A.; Wright, B.; Wollenberg Valero, K.C.; Hardege, J.D. Saxitoxin and Tetrodotoxin Bioavailability Increases in Future Oceans. Nat. Clim. Chang. 2019, 9, 840–844. [Google Scholar] [CrossRef]
- Zong, Y. Mid-Holocene Sea-Level Highstand along the Southeast Coast of China. Quat. Int. 2004, 117, 55–67. [Google Scholar] [CrossRef]
- Abdelaziz, M.; Ibrahem, M.D.; Ibrahim, M.A.; Abu-Elala, N.M.; Abdel-moneam, D.A. Monitoring of Different Vibrio Species Affecting Marine Fishes in Lake Qarun and Gulf of Suez: Phenotypic and Molecular Characterization. Egypt. J. Aquat. Res. 2017, 43, 141–146. [Google Scholar] [CrossRef]
- Okita, K.; Yamazaki, H.; Sakiyama, K.; Yamane, H.; Niina, S.; Takatani, T.; Arakawa, O.; Sakakura, Y. Puffer Smells Tetrodotoxin. Ichthyol. Res. 2013, 60, 386–389. [Google Scholar] [CrossRef] [Green Version]
- Halstead, B.W. Poisonous Fishes and Their Relationship to Marine Food Resources in the Pacific Area. In Proceedings of the Eighth Pacific Science Congress of the Pacific Science Association, Quezon City, Philippines, 16–28 November 1953; National Research Council of the Philippines: Quezon City, Philippines, 1953; Volume III—Oceanography, pp. 321–330. [Google Scholar]
- Doherty, M.J. Captain Cook on Poison Fish. Neurology 2005, 65, 1788–1791. [Google Scholar] [CrossRef] [PubMed]
- Cisneros-Montemayor, A.M.; Pauly, D.; Weatherdon, L.V.; Ota, Y. A Global Estimate of Seafood Consumption by Coastal Indigenous Peoples. PLoS ONE 2016, 11, e0166681. [Google Scholar] [CrossRef]
- O’Connor, S.; Mahirta; Kealy, S.; Boulanger, C.; Maloney, T.R.; Hawkins, S.; Langley, M.C.; Kaharudin, H.A.F.; Suniarti, Y.; Husni, M.; et al. Kisar and the Archaeology of Small Islands in the Wallacean Archipelago. J. Isl. Coast. Archaeol. 2019, 14, 198–225. [Google Scholar] [CrossRef] [Green Version]
- Samper Carro, S.C.; O’Connor, S.; Louys, J.; Hawkins, S.; Mahirta, M. Human Maritime Subsistence Strategies in the Lesser Sunda Islands during the Terminal Pleistocene–Early Holocene: New Evidence from Alor, Indonesia. Quat. Int. 2016, 416, 64–79. [Google Scholar] [CrossRef]
- Byard, R.W. Death by Food. Forensic Sci. Med. Pathol. 2018, 14, 395–401. [Google Scholar] [CrossRef]
- Kan, S.K.; Chan, M.K.; David, P. Nine Fatal Cases of Puffer Fish Poisoning in Sabah, Malaysia. Med. J. Malays. 1987, 42, 199–200. [Google Scholar]
- Kanchanapongkul, J. Puffer Fish Poisoning: Clinical Features and Management Experience in 25 Cases. J. Med. Assoc. Thai. 2001, 84, 385–389. [Google Scholar] [PubMed]
- How, C.-K.; Chern, C.-H.; Huang, Y.-C.; Wang, L.-M.; Lee, C.-H. Tetrodotoxin Poisoning. Am. J. Emerg. Med. 2003, 21, 51–54. [Google Scholar] [CrossRef] [PubMed]
- El-Ganainy, A.A.; Sabrah, M.M.; Zaky, M.A. Biology and Toxicity of the Pufferfish Lagocephalus Sceleratus (GMELIN, 1789) from the Gulf of Suez. Egypt. J. Aquat. Res. 2006, 32, 283–297. [Google Scholar]
- Homaira, N.; Rahman, M.; Luby, S.P.; Rahman, M.; Haider, M.S.; Faruque, L.I.; Khan, D.; Parveen, S.; Gurley, E.S. Multiple Outbreaks of Puffer Fish Intoxication in Bangladesh, 2008. Am. J. Trop. Med. Hyg. 2010, 83, 440–444. [Google Scholar] [CrossRef]
- Islam, Q.T.; Razzak, M.A.; Islam, M.A.; Bari, M.I.; Basher, A.; Chowdhury, F.R.; Sayeduzzaman, A.B.M.; Ahasan, H.A.M.N.; Faiz, M.A.; Arakawa, O.; et al. Puffer Fish Poisoning in Bangladesh: Clinical and Toxicological Results from Large Outbreaks in 2008. Trans. R. Soc. Trop. Med. Hyg. 2011, 105, 74–80. [Google Scholar] [CrossRef]
- Cordova, C. Puffer Fish for Breakfast Kills Woman, Sends Husband, 6 Kids to Hospital in Cebu. Manila Buletin. 2022. Available online: https://mb.com.ph/2022/08/04/puffer-fish-for-breakfast-kills-woman-sends-husband-6-kids-to-hospital-in-cebu/ (accessed on 1 September 2022).
- Amazona, W.M. 3 Dead after Eating Puffer Fish in Tabango Town. CNN Philippines. 2020. Available online: http://www.cnnphilippines.com/news/2020/12/29/three-dead-food-poisoning-puffer-fish-tabango.html (accessed on 1 September 2022).
- Diestro, D. 17 Nalason Sa Pagkain Ng “Butete” Sa Ozamiz. ABS-CBN News. 2019. Available online: https://news.abs-cbn.com/news/08/13/19/17-nalason-sa-pagkain-ng-butete-sa-ozamiz (accessed on 1 September 2022).
- Pawlik, A.; Crozier, R.; Fuentes, R.; Wood, R.; Piper, P. Burial Traditions in Early Mid-Holocene Island Southeast Asia: New Evidence from Bubog-1, Ilin Island, Mindoro Occidental. Antiquity 2019, 93, 901–918. [Google Scholar] [CrossRef] [Green Version]
- Begossi, A. Food Taboos at Búzios Island (Brazil): Their Significance and Relation to Folk Medicine. J. Ethnobiol. 1992, 12, 117–139. [Google Scholar]
- Seixas, C.; Begossi, A. Ethnozoology of Fishing Communities from Ilha Grande (Atlantic Forest Coast, Brazil). J. Ethnobiol. 2001, 21, 107–135. [Google Scholar]
- Ramires, M.; Rotundo, M.M.; Begossi, A. The Use of Fish in Ilhabela (São Paulo/Brazil): Preferences, Food Taboos and Medicinal Indications. Biota Neotrop. 2012, 12, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Yakop Yahya, M.B.; Arif, M.R.B.M. Ikan Buntal Kuning (Lagocephalus lunaris), Sebagai Produk Pelancongan Di Betong, Sarawak (Ikan Buntal Kuning (Lagocephalus lunaris) as a Tourism Product in Betong, Sarawak). Jati J. Southeast Asian Stud. 2008, 13, 245–263. [Google Scholar]
- Intoh, M. Reconnaissance Archaeological Research on Ngulu Atoll in the Western Caroline Islands. Asian Perspect. 1981, 24, 69–80. [Google Scholar]
- Leach, F.B.; Davidson, J.M. The Quest for the Rainbow Runner: Prehistoric Fishing on Kapingamarangi and Nukuoro Atolls, Micronesia. Micronesica 1988, 21, 1–22. [Google Scholar]
- Amesbury, J.R.; Hunter-Anderson, R.L. Review of Archaeological and Historical Data Concerning Reef Fishing in the US Flag Islands of Micronesia: Guam and the Northern Mariana Islands; Western Pacific Regional Fishery Management Council: Barrigada, Guam, 2003. [Google Scholar]
- Ono, R.; Morrison, A.; Addison, D. Prehistoric Marine Resource Use in the Indo-Pacific Regions; Terra Australis; ANU Press: Canberra, Australia, 2009; ISBN 978-1-925021-26-4. [Google Scholar]
- Ono, R.; Intoh, M. Island of Pelagic Fishermen: Temporal Changes in Prehistoric Fishing on Fais, Micronesia. J. Isl. Coast. Archaeol. 2011, 6, 255–286. [Google Scholar] [CrossRef]
- Ono, R.; Clark, G. A 2500-Year Record of Marine Resource Use on Ulong Island, Republic of Palau. Int. J. Osteoarchaeol. 2012, 22, 637–654. [Google Scholar] [CrossRef]
- Jones, S. Human Impacts on Ancient Marine Environments of Fiji’s Lau Group: Current Ethnoarchaeological and Archaeological Research. J. Isl. Coast. Archaeol. 2007, 2, 239–244. [Google Scholar] [CrossRef]
- Clark, G.; Anderson, A. The Early Prehistory of Fiji; Terra Australis; ANU Press: Canberra, Australia, 2009; Volume 31, ISBN 978-1-921666-07-0. [Google Scholar]
- Allen, M.S.; Schubel, S.E. Recent Archaeological Research on Aitutaki, Southern Cooks: The Moturakau Shelter. J. Polyn. Soc. 1990, 99, 265–295. [Google Scholar]
- Ono, R.; Addison, D.J. Ethnoecology and Tokelauan Fishing Lore from Atafu Atoll, Tokelau. SPC Tradit. Mar. Resour. Manag. Knowl. Inf. Bull. 2009, 26, 3–22. [Google Scholar]
- Weisler, M.I.; Green, R.C. Mangareva Fishing Strategies in Regional Context: An Analysis of Fish Bones from Five Sites Excavated in 1959. J. Pac. Archaeol. 2013, 4, 73–89. [Google Scholar]
- Cannon, A.; Wildenstein, R.; Cannon, D.Y.; Burley, D.V. Consistency and Variation in the Focus, Intensity and Archaeological Histories of Lapita and Post-Lapita Fisheries in Ha’apai, Kingdom of Tonga. J. Isl. Coast. Archaeol. 2018, 14, 515–540. [Google Scholar] [CrossRef]
- O’Connor, S.; Ono, R.; Clarkson, C. Pelagic Fishing at 42,000 Years before the Present and the Maritime Skills of Modern Humans. Science 2011, 334, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Colley, S.M. The Analysis and Interpretation of Archaeological Fish Remains. Archaeol. Method Theory 1990, 2, 207–253. [Google Scholar]
- Maxwell, D. Beyond Maritime Symbolism: Toxic Marine Objects from Ritual Contexts at Tikal. Anc. Mesoam. 2000, 11, 91–98. [Google Scholar] [CrossRef]
- Davis, E.W. The Ethnobiology of the Haitian Zombi. J. Ethnopharmacol. 1983, 9, 85–104. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.W. Preparation of the Haitian Zombi Poison. Bot. Mus. Leafl. Harv. Univ. 1983, 29, 139–149. [Google Scholar] [CrossRef]
- Benedek, C.; Rivier, L. Evidence for the Presence of Tetrodotoxin in a Powder Used in Haiti for Zombification. Toxicon 1989, 27, 473–480. [Google Scholar] [CrossRef]
- Efthimiou, C.J.; Gandhi, S. Ghosts, Vampires and Zombies—Cinema Fiction vs. Physics Reality. Skept. Inq. 2007, 31, 1–11. [Google Scholar]
- Glowacki, M. Food of the Gods or Mere Mortals? Hallucinogenic Spondylus and Its Interpretive Implications for Early Andean Society. Antiquity 2005, 79, 257–268. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Muscle | Skin | Testis | Ovary | Liver | Intestine | Blood | |||
|---|---|---|---|---|---|---|---|---|---|
| Diodontidae | Chilomycterus | affinis | ◌ | ◌ | - | ◌ | ◌ | ◌ | - |
| reticulatus | - | - | - | - | - | - | - | ||
| Cyclichthys | orbicularis | - | - | - | - | - | - | - | |
| spilostylus | - | - | - | - | - | - | - | ||
| Diodon | eydouxii | - | - | - | - | - | - | - | |
| holocanthus | ◌ | ◌ | - | ◌ | ◌ | ◌ | - | ||
| hystrix | ◌ | ◌ | - | ◌ | ◌ | ◌ | - | ||
| liturosus | - | - | - | - | - | - | - | ||
| Tetraodontidae | Arothron | hispidus | ● | ✖ | ◌ | ◎ | ◎ | ◎ | - |
| firmamentum | ◌ | ● | ◌ | ◎ | ◌ | ◌ | - | ||
| manilensis | ● | ◎ | - | ◎ | ● | ● | - | ||
| mappa | ● | ● | - | ◎ | ● | ● | - | ||
| nigropunctatus | ◎ | ◎ | - | ◎ | ◎ | ◎ | - | ||
| stellatus | ● | ● | ✖ | ✖ | ● | ● | - | ||
| Canthigaster | rivulata | ◌ | ◎ | - | ◌ | ● | ● | - | |
| Chelonodon | patoca | ◎ | ✖ | ◎ | ◎ | ◎ | ✖ | - | |
| Lagocephalus | gloveri | ◌ | ◌ | ◌ | ◌ | ◌ | ◌ | - | |
| inermis | ◌ | ◌ | ◌ | ◌ | ◎ | ◌ | - | ||
| lunaris | ◉ | ◎ | ◌ | ◌ | ◌ | ◌ | - | ||
| wheeleri | ◌ | ◌ | ◌ | ◌ | ◌ | ◌ | - | ||
| sceleratus | ● | ● | - | ✖ | ● | ◎ | - | ||
| Takifugu | chinensis | - | - | - | ✖ | ✖ | - | - | |
| chrysops | ◌ | ◎ | ◌ | ◎ | ◎ | ● | ◌ | ||
| exascurus | ◌ | ◎ | ◌ | ✖ | ◎ | - | - | ||
| flavidus | ● | ◎ | ◎ | ✖ | ✖ | ◎ | - | ||
| niphobles | ● | ◎ | ● | ✖ | ✖ | ✖ | - | ||
| obscurus | ◌ | ◎ | ◌ | ✖ | ◎ | ◎ | - | ||
| pardalis | ◌ | ◎ | ● | ✖ | ✖ | ◎ | ◌ | ||
| poecilonotus | ● | ◎ | ◎ | ✖ | ✖ | ◎ | - | ||
| porphyreus | ◌ | ◎ | ◌ | ✖ | ✖ | ◎ | - | ||
| pseudommus | ◌ | ● | ◌ | ✖ | ● | ● | - | ||
| rubripes | ◌ | ◌ | ◌ | ✖ | ◎ | ● | ◌ | ||
| snyderi | ● | ◎ | ◌ | ✖ | ✖ | ◎ | - | ||
| stictonotus | ◌ | ● | ◌ | ◎ | ◎ | ◌ | - | ||
| vermicularis | ◌ | ◎ | ◌ | ◎ | ◎ | ● | - | ||
| xanthopterus | ◌ | ◌ | ◌ | ◎ | ◎ | ● | - | ||
| Sphoeroides | pachygaster | ◌ | ◌ | ◌ | ◌ | ◌ | ◌ | - | |
| Tetraodon | alboreticulatus | ● | ● | - | ✖ | ● | ◎ | - | |
| nigrovridis | ● | ◎ | - | - | ◌ | ● | - | ||
| steindachneri | ◌ | ◌ | - | - | ◌ | ◌ | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boulanger, C.; Pawlik, A.; O’Connor, S.; Sémah, A.-M.; Reyes, M.C.; Ingicco, T. The Exploitation of Toxic Fish from the Terminal Pleistocene in Maritime Southeast Asia: A Case Study from the Mindoro Archaeological Sites, Philippines. Animals 2023, 13, 2113. https://doi.org/10.3390/ani13132113
Boulanger C, Pawlik A, O’Connor S, Sémah A-M, Reyes MC, Ingicco T. The Exploitation of Toxic Fish from the Terminal Pleistocene in Maritime Southeast Asia: A Case Study from the Mindoro Archaeological Sites, Philippines. Animals. 2023; 13(13):2113. https://doi.org/10.3390/ani13132113
Chicago/Turabian StyleBoulanger, Clara, Alfred Pawlik, Sue O’Connor, Anne-Marie Sémah, Marian C. Reyes, and Thomas Ingicco. 2023. "The Exploitation of Toxic Fish from the Terminal Pleistocene in Maritime Southeast Asia: A Case Study from the Mindoro Archaeological Sites, Philippines" Animals 13, no. 13: 2113. https://doi.org/10.3390/ani13132113