
Antioxidant Activity of Different Tissues from Rabbits Fed Dietary Bovine Colostrum Supplementation

 , , ,
, , ,  , , , , ,
, , , , ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals Diets and Samples Collection
2.2. Total Phenol Content and Antioxidant Activity of Bovine Colostrum and Experimental Diets
2.2.1. Sample Treatment
2.2.2. Total Phenol Content
2.2.3. Antioxidant Activities (DPPH, ABTS, ORAC Assays)
2.3. Antioxidant Enzymes Activity Determination
2.4. RNA Extraction and cDNA Synthesis
2.5. Real Time RT-qPCR
2.6. Statistical Analyses
3. Results
3.1. Total Phenol Content and Antioxidant Activity of Bovine Colostrum and Experimental Diets
3.2. Antioxidant Enzymes Activity in Plasma
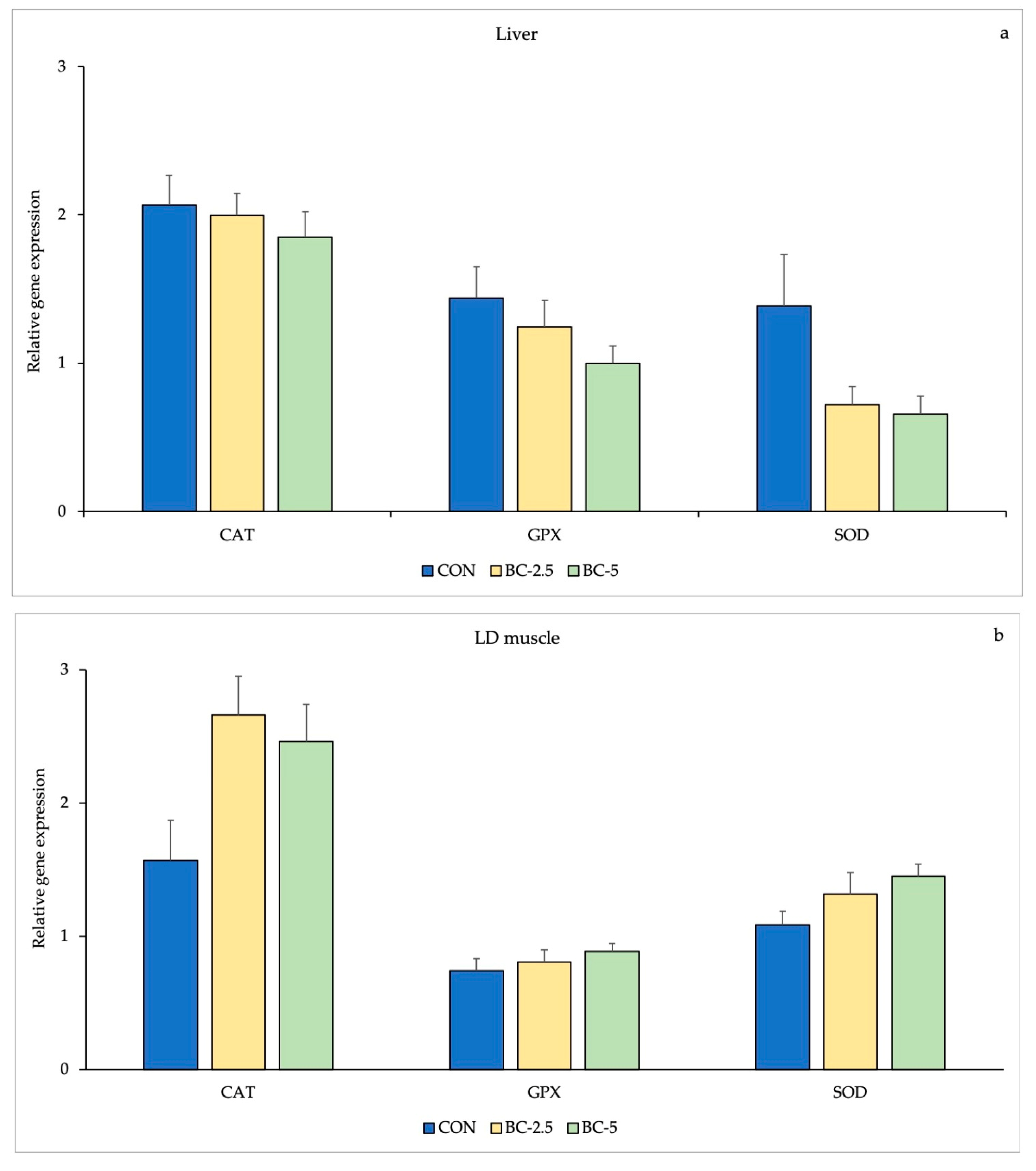
3.3. Antioxidant Enzymes Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mann, S.; Curone, G.; Chandler, T.L.; Moroni, P.; Cha, J.; Bhawal, R.; Zhang, S. Heat treatment of bovine colostrum: I. Effects on bacterial and somatic cell counts, immunoglobulin, insulin, and IGF-I concentrations, as well as the colostrum proteome. J. Dairy Sci. 2020, 103, 9368–9383. [Google Scholar] [CrossRef]
- Gomes, R.D.; Anaya, K.; Galdino, A.B.; Oliveira, J.P.; Gama, M.A.; Medeiros, C.A.; Gavioli, E.C.; Porto, A.L.F.; Rangel, A.H. Bovine colostrum: A source of bioactive compounds for prevention and treatment of gastrointestinal disorders. NFS J. 2021, 25, 1–11. [Google Scholar] [CrossRef]
- Bagwe-Parab, S.; Yadav, P.; Kaur, G.; Tuli, H.S.; Buttar, H.S. Therapeutic applications of human and bovine colostrum in the treatment of gastrointestinal diseases and distinctive cancer types: The current evidence. Front. Pharmacol. 2020, 11, 1100. [Google Scholar] [CrossRef] [PubMed]
- Taillon, C.; Andreasen, A. Veterinary nutraceutical medicine. Can. Vet. J. 2000, 41, 231. [Google Scholar] [PubMed]
- Boudry, C.; Dehoux, J.; Wavreille, J.; Portetelle, D.; Théwis, A.; Buldgen, A. Effect of a bovine colostrum whey supplementation on growth performance, faecal Escherichia coli population and systemic immune response of piglets at weaning. Animal 2008, 2, 730–737. [Google Scholar] [CrossRef] [Green Version]
- Satyaraj, E.; Reynolds, A.; Pelker, R.; Labuda, J.; Zhang, P.; Sun, P. Supplementation of diets with bovine colostrum influences immune function in dogs. Br. J. Nutr. 2013, 110, 2216–2221. [Google Scholar] [CrossRef] [Green Version]
- Afzal, I.; Khan, A.A.; Banday, M.T.; Manzoor, A.; Untoo, M.; Khurshid, A.; Bashir, S.U.; Rovida, M. Effect of incorporation of bovine colostrum in broiler chicken diets on body weight, mortality pattern and economics of production. J. Entomol. Zool. Sci. 2018, 6, 711–714. [Google Scholar]
- Corino, C.; Pastorelli, G.; Pantaleo, L.; Oriani, G.; Salvatori, G. Improvement of color and lipid stability of rabbit meat by dietary supplementation with vitamin E. Meat Sci. 1999, 52, 285–289. [Google Scholar] [CrossRef]
- Crovato, S.; Pinto, A.; Di Martino, G.; Mascarello, G.; Rizzoli, V.; Marcolin, S.; Ravarotto, L. Purchasing habits, sustainability perceptions, and welfare concerns of Italian consumers regarding rabbit meat. Foods 2022, 11, 1205. [Google Scholar] [CrossRef]
- Meineri, G.; Giacobini, M.; Forneris, G. Evaluation of physiological parameters of the plasma oxidative status in rabbits. J. Appl. Anim. Res. 2017, 45, 315–319. [Google Scholar] [CrossRef] [Green Version]
- Tremellen, K. Oxidative stress and male infertility—A clinical perspective. Hum. Reprod. Update 2008, 14, 243–258. [Google Scholar] [CrossRef]
- Sikiru, A.B.; Arangasamy, A.; Alemede, I.C.; Egena, S.S.A.; Bhatta, R. Dietary supplementation effects of Chlorella vulgaris on performances, oxidative stress status and antioxidant enzymes activities of prepubertal New Zealand White rabbits. Bull. Natl. Res. Cent. 2019, 43, 162. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.C.; Ho, J.A.A.; Shieh, M.C.; Lu, I.W. Antioxidant and antiproliferative activities of Spirulina and Chlorella water extracts. J. Agric. Food Chem. 2005, 53, 4207–4212. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Z.; Xu, C.L.; An, Z.H.; Liu, J.X.; Feng, J. Effect of dietary bovine lactoferrin on performance and antioxidant status of piglets. Anim. Feed Sci. Technol. 2008, 140, 326–336. [Google Scholar] [CrossRef]
- Yang, M.; Zou, Y.; Wu, Z.H.; Li, S.L.; Cao, Z.J. Colostrum quality affects immune system establishment and intestinal development of neonatal calves. J. Dairy Sci. 2015, 98, 7153–7163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quigley, J.D.; Lago, A.; Chapman, C.; Erickson, P.; Polo, J. Evaluation of the Brix refractometer to estimate immunoglobulin G concentration in bovine colostrum. J. Dairy Sci. 2013, 96, 1148–1155. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Ravento´s, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Saltarelli, R.; Palma, F.; Gioacchini, A.M.; Calcabrini, C.; Mancini, U.; De Bellis, R.; Stocchi, V.; Potenza, L. Phytochemical composition, antioxidant and antiproliferative activities and effects on nuclear DNA of ethanolic extract from an Italian mycelial isolate of Ganoderma lucidum. J. Ethnopharmacol. 2019, 231, 464–473. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Sicari, V.; Pellicano, T.; Xiao, J.; Poiana, M.; Tundis, R. Comparative analysis of chemical composition, antioxidant and anti-proliferative activities of Italian Vitis vinifera by-products for a sustainable agro-industry. Food Chem. Toxicol. 2019, 127, 127–134. [Google Scholar] [CrossRef]
- De Bellis, R.; Piacentini, M.P.; Meli, M.A.; Mattioli, M.; Menotta, M.; Mari, M.; Valentini, L.; Palomba, L.; Desideri, D.; Chiarantini, L. In vitro effects on calcium oxalate crystallization kinetics and crystal morphology of an aqueous extract from Ceterach officinarum: Analysis of a potential antilithiatic mechanism. PLoS ONE 2019, 14, e0218734. [Google Scholar] [CrossRef] [Green Version]
- Kehoe, S.I.; Jayarao, B.M.; Heinrichs, A.J. A survey of bovine colostrum composition and colostrum management practices on Pennsylvania dairy farms. J. Dairy Sci. 2007, 90, 4108–4116. [Google Scholar] [CrossRef] [Green Version]
- Playford, R.J.; Weiser, M.J. Bovine colostrum: Its constituents and uses. Nutrients 2021, 13, 265. [Google Scholar] [CrossRef] [PubMed]
- Elfstrand, L.; Lindmark-Månsson, H.; Paulsson, M.; Nyberg, L.; Åkesson, B. Immunoglobulins, growth factors and growth hormone in bovine colostrum and the effects of processing. Int. Dairy J. 2002, 12, 879–887. [Google Scholar] [CrossRef]
- Galiot, L.; Audet, I.; Ouattara, B.; Bissonnette, N.; Talbot, G.; Raymond, F.; Deschesnes, T.; Lapointe, J.; Lo Verso, L.; Lessard, M.; et al. Effect of the administration of copper, vitamins A and D and bovine colostrum on performances, antioxidant and micronutrients status and microbiome in lactating piglets on a commercial farm. Livest. Sci. 2021, 251, 104609. [Google Scholar] [CrossRef]
- Dzik, S.; Miciński, B.; Aitzhanova, I.; Miciński, J.; Pogorzelska, J.; Beisenov, A.; Kowalski, I.M. Properties of bovine colostrum and the possibilities of use. Pol. Ann. Med. 2017, 24, 295–299. [Google Scholar] [CrossRef]
- McGrath, B.A.; Fox, P.F.; McSweeney, P.L.; Kelly, A.L. Composition and properties of bovine colostrum: A review. Dairy Sci. Technol. 2016, 96, 133–158. [Google Scholar] [CrossRef] [Green Version]
- Kalyan, S.; Meena, S.; Kapila, S.; Yadav, R.; Deshwal, G.K. In vitro assessment of antioxidative potential of goat milk, casein and its hydrolysates: Comparison of goat milk with bovine and buffalo milk. Res. Sq. 2021, 1–19. [Google Scholar] [CrossRef]
- Tijerina-Sáenz, A.; Innis, S.M.; Kitts, D.D. Antioxidant capacity of human milk and its association with vitamins A and E and fatty acid composition. Acta Paediatr. 2009, 98, 1793–1798. [Google Scholar] [CrossRef] [Green Version]
- Aziz, M.A.; Diab, A.S.; Mohammed, A.A. Antioxidant categories and mode of action. In Antioxidants; Shalaby, E., Ed.; IntechOpen: London, UK, 2019; pp. 3–22. [Google Scholar]
- Alía, M.; Horcajo, C.; Bravo, L.; Goya, L. Effect of grape antioxidant dietary fiber on the total antioxidant capacity and the activity of liver antioxidant enzymes in rats. Nutr. Res. 2003, 23, 1251–1267. [Google Scholar] [CrossRef] [Green Version]
- Mokhtarzadeh, S.; Nobakht, A.; Mehmannavaz, Y.; Palangi, V.; Eseceli, H.; Lackner, M. Impacts of continuous and intermittent use of bovine colostrum on laying Japanese quails: Egg performance and traits, blood biochemical and antioxidant status. Animals 2022, 12, 2811. [Google Scholar] [CrossRef]
- Deng, K.P.; Fan, Y.X.; Ma, T.W.; Wang, Z.; TanTai, W.J.; Nie, H.T.; Guo, Y.X.; Sun, L.W.; Wang, F. Carcass traits, meat quality, antioxidant status and antioxidant gene expression in muscle and liver of Hu lambs fed perilla seed. J. Anim. Physiol. Anim. Nutr. 2018, 102, e828–e837. [Google Scholar] [CrossRef]
- Placha, I.; Bacova, K.; Plachy, L. Current knowledge on the bioavailability of thymol as a feed additive in humans and animals with a focus on rabbit metabolic processes. Animals 2022, 12, 1131. [Google Scholar] [CrossRef] [PubMed]
- Bacova, K.; Eglseer, K.Z.; Räuber, G.K.; Chrastinova, L.; Laukova, A.; Takacsova, M.; Simonova, M.P.; Placha, I. Effect of sustained administration of thymol on its bioaccessibility and bioavailability in rabbits. Animals 2021, 11, 2595. [Google Scholar] [CrossRef] [PubMed]
- Galinier, A.; Carriere, A.; Fernandez, Y.; Carpéné, C.; André, M.; Caspar-Bauguil, S.; Thouvenot, J.P.; Périquet, B.; Pénicaud, L.; Casteilla, L. Adipose tissue proadipogenic redox changes in obesity. J. Biol. Chem. 2006, 281, 12682–12687. [Google Scholar] [CrossRef] [Green Version]
- Elsasser, T.H.; Caperna, T.J.; Li, C.J.; Kahl, S.; Sartin, J.L. Critical control points in the impact of the proinflammatory immune response on growth and metabolism. J. Anim. Sci. 2008, 86, E105–E125. [Google Scholar] [CrossRef] [PubMed]
- Vincent, H.K.; Innes, K.E.; Vincent, K.R. Oxidative stress and potential interventions to reduce oxidative stress in overweight and obesity. Diabetes Obes. Metab. 2007, 9, 813–839. [Google Scholar] [CrossRef]
- Noeman, S.A.; Hamooda, H.E.; Baalash, A.A. Biochemical study of oxidative stress markers in the liver, kidney and heart of high fat diet induced obesity in rats. Diabetol. Metab. Syndr. 2011, 3, 17. [Google Scholar] [CrossRef] [Green Version]
- Moisá, S.J.; Shike, D.W.; Graugnard, D.E.; Rodriguez-Zas, S.L.; Everts, R.E.; Lewin, H.A.; Faulkner, D.B.; Berger, L.L.; Loor, J.J. Bioinformatics analysis of transcriptome dynamics during growth in angus cattle longissimus muscle. Bioinform. Biol. Insights 2013, 7, BBI-S12328. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2017, 114, 1752–1761. [Google Scholar] [CrossRef]
- Bashan, N.; Kovsan, J.; Kachko, I.; Ovadia, H.; Rudich, A. Positive and negative regulation of insulin signaling by reactive oxygen and nitrogen species. Physiol. Rev. 2009, 89, 27–71. [Google Scholar] [CrossRef] [Green Version]
- Avery, S.V. Molecular targets of oxidative stress. Biochem. J. 2011, 434, 201–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellogg, E.W.; Fridovich, I. Superoxide dismutase in the rat and mouse as a function of age and longevity. J. Gerontol. 1976, 31, 405–408. [Google Scholar] [CrossRef]
- Tolmasoff, J.M.; Ono, T.; Cutler, R.G. Superoxide dismutase: Correlation with life-span and specific metabolic rate in primate species. Proc. Natl. Acad. Sci. USA 1980, 77, 2777–2781. [Google Scholar] [CrossRef] [Green Version]
- Mattioli, S.; Collodel, G.; Signorini, C.; Cotozzolo, E.; Noto, D.; Cerretani, D.; Micheli, L.; Fiaschi, A.I.; Brecchia, G.; Menchetti, L.; et al. Tissue antioxidant status and lipid peroxidation are related to dietary intake of n-3 polyunsaturated acids: A rabbit model. Antioxidants 2021, 10, 681. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Korshom, M.A.; Mandour, A.A.; El-Bessoumy, A.A.; El-Sayed, Y.S. Hepatoprotective efficacy of Nigella sativa seeds dietary supplementation against lead acetate-induced oxidative damage in rabbit–purification and characterization of glutathione peroxidase. Biomed. Pharmacother. 2017, 89, 711–718. [Google Scholar] [CrossRef]
- Aitken, S.L.; Karcher, E.L.; Rezamand, P.; Gandy, J.C.; VandeHaar, M.J.; Capuco, A.V.; Sordillo, L.M. Evaluation of antioxidant and proinflammatory gene expression in bovine mammary tissue during the periparturient period. J. Dairy Sci. 2009, 92, 589–598. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Maiorino, M.; Roveri, A.; Ursini, F. Phospholipid hydroperoxide glutathione peroxidase: Specific activity in tissues of rats of different age and comparison with other glutathione peroxidases. Biochim. Biophys. Acta-Lipids Lipid Metab. 1989, 1006, 140–143. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.L. Modulation of skeletal muscle antioxidant defense by exercise: Role of redox signaling. Free Radic. Biol. Med. 2008, 44, 142–152. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Ingredients | Basal Diet | Experimental Diets | |
|---|---|---|---|
| CON | BC-2.5 | BC-5 | |
| Dehydrated alfalfa meal | 32.40 | 32.40 | 32.40 |
| Barley | 22.40 | 19.90 | 17.40 |
| Wheat bran | 20.50 | 20.50 | 20.50 |
| Sunflower meal | 8.00 | 8.00 | 8.00 |
| Soybean meal | 4.00 | 4.00 | 4.00 |
| Cane molasses | 3.00 | 3.00 | 3.00 |
| Carob pods | 2.70 | 2.70 | 2.70 |
| Wheat meal | 2.50 | 2.50 | 2.50 |
| Calcium Carbonate | 2.00 | 2.00 | 2.00 |
| Vitamin–mineral premix 1 | 1.60 | 1.60 | 1.60 |
| Soybean oil | 0.50 | 0.50 | 0.50 |
| Sodium chloride | 0.40 | 0.40 | 0.40 |
| Bovine colostrum | - | 2.50 | 5.00 |
| Chemical composition 2 | |||
| Dry matter | 92.34 | 91.71 | 91.69 |
| Crude protein | 14.82 | 14.76 | 15.23 |
| Ether extract | 2.79 | 2.95 | 3.02 |
| Ash | 7.04 | 7.23 | 7.62 |
| NDF | 40.00 | 36.81 | 35.79 |
| ADF | 27.04 | 24.92 | 24.31 |
| ADL | 12.02 | 10.03 | 9.11 |
| Gene | Forward Primer | Reverse Primer | Tm F (°C) | Tm R (°C) | Amplicon Size (bp) |
|---|---|---|---|---|---|
| GAPDH | GTCAAGGCTGAGAACGGGAA | TCTCCATGGTGGTGAAGACG | 59.4 | 59.4 | 142 |
| ACTB | ACATGGAGAAGATCTGGCAC | GCGTGTTGAACGTCTCGAAC | 57.3 | 59.4 | 147 |
| SOD | TCGGGAGATATGTCCGTC | GACACCACAGGCCAAACG | 56.0 | 58.2 | 126 |
| CAT | GCTGAGATTGAACAGTTGGC | GGTGAGTATCGGGATAGGAG | 57.3 | 59.4 | 109 |
| GPx | CAGTTTGGGCATCAGGAGAAC | GCATGAAGTTGGGCTCGAAC | 59.8 | 59.4 | 94 |
| CON | BC-2.5 | BC-5 | p-Value | |
|---|---|---|---|---|
| TPC (mg GAE/g) | 3.85 ± 0.15 | 4.09 ± 0.27 | 3.84 ± 0.08 | ns |
| DPPH (μmol TE/g) | 11.13 ± 0.20 | 11.14 ± 0.19 | 11.16 ± 0.17 | ns |
| ABTS (μmol TE/g) | 36.42 ± 1.84 | 34.88 ± 0.23 | 36.12 ± 0.42 | ns |
| ORAC (μmol TE/g) | 113.00 ± 3.8 | 136.3 ± 4.5 | 150.70 ± 5.8 | <0.001 |
| CON | BC-2.5 | BC-5 | p-Value | |
|---|---|---|---|---|
| Plasma CAT (U/mL) | 11.68 ± 1.86 | 9.82 ± 1.75 | 7.66 ± 2.11 | 0.347 |
| Plasma GPx (U/mL) | 196.21 ± 16.67 | 253.46 ± 33.39 | 237.27 ± 21.85 | 0.265 |
| Plasma SOD (U/L) | 91.72 ± 8.69 | 87.38 ± 7.83 | 77.82 ± 6.85 | 0.449 |
| Liver | LD muscle | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|
| CON | BC-2.5 | BC-5 | CON | BC-2.5 | BC-5 | Diet | Tissue | |
| CAT | 2.067 ± 0.79 | 1.995 ± 0.56 | 1.786 ± 0.35 | 1.568 ± 1.08 | 2.659 ± 1.28 | 2.459 ± 1.18 | 0.237 | 0.257 |
| GPx | 1.439 ± 0.72 | 1.244 ± 0.61 | 0.997 ± 0.24 | 0.739 ± 0.31 | 0.806 ± 0.35 | 0.886 ± 0.31 | 0.606 | 0.001 |
| SOD | 1.384 ± 1.25 | 0.722 ± 0.15 | 0.657 ± 0.15 | 1.086 ± 0.40 | 1.315 ± 0.44 | 1.449 ± 0.37 | 0.471 | 0.022 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serra, V.; Castrica, M.; Agradi, S.; Curone, G.; Vigo, D.; Di Giancamillo, A.; Modina, S.C.; Riva, F.; Balzaretti, C.M.; De Bellis, R.; et al. Antioxidant Activity of Different Tissues from Rabbits Fed Dietary Bovine Colostrum Supplementation. Animals 2023, 13, 850. https://doi.org/10.3390/ani13050850
Serra V, Castrica M, Agradi S, Curone G, Vigo D, Di Giancamillo A, Modina SC, Riva F, Balzaretti CM, De Bellis R, et al. Antioxidant Activity of Different Tissues from Rabbits Fed Dietary Bovine Colostrum Supplementation. Animals. 2023; 13(5):850. https://doi.org/10.3390/ani13050850
Chicago/Turabian StyleSerra, Valentina, Marta Castrica, Stella Agradi, Giulio Curone, Daniele Vigo, Alessia Di Giancamillo, Silvia Clotilde Modina, Federica Riva, Claudia Maria Balzaretti, Roberta De Bellis, and et al. 2023. "Antioxidant Activity of Different Tissues from Rabbits Fed Dietary Bovine Colostrum Supplementation" Animals 13, no. 5: 850. https://doi.org/10.3390/ani13050850