1. Introduction
The giant freshwater prawn
Macrobrachium rosenbergii is one of the most cultured crustacean species worldwide due to its nutritional value, good meat quality, and high growth rate [
1,
2]. The production of
M. rosenbergii is considered economical and more environmentally sustainable than that of other intensively reared species of crustaceans, as low stocking densities are often used [
3,
4]. However, this aquaculture sector is still challenged by the species’ dependence on diets containing high percentages of unsustainable and expensive marine-derived ingredients, such as fish meal (FM), which necessary to ensure proper growth and welfare [
5,
6].
In recent decades, great efforts have been made within
M. rosenbergii farming to reduce the costs and the environmental footprint of dietary formulations by replacing FM with alternative protein sources of both plant and animal origin [
7,
8,
9]. Plant-derived ingredients were first sought because of their wider availability and competitive costs [
10]. In particular, it was first demonstrated that soybean meal could partially or totally replace FM in diets for
M. rosenbergii without a negative effect on growth [
11,
12]. However, further studies on
M. rosenbergii that coupled zootechnical performances with protein metabolism, antioxidant capacity, and intestinal microbiota demonstrated that FM replacement with soybean meal should not exceed 50% [
9,
13].
Monitoring the overall welfare of crustaceans through their physiological responses is of primary importance when new aquafeed ingredients are tested, especially for species such as the giant freshwater prawn that are frequently subject to infectious diseases with consequent economic losses[
14,
15]. In this regard, feed additives or probiotics that are able to enforce the resistance to pathological and environmental stresses of
M. rosenbergii have been successfully used as a better alternative to chemicals and antibiotics [
16,
17,
18]. Furthermore, to increase aquafeed sustainability, great attention has been given to alternative ingredients; for example, microbial biomass and insect meal, in addition to their immunostimulant features, may offer the possibility of replacing high amounts of FM as a major protein source [
19,
20]. In this context, promising results on both the growth performances and the immune responses of
M. rosenbergii have been obtained by partially replacing FM with microbial biomass, such as yeasts [
6] and microalgae [
21,
22,
23], or by including insect meal in diets totally deprived of FM [
24]. However, the high production costs and prices of microalgae and the fatty-acids (FA) profile of insects represent the main bottleneck that limits a wider use of those ingredients in aquaculture [
25,
26]. However, some insect species, such as the black soldier fly (
Hermetia illucens), which have the proper protein tenor and amino acid profile and an appreciable content of bioactive molecules (i.e., lauric acid, chitin, and antimicrobial peptides) with antimicrobial and immunostimulatory features [
27,
28,
29], are able to modulate their nutritional profile in relation to the growth substrate [
30,
31]. In this regard, the enrichment of the
H. illucens growth substrate with microbial biomass represents an efficient method of exploiting the beneficial properties of both ingredients [
32,
33]. In particular, a recent study demonstrated that the 15% (
w/
w) inclusion of dried biomass from
Arthrospira platensis (spirulina) in insects’ growth substrate resulted in a successful transfer of antioxidant molecules (tocopherols and carotenoids) of this cyanobacterium to
H. illucens larvae, with consequent beneficial effects on fish-fed diets, including enriched insect meal [
33].
Arthrospira platensis has gained notable attention for aquafeed formulation due to its protein content (55–70% dry weight), essential amino acids (methionine, cystine, and lysine), and pigments with antioxidant properties [
34]. These pigments can efficiently preserve the muscle quality of fish, counteracting the oxidation of important molecules such as polyunsaturated fatty acids (PUFA) [
35,
36,
37]. Spirulina has been directly used as an alternative ingredient to FM in diets for
M. rosenbergii, with promising results on growth performance, body composition, and digestive and antioxidant activities [
38,
39]. Conversely, to the best of our knowledge,
H. illucens prepupae meal (HM) has been scarcely tested in aquafeeds intended for this crustacean species [
40].
The present study represents the first attempt to replace 3% and 20% of marine derivatives (both FM and oil) with enriched HM (15%
w/
w of
A. platensis added to the growth substrate of
H. illucens larvae) in diets intended for
M. rosenbergii post-larvae. The 3% and 20% FM replacement levels were chosen, in accordance with a previous study [
33], because they represent a feed supplement inclusion or an ecological inclusion level (a global 20% FM substitution in aquafeeds has a proper ecological impact), respectively. Particular emphasis was given to (i) growth performances and morphological indices, (ii) physical and chemical features of both muscle and exoskeleton, and (iii) the overall health status determined by the histological and molecular analyses on the hepatopancreas, which is the largest organ in the digestive tract of Decapoda and which is involved in absorption and metabolization of nutrients, as well as in the immune response [
41]. To further improve prawn culture sustainability, a 60-day feeding trial was conducted in an aquaponic system, as different studies evidenced the suitability of this rearing method for
M. rosenbergii culture [
42,
43].
4. Discussion
The use of HM as an alternative and more sustainable ingredient in aquafeed formulation has been widely studied in fish [
63], but less in crustaceans. Several studies have been performed on the Pacific white shrimp (
Litopenaeus vannamei) using different dietary inclusion levels of
H. illucens larvae meal, obtaining promising results in terms of growth performances, antioxidant and immune response, digestive enzyme activity, intestinal microbiota, histomorphology of hepatopancreas, and muscle biochemical composition [
64,
65,
66]. However, to the best of our knowledge,
H. illucens has been scarcely explored as a possible alternative to FM in dietary formulations intended for a widely farmed Decapoda, such as the giant freshwater prawn [
40]. The present study represents the first attempt to evaluate the replacement of FM and FO with spirulina-enriched full-fat HM on
M. rosenbergii post-larvae reared in aquaponic systems to further improve aquaculture sustainability, promoting a zero-waste concept [
67]. The three experimental diets tested in the present study, using aquaponic systems, resulted in comparable survival and growth rates, confirming the suitability of the diets and the farming technique for
M. rosenbergii [
42,
68]. HM represents a good source of proteins and it was successfully used in the
M. rosenbergii diet, which is known to require a relatively high dietary protein content (37% on DM) to support growth performances [
69,
70]. Accordingly, growth performances and morphological features were not negatively affected by the HM dietary inclusions, which additionally led to a higher HEW in prawns from the HM20 group. These results are in line with previous studies that demonstrated that a replacement of FM with
H. illucens larvae meal up to 20% did not affect the growth performances of the Pacific white shrimp [
64,
65,
66,
71,
72]. However, it should be pointed out that the use of high-protein diets in crustacean aquaculture generally causes a consequently high nitrogenous waste production that can interfere with water quality [
73]. In this sense, as demonstrated by water quality analyses, aquaponic systems represented a valid method of exploiting this metabolic waste as a macronutrient for plant growth [
3,
42], avoiding economic losses due to prawn mortality linked to high ammonia levels and possible environmental issues [
74,
75].
The addition of spirulina in the insects’ rearing substrate resulted in an increase in α-tocopherol from the HM3 to HM20 diets and in the presence of carotenoids in the HM20 diet, respectively, highlighting that this enrichment procedure can be a valid method to transfer these important antioxidant molecules from spirulina to
H. illucens and, then, to the experimental diets. The provision of diets enriched with α-tocopherol and carotenoids has possibly reduced the secondary oxidative products in muscle samples from the HM0 group to the HM20 group, confirming the role of these molecules in preserving the muscle-quality traits [
35,
36,
37,
76].
However, the inclusion of full-fat HM in aquafeeds is usually associated with a dietary fatty acid profile alteration, especially regarding the SFA and the n3 PUFA contents [
32,
77]. Accordingly, the experimental diets tested here showed a slight increase in SFA content and a parallel wider decrease in n3 PUFA (especially in terms of EPA and DHA) from HM0 to HM20. Comparable amounts of n6 PUFA (largely constituted by LNA) were observed among the experimental diets. It has been demonstrated that the balance (more than the relative content) of dietary n3 and n6 PUFA is a crucial factor for the fatty acid requirement in prawns’ nutrition [
78]. In fact, despite the necessity to provide n3 PUFA [
79], freshwater prawns such as
M. rosenbergii might achieve the desired growth performance by relying on the dietary n6 PUFA [
69]. This is a consequence of the terrestrial origin of the giant freshwater prawn’s natural diet, rich in fatty acids from the n6 series [
80]. The absence of a preference for either n3 or n6 series fatty acids to fulfil the PUFA requirements [
81] sustained a proper growth of each experimental group. In addition, the tissue fatty acid profile of giant freshwater prawn generally reflects the dietary profile, due to the scarce ability of this crustacean to synthesize long-chain PUFA from shorter-chain precursors [
82,
83]. Accordingly, in the present study, the proportion among the fatty-acid classes in the diets reflected that of the muscle samples. The significantly higher amount of DHA in the muscle samples of prawns from the HM0 group emphasizes the importance of providing diets characterized by a proper amount of long-chain PUFA, in order to guarantee the quality of the final product and to meet the consumer’s requirements. However, while the experimental diets showed a higher DHA content compared to EPA, an opposite result was obtained in muscle samples, reflecting the ability of
M. rosenbergii to synthesize EPA from DHA and its bioconversion inability in the opposite direction [
83].
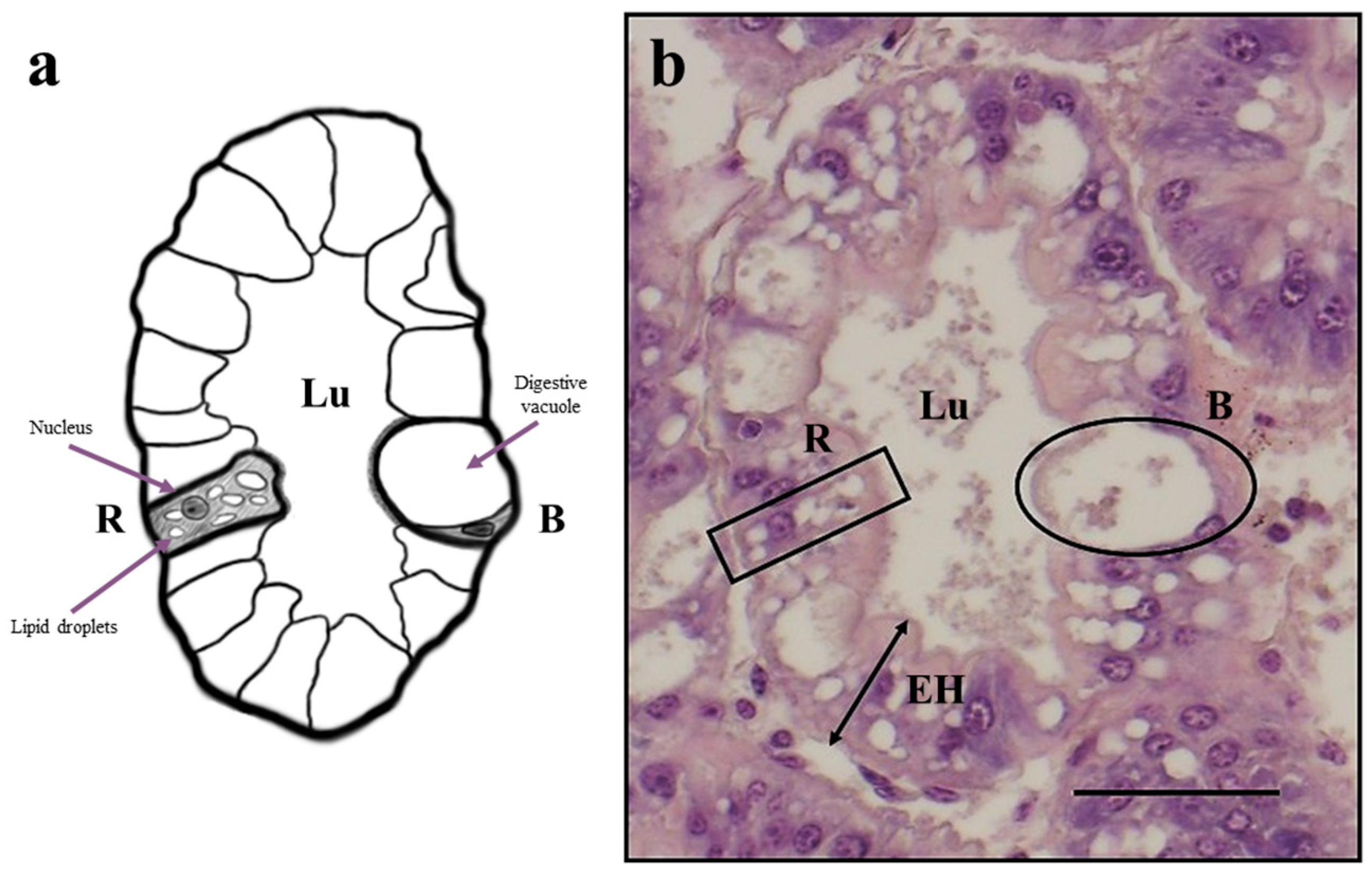
The dietary lipid content can also affect the lipid deposition in hepatopancreas; in particular, the R cells are known to be the main site for lipid storage and are thus considered to be a crucial indicator of crustaceans’ nutritional condition [
84,
85]. R cells are the most abundant cell type in the hepatopancreas, but nutritional stress can lead to a decrease in their prevalence in the tubules and vacuolation, with a consequent epithelium atrophy [
84,
86]. Accordingly, Wang et al. [
87] evidenced vacuolar degeneration coupled with an alteration of the epithelium structure in Pacific white shrimp when the replacement of FM with
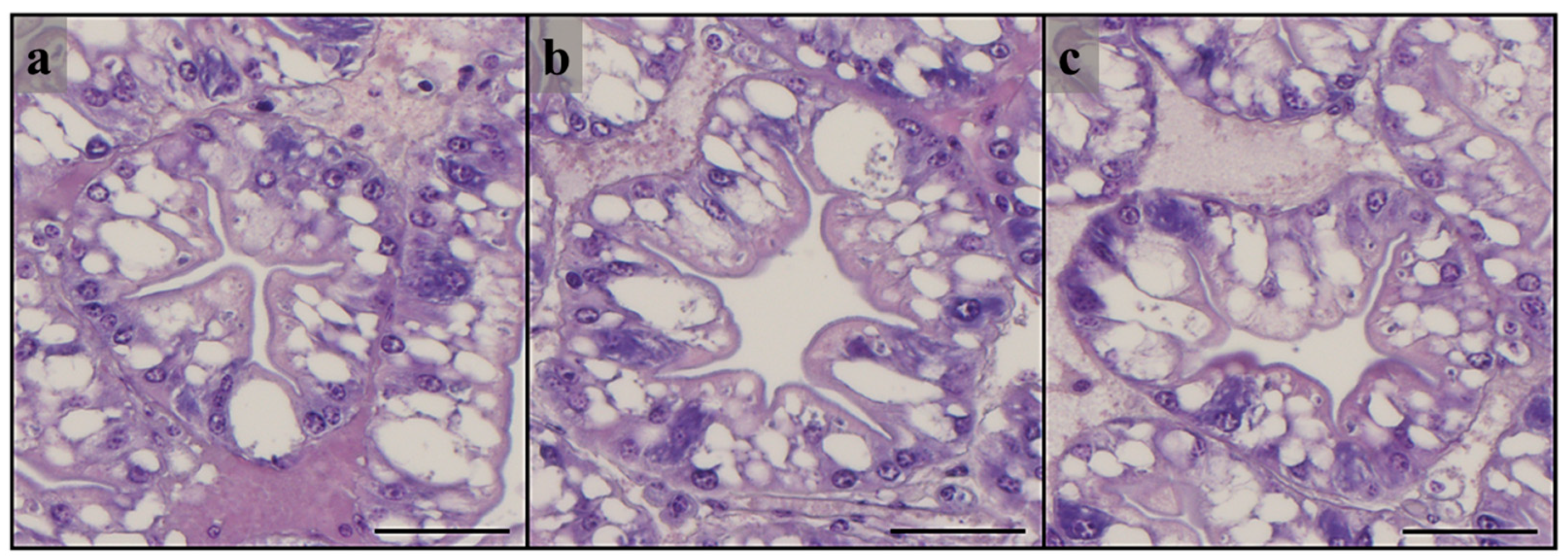
H. illucens larvae meal exceeded 60%. However, in the present study, which used lower replacement percentages, epithelium height and lipid storage were not affected by the provision of HM diets, as demonstrated by the comparable frequency of R cells and the high abundance of lipid droplets in all the experimental groups.
Dietary nutrients are absorbed and stored by the R cells to be subsequently mobilized to sustain energy-demanding processes such as molting [
88]. The massive presence of lipid droplets in R cells in all the experimental groups reflected a proper nutrient provision by the experimental diets and evidenced the necessity of prawns to store energy reserves during the inter-molt stage [
83]. Crustaceans periodically degrade the old exoskeleton in favor of a newly synthesized one (pre-molt phase) and, during molting, they face a rapid uptake of water that increases the animal size. After a post-molt phase, in which the hardness of the new exoskeleton is improved, the water absorbed during molting is gradually replaced by tissue growth and reserves are stored (inter-molt phase) [
83]. This process is under hormonal control. In particular, methyl farnesoate is the crustacean analogue of the pleiotropic juvenile hormone (JH) that is typical of insects and, like JH, it is involved in several processes including molting [
89]. This hormone, together with the ecdysone, gradually increases in the pre-molt phase, reaching the maximum levels near molting [
90,
91]. However, a key determinant for successful molting is the rapid decrease in methyl farnesoate due to its JHEH-mediated inactivation. It has been demonstrated that in
M. rosenbergii, the higher
jheh expression in the pre-molt stage led to a successful molt in a short time [
92]. The crustacean hepatopancreas is considered the main site for methyl farnesoate inactivation due to the highest level of
jheh expression [
92]. In the present study, the absence of significant differences in
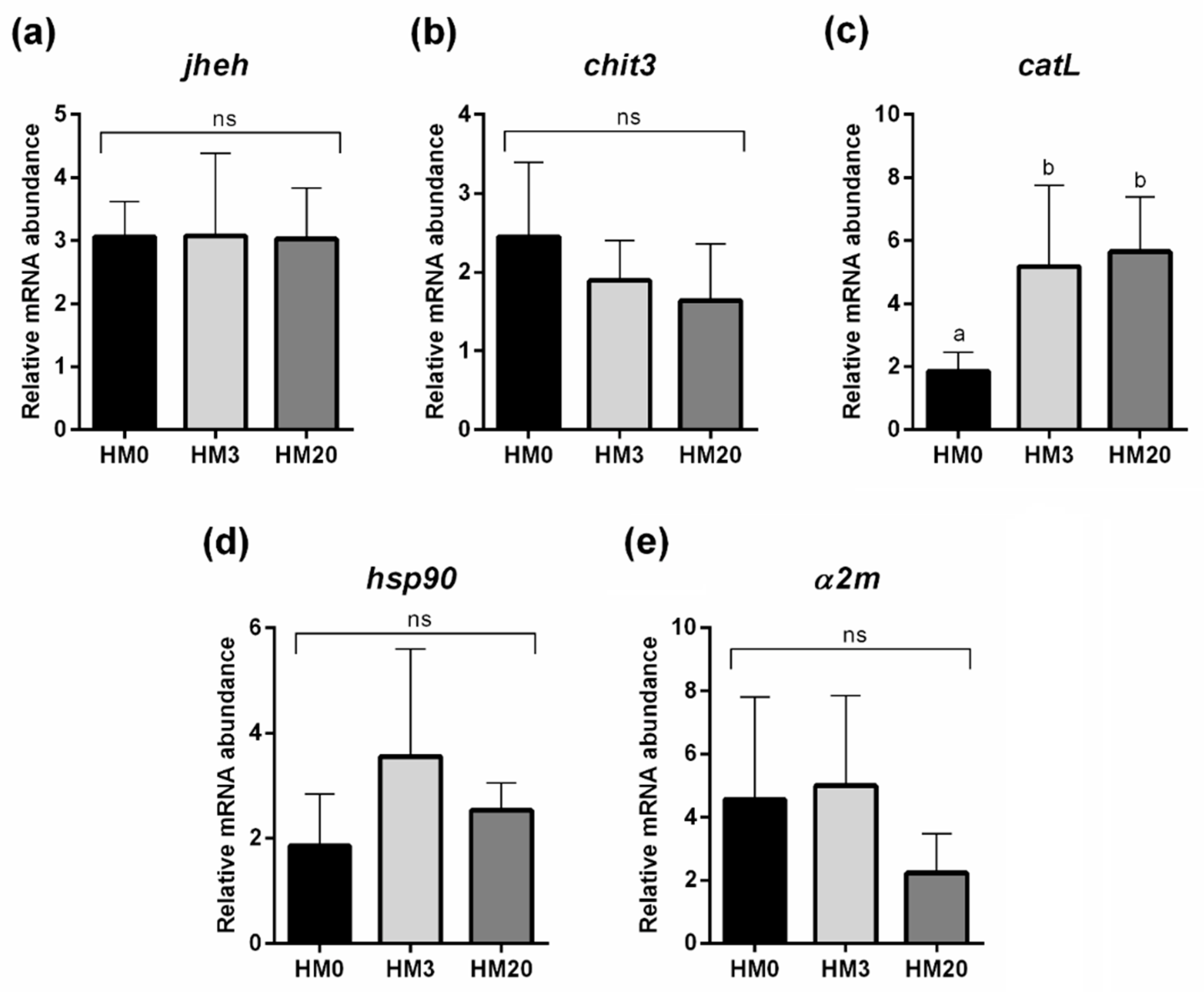
jheh gene expression represented further confirmation that the molting cycle (and thus somatic growth) and its regulation were not affected by the different dietary treatments.
The activity of digestive enzymes can have a direct effect in supporting molting and growth because of the breakage of the dietary high-weight biomolecules into more available forms required for metabolism or storage [
38]. In this regard, the primary role of the hepatopancreas is the digestive enzymes’ synthesis and secretion, the digestion of the ingested food, and the subsequent nutrients’ uptake [
93]. In particular, B cells produce digestive enzymes that are involved in intracellular digestion and concentrate in the vacuole the absorbed materials, and secrete them in the tubular lumen at the end of the digestive process for reabsorption by R cells [
88]. In the present study, a higher relative abundance of B cells was detected in the HM3 and HM20 groups than in the HM0 group. This result was fully supported by the expression of
catL which codifies for the cathepsin L, a protease stored in the digestive vacuole of B cells and secreted into the tubular lumen of the hepatopancreas for extracellular digestion [
94]. In this regard, it should be pointed out that Hu et al. [
94] detected
catL mRNA only in F cells but not in mature B cells that only host the active enzyme. However, several authors suggested that F cells are the precursors of B cells, and from an F cell to a B cell there is a continuum of transition (referred to as F/B cells) [
95]. Therefore, although we only measured the mRNA relative abundance of
catL, this result supports the abundance of B cells due to the fast synthesis of cathepsin L and the quick transition from F cell type to B cell type [
94].
An increased B cells population in the hepatopancreas is usually associated with the reduced bioavailability of dietary nutrients [
96,
97]. Several studies on fish reported that one of the possible drawbacks related to the dietary inclusion of HM can be attributed to chitin, which, due to its complex matrix, can impair nutrient absorption, creating a barrier on the absorptive epithelium [
98,
99,
100]. In addition, Kumar et al. [
101] found that
M. rosenbergii that were fed post-larvae with 10% purified chitin had a lower growth rate than those fed with 5% purified chitin, confirming that in crustaceans this complex molecule can act as a barrier against nutrient absorption. However, crustaceans possess specific chitinases expressed in the hepatopancreas, among which is the CHIT3 that is involved in the degradation of chitin-containing feeds; chitinases are part of the natural diet of this species [
83,
102]. In the present study, the
chit3 gene expression in the hepatopancreas did not significantly vary among the experimental groups, highlighting that HM diets did not enhance the production of this chitinase. For that reason, instead of a nutrient absorption impairment caused by chitin, the increased B cells number observed in the HM3 and HM20 groups could be related to the different compositions of the lipid fraction among the experimental diets. In fact, B cells may increase in number and size in response to high dietary lipids, producing and recycling fat emulsifiers to counteract their hydrophobic features that slow down the digestive processes and to facilitate their endocytosis [
103,
104]. As the diets were iso-lipidic, this conclusion could possibly be related to the dietary amount of SFA and MUFA that increased with the increasing dietary inclusion levels of HM and to the ability of
M. rosenbergii to digest and use these fatty-acid classes as an energy source [
80,
83].
Finally, the hepatopancreas, in addition to being a sensitive indicator for the nutritional status, is used to monitor the health conditions of crustaceans, as it is involved in the immune and stress response [
96,
105,
106]. Crustaceans only possess innate immunity due to the absence of cells from the myeloid lineage that are able to produce specific antibodies to face repetitive infections [
107]. Specifically, crustaceans have a rapid innate immunity consisting of various prophenoloxidase (proPO) system immune-related proteins, including α2-macroglobulin, which acts against pathogens and regulates the proPO activation [
108]. In addition, it has been demonstrated that the chitin-related genes expressed in the hepatopancreas of crustaceans may be involved in the immune response and disease resistance through direct or indirect regulation [
109,
110]. The expression of the immune response marker
a2m, supported by the absence of significant differences in terms of
chit3 gene expression, confirmed that the experimental diets used in the present study did not affect the health status of
M. rosenbergii. This conclusion is further supported by the expression of
hsp90, a stress-related biomarker that can also play a significant role in host immunity and the health of crustaceans [
111].


 ,
, 



{kind=link}
{kind=link}
{kind=link}