
Effects of Dietary Supplementation of Chinese Yam Polysaccharide on Carcass Composition, Meat Quality, and Antioxidant Capacity in Broilers

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Care
2.2. Experimental Diets and Animal Management
2.3. Slaughter Performance Measurement
2.4. Color Measurement
2.5. Shear Force Measurement
2.6. Serum Antioxidant Measurement
2.7. Quantitative Real-Time Polymerase Chain Reaction
2.8. Statistical Analysis
3. Result
3.1. Slaughter Performance
3.2. Color and Shear Force
3.3. Antioxidant Enzyme Activities
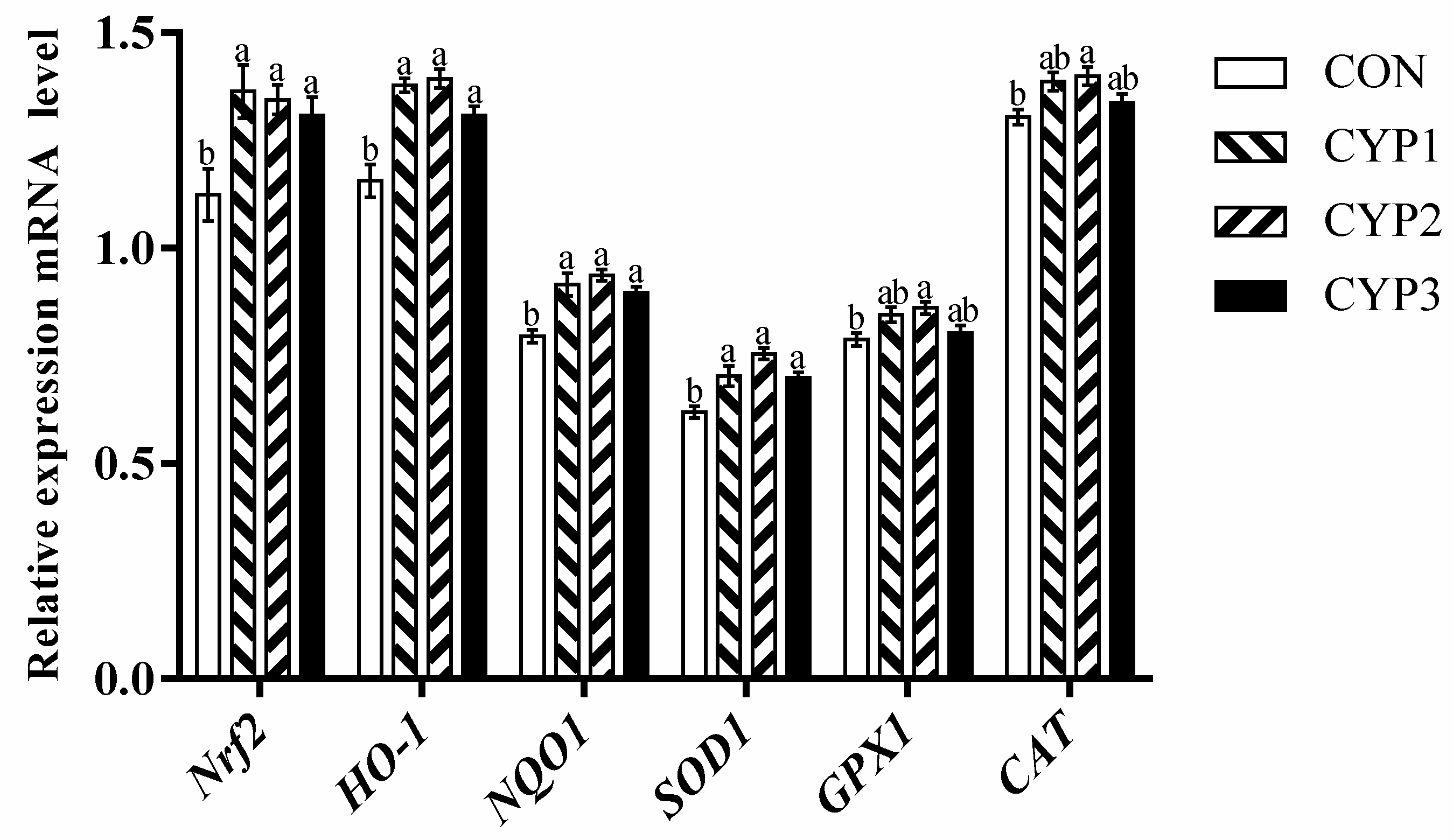
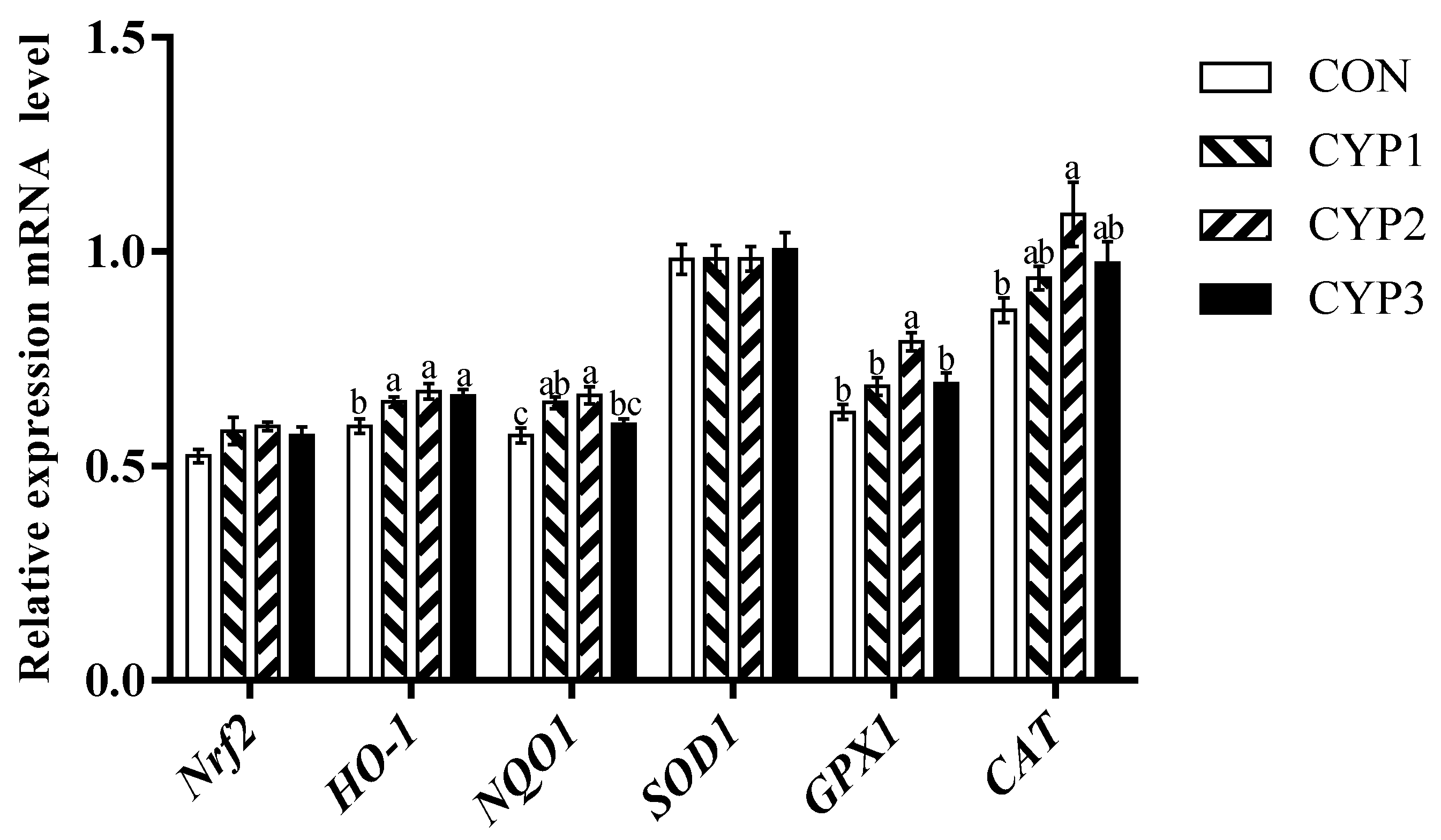
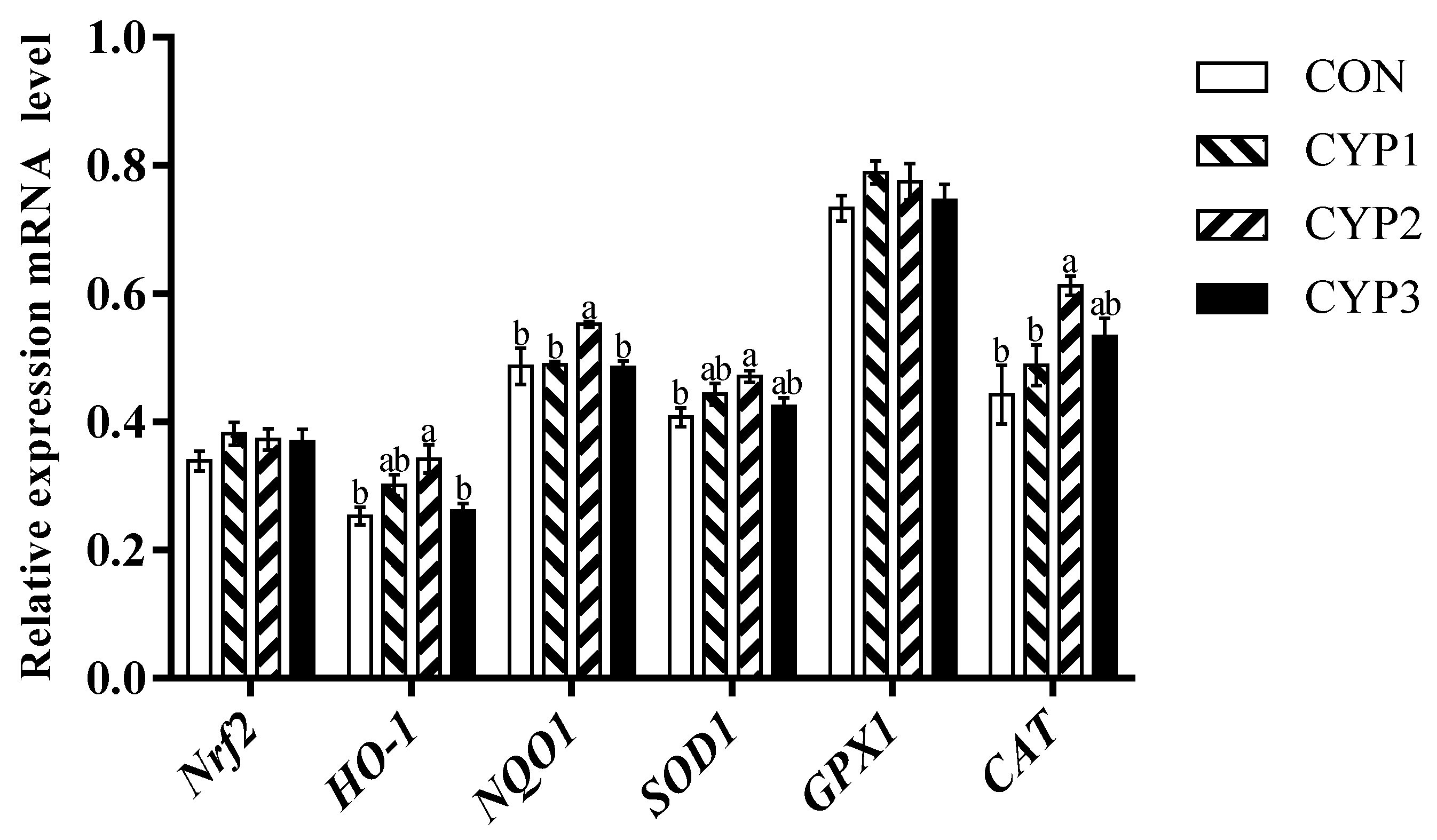
3.4. Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tsiouris, V.; Georgopoulou, I.; Batzios, C.; Pappaioannou, N.; Ducatelle, R.; Fortomaris, P. High stocking density as a predisposing factor for necrotic enteritis in broiler chicks. Avian Pathol. 2015, 44, 59–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.-C.; Pan, Z.-Y.; Zhao, Y.; Guo, Y.; Qiu, S.-J.; Balasubramanian, B.; Jha, R. Effects of Heat Stress on Production Performance, Redox Status, Intestinal Morphology and Barrier-Related Gene Expression, Cecal Microbiome, and Metabolome in Indigenous Broiler Chickens. Front. Physiol. 2022, 13, 890520. [Google Scholar] [CrossRef] [PubMed]
- Stein, K.; Farmer, J.; Singhal, S.; Marra, F.; Sutherland, S.; Quiñonez, C. The use and misuse of antibiotics in dentistry. J. Am. Dent. Assoc. 2018, 149, 869–884.e5. [Google Scholar] [CrossRef]
- Long, L.; Zhang, H.; Wang, F.; Yin, Y.; Yang, L.; Chen, J. Research Note: Effects of polysaccharide-enriched Acanthopanax senticosus extract on growth performance, immune function, antioxidation, and ileal microbial populations in broiler chickens. Poult. Sci. 2021, 100, 101028. [Google Scholar] [CrossRef]
- Li, W.; Hu, X.; Wang, S.; Jiao, Z.; Sun, T.; Liu, T.; Song, K. Characterization and anti-tumor bioactivity of astragalus polysaccharides by immunomodulation. Int. J. Biol. Macromol. 2020, 145, 985–997. [Google Scholar] [CrossRef]
- Wang, D.; Li, C.; Fan, W.; Yi, T.; Wei, A.; Ma, Y. Hypoglycemic and hypolipidemic effects of a polysaccharide from Fructus Corni in streptozotocin-induced diabetic rats. Int. J. Biol. Macromol. 2019, 133, 420–427. [Google Scholar] [CrossRef]
- Miao, Z.; Guo, L.; Liu, Y.; Zhao, W.; Zhang, J. Effects of dietary supplementation of chitosan on carcass composition and meat quality in growing Huoyan geese. Poult. Sci. 2020, 99, 3079–3085. [Google Scholar] [CrossRef]
- Undie, A.S.; Akubue, P. Pharmacological evaluation of Dioscorea dumetorum tuber used in traditional antidiabetic therapy. J. Ethnopharmacol. 1986, 15, 133–144. [Google Scholar] [CrossRef]
- Liu, W.; Yuan, Y.; Sun, C.; Balasubramanian, B.; Zhao, Z.; An, L. Effects of Dietary Betaine on Growth Performance, Digestive Function, Carcass Traits, and Meat Quality in Indigenous Yellow-Feathered Broilers under Long-Term Heat Stress. Animals 2019, 9, 506. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Li, D. Advances in research on the function and active ingredients of Chinese yam. Northwest Pharm. J. 2010, 25, 398–400. [Google Scholar]
- Shujun, W.; Hongyan, L.; Wenyuan, G.; Haixia, C.; Jiugao, Y.; Peigen, X. Characterization of new starches separated from different Chinese yam (Dioscorea opposita Thunb.) cultivars. Food Chem. 2006, 99, 30–37. [Google Scholar] [CrossRef]
- Huang, R.; Shen, M.; Yu, Y.; Liu, X.; Xie, J. Physicochemical characterization and immunomodulatory activity of sulfated Chinese yam polysaccharide. Int. J. Biol. Macromol. 2020, 165, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, W.; Gao, Q.; Zou, Y. Hypoglycemic Effect of Chinese Yam (Dioscorea opposita rhizoma) Polysaccharide in Different Structure and Molecular Weight. J. Food Sci. 2017, 82, 2487–2494. [Google Scholar] [CrossRef]
- Zhou, S.; Huang, G.; Chen, G. Extraction, structural analysis, derivatization and antioxidant activity of polysaccharide from Chinese yam. Food Chem. 2021, 361, 130089. [Google Scholar] [CrossRef]
- Liu, Y.; Li, H.; Fan, Y.; Man, S.; Liu, Z.; Gao, W.; Wang, T. Antioxidant and Antitumor Activities of the Extracts from Chinese Yam (Dioscorea opposite Thunb.) Flesh and Peel and the Effective Compounds. J. Food Sci. 2016, 81, H1553–H1564. [Google Scholar] [CrossRef]
- Ju, Y.; Xue, Y.; Huang, J.; Zhai, Q.; Wang, X.-H. Antioxidant Chinese yam polysaccharides and its pro-proliferative effect on endometrial epithelial cells. Int. J. Biol. Macromol. 2014, 66, 81–85. [Google Scholar] [CrossRef]
- National Research Council (NRC). Nutrient Requirements of Poultry, 9th ed.; The National Academy Press: Washington, DC, USA, 1994. [Google Scholar]
- Geldenhuys, G.; Hoffman, L.C.; Muller, N. The effect of season, sex, and portion on the carcass characteristics, pH, color, and proximate composition of Egyptian Goose (Alopochen aegyptiacus) meat. Poult. Sci. 2013, 92, 3283–3291. [Google Scholar] [CrossRef]
- Liu, H.W.; Zhou, D.W. Influence of pasture intake on meat quality, lipid oxidation, and fatty acid composition of geese. J. Anim. Sci. 2013, 91, 764–771. [Google Scholar] [CrossRef]
- Van Hoeck, V.; Sonawane, M.; Sanchez, A.L.G.; Van Dosselaer, I.; Buyens, C.; Morisset, D. Chromium propionate improves performance and carcass traits in broilers. Anim. Nutr. 2020, 6, 480–487. [Google Scholar] [CrossRef]
- Bai, H.; Bao, Q.; Zhang, Y.; Song, Q.; Liu, B.; Zhong, L.; Zhang, X.; Wang, Z.; Jiang, Y.; Xu, Q.; et al. Research Note: Effects of the rearing method and stocking density on carcass traits and proximate composition of meat in small-sized meat ducks. Poult. Sci. 2020, 99, 2011–2016. [Google Scholar] [CrossRef]
- Ölmez, M.; Şahin, T.; Karadağoğlu, Ö.; Yörük, M.A.; Kara, K.; Dalğa, S. Growth performance, carcass characteristics, and fatty acid composition of breast and thigh meat of broiler chickens fed gradually increasing levels of supplemental blueberry extract. Trop. Anim. Health Prod. 2021, 53, 109. [Google Scholar] [CrossRef] [PubMed]
- Hussein, M.M.A.; Abd El-Hack, M.E.; Mahgoub, S.A.; Saadeldin, I.M.; Swelum, A.A. Effects of clove (Syzygium aromaticum) oil on quail growth, carcass traits, blood components, meat quality, and intestinal microbiota. Poult. Sci. 2019, 98, 319–329. [Google Scholar] [CrossRef]
- Miao, Z.G.; Wei, P.P.; Yang, Z.G.; Wang, S.; Zhang, J.Z.; Liu, Y.L.; Chai, M.Y. Effect of expanded chitosan particle on carcass composition and meat quality characteristics of growing-finishing pigs. China Feed 2018, 15, 54–57. (In Chinese) [Google Scholar]
- Moore, V.; Young, O. The effects of electrical stimulation, thawing, ageing and packaging on the colour and display life of lamb chops. Meat Sci. 1991, 30, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Razafindrabe, R.-H.A.K.; Chen, K.; Zhao, X.; Yang, L.; Wang, L.; Chen, X.; Jin, S.; Geng, Z. Effects of different rearing systems on growth performance, carcass traits, meat quality and serum biochemical parameters of Chaohu ducks. Anim. Sci. J. 2018, 89, 672–678. [Google Scholar] [CrossRef]
- Chen, K.-L.; Chen, T.-T.; Lin, K.-J.; Chiou, P.W.-S. The Effects of Caponization Age on Muscle Characteristics in Male Chicken. Asian Australas. J. Anim. Sci. 2007, 20, 1684–1688. [Google Scholar] [CrossRef]
- Wang, W.; Wang, S.; Gong, Y.; Wang, J.; Tan, Z. Effects of vitamin A supplementation on growth performance, carcass characteristics and meat quality in Limosin×Luxi crossbreed steers fed a wheat straw-based diet. Meat Sci. 2007, 77, 450–458. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, M.; Gou, Z.; Jiang, S.; Zhang, Y.; Wang, M.; Tang, X.; Xu, B. The Effect of Camellia oleifera Cake Polysaccharides on Growth Performance, Carcass Traits, Meat Quality, Blood Profile, and Caecum Microorganisms in Yellow Broilers. Animals 2020, 10, 266. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, Y.; Wang, B.; Mei, X.; Jiang, S.; Li, W. Protocatechuic acid improved growth performance, meat quality, and intestinal health of Chinese yellow-feathered broilers. Poult. Sci. 2019, 98, 3138–3149. [Google Scholar] [CrossRef]
- Ding, X.; Yang, C.; Yang, Z.; Ren, X.; Wang, P. Effects of star anise (Illicium verum Hook.f) oil on the nuclear factor E2–related factor 2 signaling pathway of chickens during subclinical Escherichia coli challenge. Poult. Sci. 2020, 99, 3092–3101. [Google Scholar] [CrossRef]
- Abdel-Wahhab, M.A.; Aly, S.E. Antioxidant property of Nigella sativa (black cumin) and Syzygium aromaticum (clove) in rats during aflatoxicosis. J. Appl. Toxicol. 2005, 25, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Vararattanavech, A.; Ketterman, A.J. Multiple roles of glutathione binding-site residues of glutathione s-transferase. Protein Pept. Lett. 2003, 10, 441–448. [Google Scholar] [CrossRef]
- Zhu, Y.; Yang, L.; Zhang, C.; Tian, Y.; Zhang, F.; Li, X. Structural and functional analyses of three purified polysaccharides isolated from Chinese Huaishan-yams. Int. J. Biol. Macromol. 2018, 120, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-F.; Zhu, Q.; Wu, S. Preparation of oligosaccharides from Chinese yam and their antioxidant activity. Food Chem. 2015, 173, 1107–1110. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.A. Effects of Yam Polysaccharides on the Growth and Immune Performance of 1–21 Days. Master’s Thesis, Hunan Agricultural University, Changsha, China, 2014. (In Chinese). [Google Scholar]
- Rowe, L.J.; Maddock, K.R.; Lonergan, S.M.; Huff-Lonergan, E. Oxidative environments decrease tenderization of beef steaks through inactivation of μ-calpain1. J. Anim. Sci. 2004, 82, 3254–3266. [Google Scholar] [CrossRef]
- Estévez, M. Protein carbonyls in meat systems: A review. Meat Sci. 2011, 89, 259–279. [Google Scholar] [CrossRef]
- Shin, J.M.; Lee, K.-M.; Lee, H.J.; Yun, J.H.; Nho, C.W. Physalin A regulates the Nrf2 pathway through ERK and p38 for induction of detoxifying enzymes. BMC Complement. Altern. Med. 2019, 19, 101. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Wei, H.; Yu, H.; Xing, Q.; Zou, Y.; Zhou, Y.; Peng, J. Fish Skin Gelatin Hydrolysate Production by Ginger Powder Induces Glutathione Synthesis to Prevent Hydrogen Peroxide Induced Intestinal Oxidative Stress via the Pept1-p62-Nrf2 Cascade. J. Agric. Food Chem. 2018, 66, 11601–11611. [Google Scholar] [CrossRef]
- González-Burgos, E.; Carretero, M.E.; Gómez-Serranillos, M.P. Diterpenoids isolated from Sideritis species protect astrocytes against oxidative stress via Nrf2. J. Nat. Prod. 2012, 75, 1750–1758. [Google Scholar] [CrossRef]
- Liu, W.-C.; Zhu, Y.-R.; Zhao, Z.-H.; Jiang, P.; Yin, F.-Q. Effects of Dietary Supplementation of Algae-Derived Polysaccharides on Morphology, Tight Junctions, Antioxidant Capacity and Immune Response of Duodenum in Broilers under Heat Stress. Animals 2021, 11, 2279. [Google Scholar] [CrossRef]
- Long, L.; Kang, B.; Jiang, Q.; Chen, J. Effects of dietary Lycium barbarum polysaccharides on growth performance, digestive enzyme activities, antioxidant status, and immunity of broiler chickens. Poult. Sci. 2020, 99, 744–751. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Items | 1–28 d | 29–48 d |
|---|---|---|
| Ingredients (%) | ||
| Corn | 60.00 | 63.50 |
| Soybean meal | 32.00 | 29.00 |
| Wheat bran | 1.00 | - |
| Soybean oil | 1.00 | 2.00 |
| Fish meal | 2.00 | 1.60 |
| CaHPO4 | 1.30 | 1.30 |
| Limestone | 1.40 | 1.30 |
| NaCl | 0.30 | 0.30 |
| Premix 1 | 1.00 | 1.00 |
| Total | 100.00 | 100.00 |
| Nutrient levels (%) | ||
| Metabolic energy (MJ/kg) 2 | 12.13 | 12.55 |
| Crude protein | 21.00 | 20.00 |
| Calcium | 1.00 | 0.90 |
| Total p | 0.65 | 0.60 |
| Available p | 0.45 | 0.35 |
| Lysine | 0.50 | 0.38 |
| Methionine | 1.10 | 1.00 |
| Genes (Accession) 1 | Primer Sequence | Length (bp) |
|---|---|---|
| Nrf2 (MN416129) | F:5′-AACACACCAAAGAAAGACCCTCCTG -3′ R:5′- TTCACTGAACTGCTCCTTCGACATC-3′ | 207 |
| SOD1 (NM_205064) | F:5′- GGTCATCCACTTCCAGCAGCAG-3′ R:5′- AACGAGGTCCAGCATTTCCAGTTAG-3′ | 377 |
| CAT (NM_001031215) | F:5′- CTCTCAGAAGCCAGATGCCTTGAC-3′ R:5′- CAGCAACAGTGGAGAACCGTATAGC-3′ | 293 |
| GPX1 (NM_001277853) | F:5′-GAAGTGCGAGGTGAACGGGAAG-3′ R:5′-TGCAGTTTGATGGTCTCGAAGTGG-3′ | 228 |
| NQO1 (NM_001277621) | F:5′-AAGATTGAAGCGGCTGACCTGATC-3′ R:5′-AGGCGGCTCTTCCATGTACTCAG-3′ | 365 |
| HO-1 (HM237181) | F:5′-GAGTCTCCAACGCCACCAAGTTC-3′ R:5′-TCCTGCTTGTCCTCTCACTGTCC-3′ | 273 |
| β-actin (L08165) | F:5′-CATTGAACACGGTATTGTCACCAACTG-3′ R:5′-GTAACACCATCACCAGAGTCCATCAC-3′ | 270 |
| Item | CYP Level (mg/kg) 1 | SEM 2 | p-Value | |||
|---|---|---|---|---|---|---|
| CON | CYP1 | CYP2 | CYP3 | |||
| Live weight (kg) | 1.42 b | 1.60 a | 1.68 a | 1.58 a | 0.067 | 0.029 |
| Dressing percentage (%) | 89.96 | 91.02 | 92.20 | 92.62 | 0.838 | 0.087 |
| Half-eviscerated carcass percentage (%) | 79.70 b | 82.77 a | 82.87 a | 81.07 ab | 1.216 | 0.024 |
| Eviscerated carcass percentage (%) | 67.49 b | 69.56 ab | 70.40 a | 67.75 b | 1.315 | 0.024 |
| Breast muscle percentage (%) | 12.06 | 12.71 | 13.03 | 12.35 | 0.635 | 0.571 |
| Thigh muscle percentage (%) | 15.93 b | 17.89 a | 18.01 a | 16.50 b | 0.826 | 0.050 |
| Item 2 | CYP Level (mg/kg) 1 | SEM 3 | p-Value | |||
|---|---|---|---|---|---|---|
| CON | CYP1 | CYP2 | CYP3 | |||
| Breast muscle | ||||||
| L* | 52.50 | 52.08 | 52.08 | 52.20 | 0.426 | 0.738 |
| a* | 4.94 | 5.02 | 5.22 | 4.96 | 0.182 | 0.416 |
| b* | 11.29 a | 10.46 b | 10.46 b | 10.56 a | 0.288 | 0.039 |
| Shear force (N) | 16.31 a | 15.48 b | 14.71 b | 14.88 b | 0.270 | 0.002 |
| Thigh muscle | ||||||
| L* | 55.96 | 55.44 | 55.24 | 55.31 | 0.481 | 0.481 |
| a* | 10.22 | 10.04 | 9.71 | 10.22 | 0.201 | 0.084 |
| b* | 13.88 a | 12.57 ab | 11.90 b | 12.94 ab | 0.572 | 0.047 |
| Shear force (N) | 27.48 a | 21.94 c | 21.83 c | 23.18 b | 0.533 | 0.001 |
| Item 2 | CYP Level (mg/kg) 1 | SEM 3 | p-Value | |||
|---|---|---|---|---|---|---|
| CON | CYP1 | CYP2 | CYP3 | |||
| T-SOD (pg/mL) | 44.07 b | 43.96 b | 48.04 a | 47.49 a | 0.527 | 0.001 |
| T-AOC (U/mL) | 5.58 b | 5.89 a | 5.94 a | 5.88 a | 0.088 | 0.013 |
| GPX (pmol/mL) | 15.22 b | 16.50 a | 16.63 a | 16.57 a | 0.217 | 0.001 |
| GST (ng/L) | 459.74 d | 482.87 c | 530.34 a | 502.82 b | 4.693 | 0.001 |
| MDA (nmol/L) | 15.88 a | 15.40 b | 15.11 b | 15.27 b | 0.130 | 0.002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, Y.; Zhang, J.; Jin, Y.; Deng, J.; Shi, M.; Miao, Z. Effects of Dietary Supplementation of Chinese Yam Polysaccharide on Carcass Composition, Meat Quality, and Antioxidant Capacity in Broilers. Animals 2023, 13, 503. https://doi.org/10.3390/ani13030503
Chang Y, Zhang J, Jin Y, Deng J, Shi M, Miao Z. Effects of Dietary Supplementation of Chinese Yam Polysaccharide on Carcass Composition, Meat Quality, and Antioxidant Capacity in Broilers. Animals. 2023; 13(3):503. https://doi.org/10.3390/ani13030503
Chicago/Turabian StyleChang, Yadi, Jinzhou Zhang, Yan Jin, Jiahua Deng, Mingyan Shi, and Zhiguo Miao. 2023. "Effects of Dietary Supplementation of Chinese Yam Polysaccharide on Carcass Composition, Meat Quality, and Antioxidant Capacity in Broilers" Animals 13, no. 3: 503. https://doi.org/10.3390/ani13030503