
Genetic Analyses of Response of Local Ghanaian Tanzanian Chicken Ecotypes to a Natural Challenge with Velogenic Newcastle Disease Virus
 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Phenotypic Measurements
2.3. Bird Genotyping and Quality Control
2.4. Data Analyses
3. Results
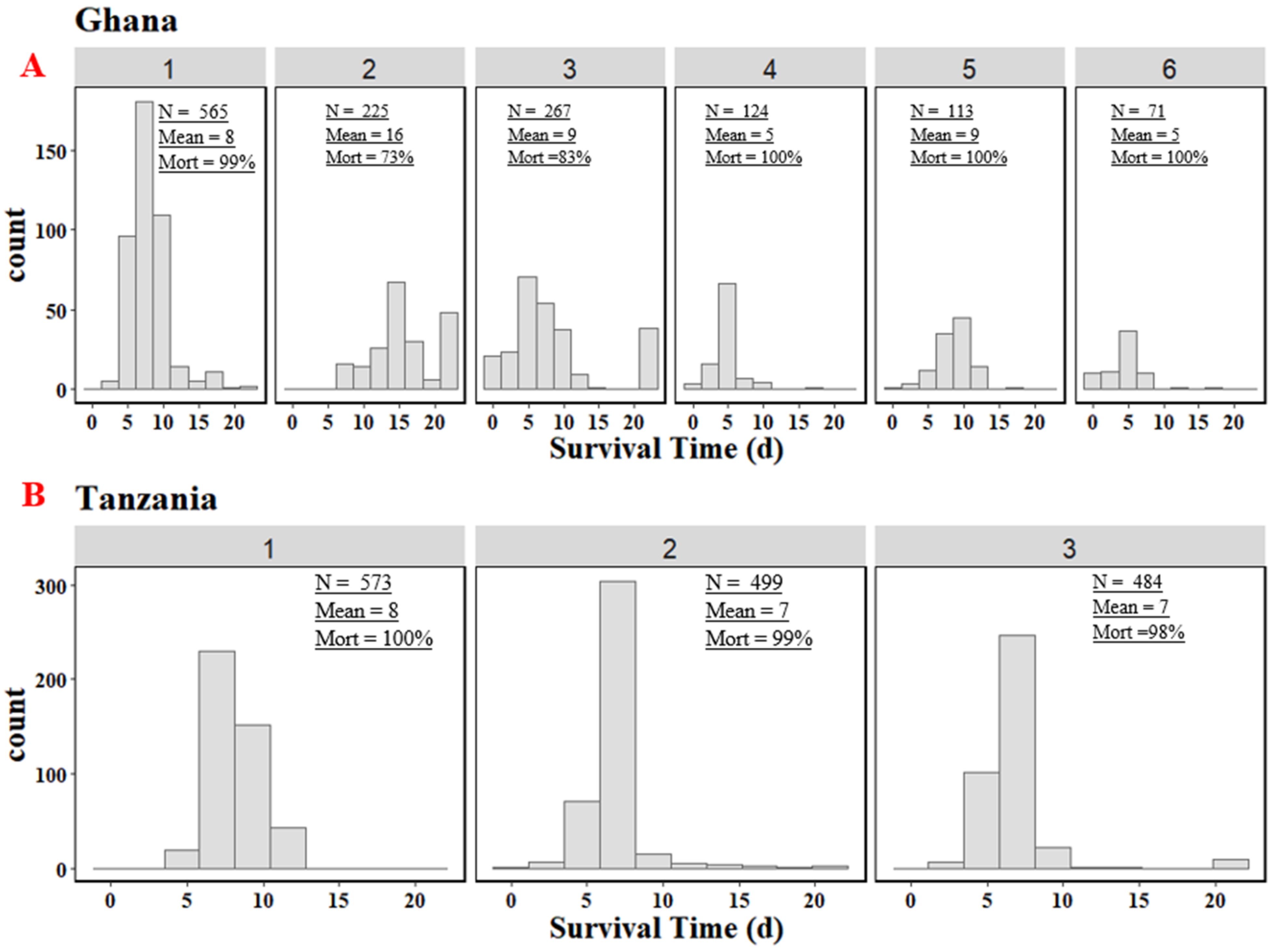
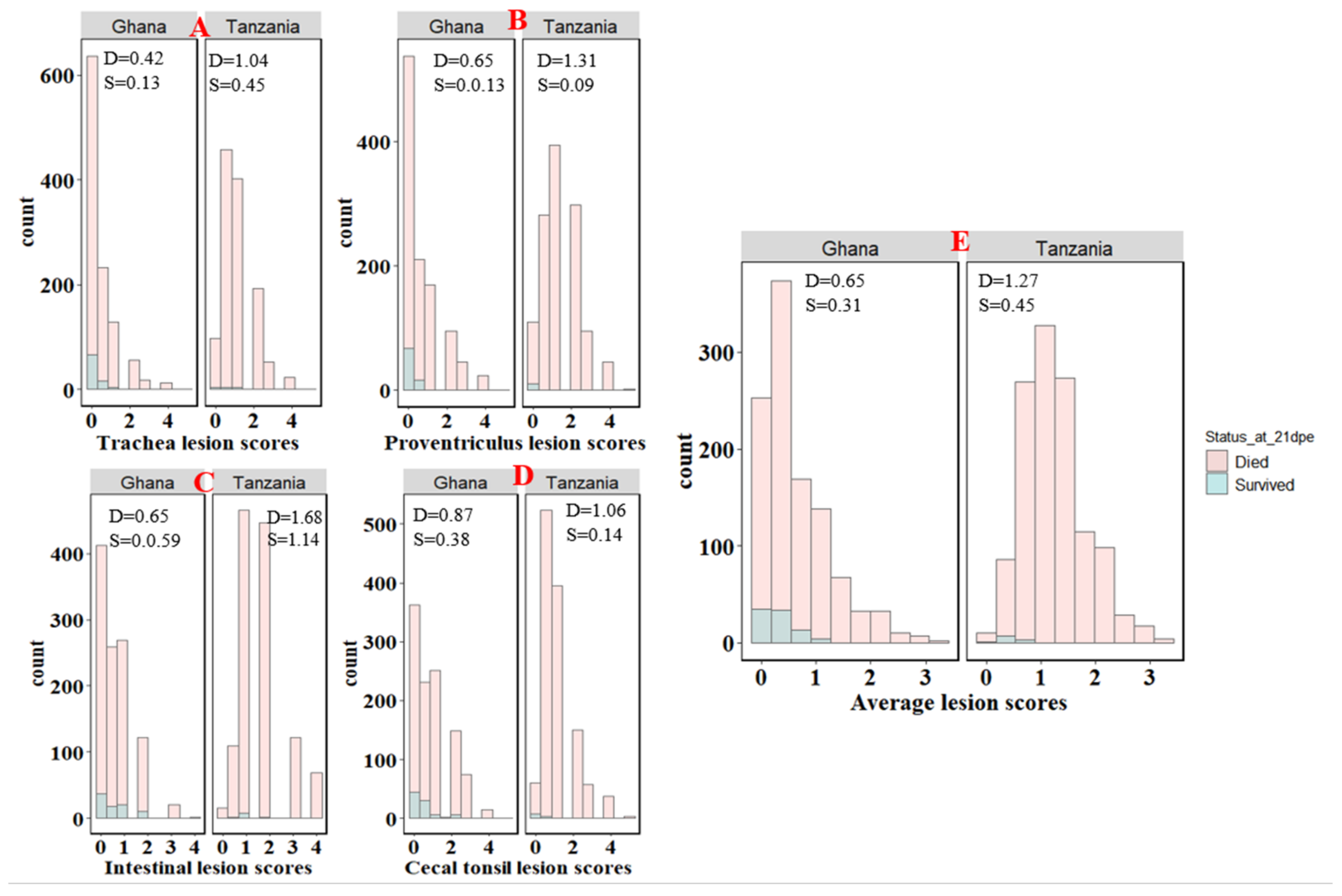
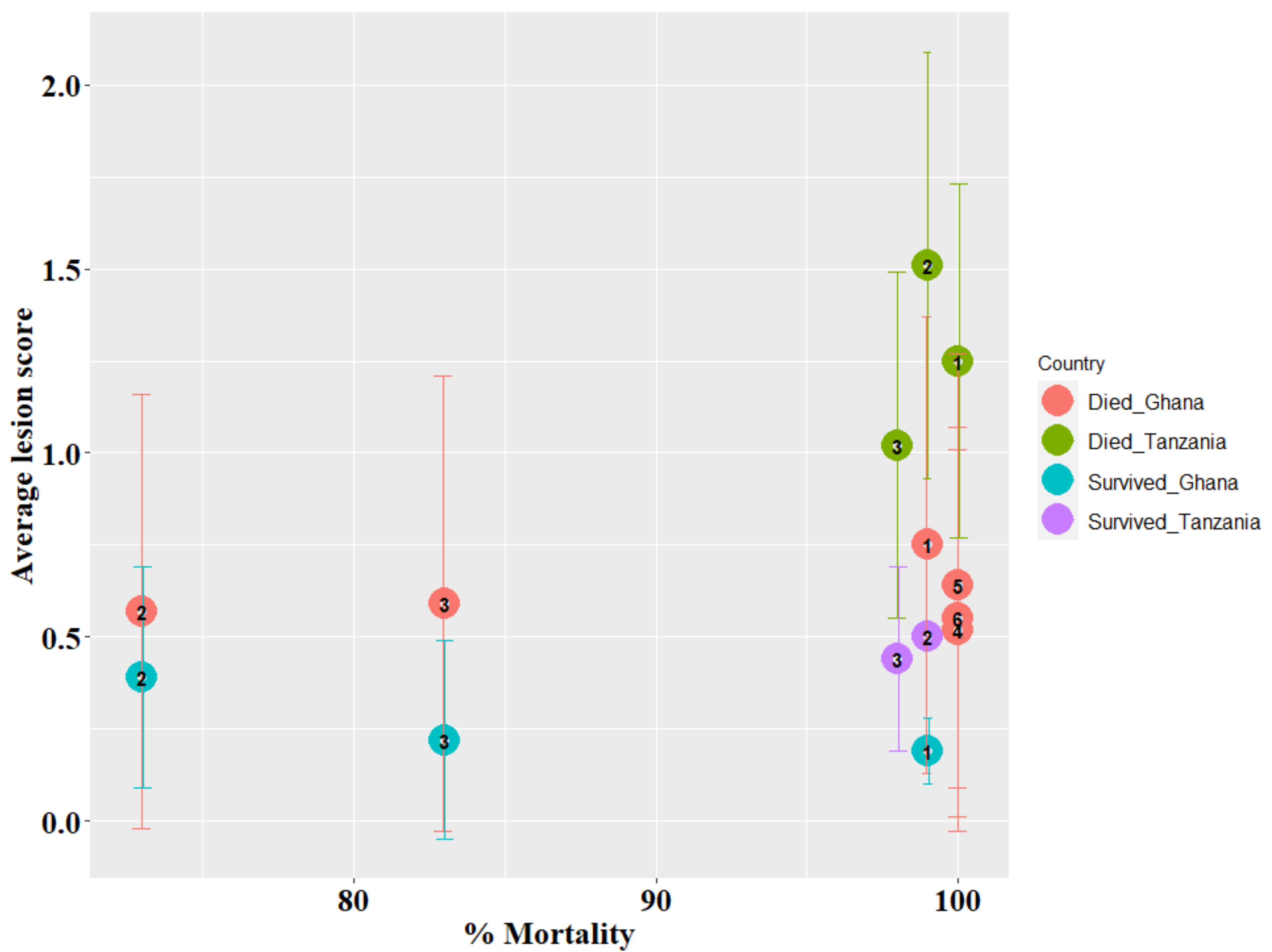
3.1. Survival Time and Lesion Scores
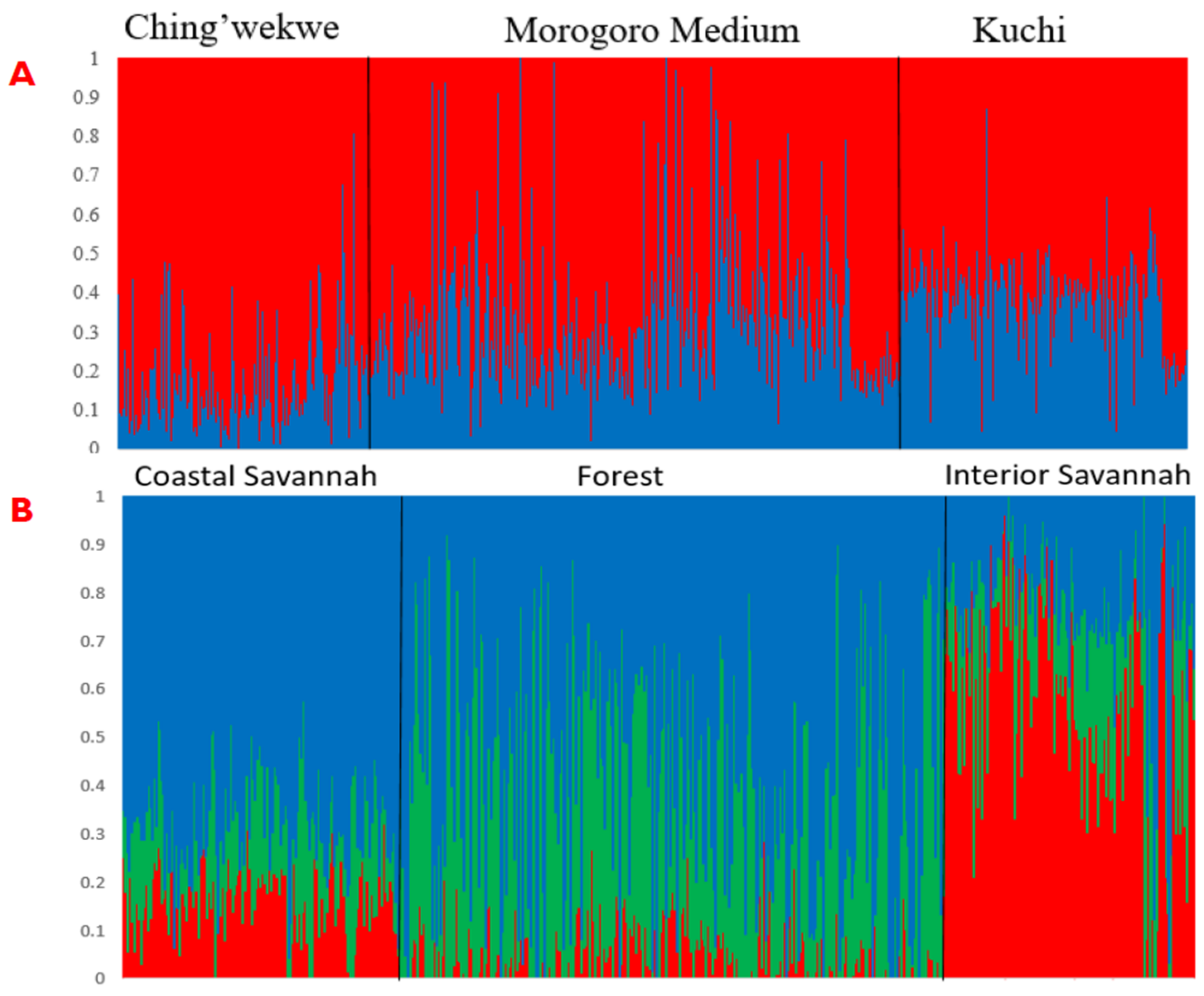
3.2. Subpopulation Proportions and Effects
3.3. Heritability Estimates
3.4. Correlations among Velogenic NDV Response Traits
3.5. Genetic Correlations between Velogenic and Lentogenic NDV Response Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guèye, E. The role of family poultry in poverty alleviation, food security and the promotion of gender equality in rural Africa. Outlook Agric. 2000, 29, 129–136. [Google Scholar] [CrossRef]
- Guèye, E.H.F. Village egg and fowl meat production in Africa. Worlds Poult. Sci. J. 1998, 54, 73–86. [Google Scholar] [CrossRef]
- Guèye, E. Family poultry research and development in low-income food-deficit countries: Approaches and prospects. Outlook Agric. 2002, 31, 13–21. [Google Scholar] [CrossRef]
- Kitalyi, A.J. Village Chicken Production Systems in Rural Africa: Household Food Security and Gender Issues. Food and Agriculture Organization of the United Nations: Rome, Italy, 1998. [Google Scholar]
- Mayo, M.A. Virus taxonomy—Houston 2002. Arch. Virol. 2002, 147, 1071–1076. [Google Scholar]
- Snoeck, C.J.; Ducatez, M.F.; Owoade, A.A.; Faleke, O.O.; Alkali, B.R.; Tahita, M.C.; Tarnagda, Z.; Ouedraogo, J.-B.; Maikano, I.; Mbah, P.O.; et al. Newcastle disease virus in West Africa: New virulent strains identified in non-commercial farms. Arch. Virol. 2009, 154, 47–54. [Google Scholar] [CrossRef]
- Adu, F.D.; Oyejide, O.; Ikede, B.O. Characterization of Nigerian Strains of Newcastle Disease Virus. Avian Diseases 1985, 29, 829–831. [Google Scholar] [CrossRef]
- Alexander, D.J. Newcastle disease. Br. Poult. Sci. 2001, 42, 5–22. [Google Scholar] [CrossRef]
- Alexander, D.J.; Senne, D.A. Newcastle disease, other avian paramyxoviruses, and Pneumovirus infections. Dis. Poult. 2008, 19, 75–115. [Google Scholar] [CrossRef]
- Miller, P.J.; Decanini, E.L.; Afonso, C.L. Newcastle disease: Evolution of genotypes and the related diagnostic challenges. Infect. Genet. Evol. 2010, 10, 26–35. [Google Scholar] [CrossRef]
- Hassan, M.K.; Afify, M.A.; Aly, M.M. Genetic Resistance of Egyptian Chickens to Infectious Bursal Disease and Newcastle Disease. Trop. Anim. Health Prod. 2004, 36, 1–9. [Google Scholar]
- Solomon, P.; Abolnik, C.; Joannis, T.M.; Bisschop, S. Virulent Newcastle disease virus in Nigeria: Identification of a new clade of sub-lineage 5f from livebird markets. Virus Genes 2012, 44, 98–103. [Google Scholar] [CrossRef]
- Da Silva, A.; Aston, E.; Chiwanga, G.; Birakos, A.; Muhairwa, A.; Kayang, B.; Kelly, T.; Zhou, H.; Gallardo, R. Molecular characterization of newcastle disease viruses isolated from chickens in Tanzania and Ghana. Viruses 2020, 12, 916. [Google Scholar] [CrossRef]
- Liu, X.F.; Wan, H.Q.; Ni, X.X.; Wu, Y.T.; Liu, W.B. Pathotypical and genotypical characterization of strains of Newcastle disease virus isolated from outbreaks in chicken and goose flocks in some regions of China during 1985–2001. Arch. Virol. 2003, 148, 1387–1403. [Google Scholar] [CrossRef]
- Cattoli, G.; Susta, L.; Terregino, C.; Brown, C. Newcastle disease: A review of field recognition and current methods of laboratory detection. J. Vet. Diagn. Investig. 2011, 23, 637–656. [Google Scholar]
- Copland, J.; Alders, R.G.; Southern, A.; Newcastle, A.; Project, C. The Australian village poultry development programme in Asia and Africa. Worlds Poult. Sci. J. 2018, 61, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Walugembe, M.; Mushi, J.R.; Amuzu-Aweh, E.N.; Chiwanga, G.H.; Msoffe, P.L.; Wang, Y.; Saelao, P.; Kelly, T.; Gallardo, R.A.; Zhou, H.; et al. Genetic analyses of tanzanian local chicken ecotypes challenged with newcastle disease virus. Genes 2019, 10, 546. [Google Scholar] [CrossRef] [Green Version]
- Walugembe, M.; Amuzu-Aweh, E.N.; Botchway, P.K.; Naazie, A.; Aning, G.; Wang, Y.; Saelao, P.; Kelly, T.; Gallardo, R.A.; Zhou, H.; et al. Genetic basis of response of ghanaian local chickens to infection with a lentogenic Newcastle disease virus. Front. Genet. 2020, 11, 739. [Google Scholar] [CrossRef]
- Igwe, A.O.; Ezema, W.S.; Eze, D.C.; Okoye, J.O.A. Lesions_2. Int. J. Poult. Sci. 2014, 13, 582–590. [Google Scholar]
- Onyema, I.; Eze, D.C.; Abba, Y.; Emennaa, P.E.; Shoyinka, S.V.O.; Okwor, E.C.; Ezema, W.S.; Ihedioha, J.I.; Okoye, J.O.A. Lesions of velogenic viscerotropic Newcastle disease virus infection were more severe in broilers than pullets. J. Appl. Anim. Res. 2019, 47, 189–194. [Google Scholar] [CrossRef]
- Fulton, J.E.; Arango, J.; Arthur, J.A.; Settar, P.; Kreager, K.S.; O’Sullivan, N.P. Improving the outcome of a Marek’s disease challenge in multiple lines of egg type chickens. Avian Dis. 2013, 57, 519–522. [Google Scholar] [CrossRef]
- Lewis, C.R.G.; Torremorell, M.; Galina-Pantoja, L.; Bishop, S.C. Genetic parameters for performance traits in commercial sows estimated before and after an outbreak of porcine reproductive and respiratory syndrome. J. Anim. Sci. 2009, 87, 876–884. [Google Scholar] [CrossRef]
- Serão, N.V.L.; Matika, O.; Kemp, R.A.; Harding, J.C.S.; Bishop, S.C.; Plastow, G.S.; Dekkers, J.C.M. Genetic analysis of reproductive traits and antibody response in a PRRS outbreak herd1. J. Anim. Sci. 2014, 92, 2905–2921. [Google Scholar] [CrossRef] [Green Version]
- Putz, A.M.; Harding, J.C.S.; Dyck, M.K.; Fortin, F.; Plastow, G.S.; Dekkers, J.C.M.; Canada, P. Novel resilience phenotypes using feed intake data from a natural disease challenge model in wean-to-finish pigs. Front. Genet. 2019, 9, 660. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Tibbs-Cortes, L.E.; Ashley, C.; Putz, A.M.; Lim, K.-S.; Dyck, M.K.; Fortin, F.; Plastow, G.S.; Dekkers, J.C.M.; Harding, J.C.S.; et al. The genetic basis of natural antibody titers of young healthy pigs and relationships with disease resilience. BMC Genom. 2020, 21, 648. [Google Scholar] [CrossRef]
- Mushi, J.R.; Amuzu-Aweh, E.N.; Chiwanga, G.C.; Mollel, E.L.; Walugembe, M.; Gallardo, R.A.; Kelly, T.R.; Lamont, S.J.; Zhou, H.; Msoffe, P.M.; et al. Phenotypic Differences and Relationships in Response to Infection with Lentogenic and Velogenic Strains of Newcastle Disease Virus in Local Tanzanian Chickens. 2022; Department of Veterinary Medicine and Public Health, Sokoine University: Morogoro, Tanzania, (Manuscript in preparation). [Google Scholar]
- Botchway, P.; Amuzu-Aweh, E.; Naazie, A.; Aning, G.; Otsyina, H.; Saelao, P.; Wang, Y.; Zhou, H.; Walugembe, M.; Dekkers, J.; et al. Host response to successive challenges with lentogenic and velogenic Newcastle disease virus in local chickens of Ghana. Poult. Sci. 2022, 101, 102138. [Google Scholar] [CrossRef]
- Mushi, J.R.; Chiwanga, G.H.; Amuzu-Aweh, E.N.; Walugembe, M.; Max, R.A.; Lamont, S.J.; Kelly, T.R.; Mollel, E.L.; Msoffe, P.L.; Dekkers, J.; et al. Phenotypic variability and population structure analysis of Tanzanian free-range local chickens. BMC Veter- Res. 2020, 16, 360. [Google Scholar] [CrossRef]
- Spackman, E.; Senne, D.A.; Myers, T.J.; Bulaga, L.L.; Garber, L.P.; Perdue, M.L.; Lohman, K.; Daum, L.T.; Suarez, D.L. Development of a real-time reverse transcriptase PCR assay for type a influenza virus and the avian H5 and H7 hemagglutinin subtypes. J. Clin. Microbiol. 2002, 40, 3256–3260. [Google Scholar] [CrossRef] [Green Version]
- Wise, M.G.; Suarez, D.L.; Seal, B.S.; Pedersen, J.C.; Senne, D.A.; King, D.J.; Kapczynski, D.R.; Spackman, E. Development of a real-time reverse-transcription PCR for detection of Newcastle disease virus RNA in clinical samples. J. Clin. Microbiol. 2004, 42, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Rowland, K.; Wolc, A.; Gallardo, R.A.; Kelly, T.; Zhou, H.; Dekkers, J.C.M.; Lamont, S.J. Genetic analysis of a commercial egg laying line challenged with Newcastle disease virus. Front. Genet. 2018, 9, 326. [Google Scholar] [CrossRef]
- Walugembe, M.; Amuzu-Aweh, E.N.; Lim, K.-S.; Wang, Y.; Chouicha, N.; Kelly, T.; Muhairwa, A.P.; Kayang, B.B.; Msoffe, P.L.M.; Naazie, A.; et al. Development of a Low-Density SNP Panel for Local Ghanaian and Tanzanian Chicken Ecotypes. Available online: https://www.wageningenacademic.com/pb-assets/wagen/WCGALP2022/55_013.pdf (accessed on 15 July 2022).
- Sargolzaei, M.; Chesnais, J.P.; Schenkel, F.S. A new approach for efficient genotype imputation using information from relatives. BMC Genom. 2014, 15, 478. [Google Scholar] [CrossRef] [Green Version]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef] [Green Version]
- Gilmour, A.R.; Gogel, B.J.; Cullis, B.R.; Welham, S.J.; Thompson, R. ASReml User Guide Release 4.1 Structural Specification; VSN International Ltd.: Hemel Hempstead, UK, 2015; pp. 1–30. [Google Scholar] [CrossRef]
- VanRaden, P. Efficient methods to compute genomic predictions. J. Dairy Sci. 2008, 91, 4414–4423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimitrov, K.M.; Afonso, C.L.; Yu, Q.; Miller, P.J. Newcastle disease vaccines—A solved problem or a continuous challenge? Vet. Microbiol. 2017, 206, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Msoffe, P.L.M.; Chiwanga, G.H.; Cardona, C.J.; Miller, P.J.; Suarez, D.L. Isolation and characterization of newcastle disease virus from live bird markets in Tanzania. Avian Dis. 2019, 63, 634–640. [Google Scholar] [CrossRef]
- Hussein, E.A.; Hair-Bejo, M.; Adamu, L.; Omar, A.R.; Arshad, S.S.; Awad, E.A.; Aini, I. Scoring system for lesions induced by different strains of Newcastle disease virus in chicken. Veter. Med. Int. 2018, 2018, 9296520. [Google Scholar] [CrossRef]
- Ezema, W.S.; Eze, D.C.; Shoyinka, S.V.O.; Okoye, J.O.A. Atrophy of the lymphoid organs and suppression of antibody response caused by velogenic Newcastle disease virus infection in chickens. Trop. Anim. Health Prod. 2016, 48, 1703–1709. [Google Scholar] [CrossRef] [PubMed]
- Touko, B.A.H.; Mbiydzenyuy, A.T.K.; Tumasang, T.T.; Awah-Ndukum, J. Heritability estimate for antibody response to vaccination and survival to a newcastle disease infection of native chicken in a low-input production system. Front. Genet. 2021, 12, 666947. [Google Scholar] [CrossRef]
- Berghof, T.V.L.; Van Der Klein, S.A.S.; Arts, J.A.J.; Parmentier, H.K.; Van Der Poel, J.J.; Bovenhuis, H. Genetic and non-genetic inheritance of natural antibodies binding keyhole limpet hemocyanin in a purebred layer chicken line. PLoS ONE 2015, 10, e0131088. [Google Scholar] [CrossRef]
- Berghof, T.V.L.; Visker, M.H.P.W.; Arts, J.A.J.; Parmentier, H.K.; Van Der Poel, J.J.; Vereijken, A.L.J.; Bovenhuis, H. Genomic region containing toll-like receptor genes has a major impact on total IgM antibodies including KLH-Binding IgM natural antibodies in chickens. Front. Immunol. 2018, 8, 1879. [Google Scholar] [CrossRef]
- Fulton, J.E.; Juul-Madsen, H.R.; Ashwell, C.M.; McCarron, A.M.; Arthur, J.A.; O, N.P.; Taylor, R.L., Jr. Molecular genotype identification of the gallus Gallus major histocompatibility complex. Immunogenetics 2006, 58, 407–421. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ghana Subpopulations | Tanzania | ||
|---|---|---|---|
| Trait | Red | Green | Red |
| Pre-exposure growth rate (g/d) | 0.61 (0.29) | 0.35 (0.26) | 1.22 (0.53) |
| Pre-exposure growth rate (g/d) | 0.96 (0.62) | 1.15 (0.59) | −2.02 (1.77) |
| Trachea lesion score | 0.16 (0.09) | −0.04 (0.09) | 0.39 (0.11) |
| Proventriculus lesion score | 0.08 (0.19) | −0.15 (0.17) | 0.33 (0.34) |
| Intestinal lesion score | 0.08 (0.10) | 0.05 (0.10) | 0.21 (0.21) |
| Cecal tonsil lesion score | 0.17 (0.17) | −0.07 (0.16) | 0.22 (0.18) |
| Average lesion score | 0.09 (0.10) | −0.05 (0.10) | 0.28 (0.144) |
| Natural antibody 1 | −0.06 (0.07) | 0.04 (0.07) | −0.09 (0.14) |
| Anti-NDV antibody 7 dpe 1 | 0.03 (0.18) | 0.24 (0.17) | −0.24 (0.18) |
| Survival Time (days) | 0.82 (0.99) | 2.43 (0.90) | −0.51 (0.55) |
| Tear viral load 2 dpe 1 | −0.06 (0.15) | −0.06 (0.15) | 0.03 (0.08) |
| Tear viral load 6 dpe 1 | 0.48 (0.51) | −0.05 (0.48) | 0.10 (0.24) |
| Cloacal viral load 2 dpe 1 | 0.06 (0.23) | −0.01 (0.21) | 0.18 (0.13) |
| Cloacal viral load 4 dpe 1 | 0.02 (0.23) | 0.25 (0.21) | 0.12 (0.18) |
| Tanzania | Ghana | Combined | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N 1 | Mean (SD) | h 2 (se) | Dam (se) | N 1 | Mean (SD) | h 2 (se) | Dam (se) | N 1 | Mean (SD) | h 2 (se) | Dam (se) | |
| Pre-exposure growth (g/d) | 1239 | 3.82 (1.56) | 0.32 (0.06) | 0.02 (0.00) | 1344 | 4.34 (1.68) | 0.37 (0.06) | 0.00 (0.01) | 2572 | 4.16 (1.61) | 0.35 (0.04) | 0.01 (0.01) |
| Post-exposure growth (g/d) | 1002 | 2.93 (5.33) | 0.01 (0.05) | 0.03 (0.03) | 816 | 0.14 (3.93) | 0.01 (0.07) | 0.04 (0.03) | 1818 | 1.69 (8.42) | 0.04 (0.04) | 0.03 (0.02) |
| Trachea lesions | 1239 | 1.03 (0.83) | 0.00 (0.00) | 0.04 (0.01) | 1170 | 0.43 (0.77) | 0.00 (0.04) | - | 2409 | 0.72 (0.83) | 0.01 (0.03) | 0.01 (0.01) |
| Proventriculus lesions | 1239 | 1.29 (0.98) | 0.23 (0.07) | 0.04 (0.02) | 1170 | 0.61 (0.90) | 0.14 (0.06) | 0.01 (0.02) | 2409 | 0.96 (1.01) | 0.19 (0.05) | 0.02 (0.02) |
| Intestinal lesions | 1239 | 1.66 (0.91) | 0.09 (0.04) | - | 1170 | 0.66 (0.77) | 0.03 (0.04) | - | 2409 | 1.18 (0.97) | 0.08 (0.03) | - |
| Cecal Tonsil lesions | 1239 | 1.04 (0.87) | 0.05 (0.04) | - | 1170 | 0.84 (0.94) | 0.09 (0.05) | - | 2409 | 0.95 (0.92) | 0.08 (0.03) | - |
| Average lesion score | 1239 | 1.26 (0.55) | 0.14 (0.06) | 0.02 | 1170 | 0.64 (0.62) | 0.08 (0.05) | - | 2409 | 0.95 (0.66) | 0.10 (0.03) | 0.01 (0.03) |
| Natural antibody 3 | 430 | 7.11 (1.92) | 0.18 (0.14) | 0.05 | 936 | 3.70 (0.45) | 0.11 (0.10) | 0.01 (0.02) | 1960 | 4.35 (0.79) | 0.13 (0.06) | 0.04 (0.02) |
| Anti-NDV antibody 7 dpe 3 | 298 | 1.98 (0.65) | 0.00 (0.00) | 0.00 | 576 | 2.03 (0.72) | 0.21 (0.10) | - | 874 | 2.85 (1.87) | 0.12 (0.08) | - |
| Survival_time (days) | 1238 | 7.12 (2.10) | 0.23 (0.06) | 0.00 | 1160 | 9.11 (5.19) | 0.27 (0.06) | 0.00 | 2398 | 8.00 (3.97) | 0.21 (0.04) | - |
| Tears viral load 2 dpe 3 | 660 | 0.11 (0.39) | - | - | 546 | 1.36 (1.06) | 0.00 (0.00) | - | 617 | 1.23 (1.07) | 0.00 (0.00) | - |
| Tears viral load 6 dpe 3 | 266 | 1.15 (1.42) | - | 0.9 (0.06) | 187 | 3.59 (1.47) | 0.44 (0.29) | - | 454 | 3.16 (1.56) | 0.28 (0.15) | - |
| Cloacal viral load 2 dpe 3 | 280 | 0.44 (0.57) | 0.06 (0.19) | - | 546 | 1.78 (0.33) | 0.00 (0.00) | 0.05 (0.04) | 826 | 0.28 (0.51) | 0.16 (0.11) | - |
| Cloacal viral load 4 dpe 3 | 297 | 0.56 (0.69) | 0.00 (0.00) | - | 428 | 3.19 (0.81) | 0.38 (0.14) | 0.04 (0.04) | 725 | 0.59 (0.74) | 0.21 (0.11) | 0.03 (0.04) |
| Cloacal viral load 6 dpe 3 | 143 | - | - | - | 310 | 4.51 (1.17) | 0.17 (0.17) | 0.12 (0.07) | 453 | 1.05 (1.07) | 0.15 (0.15) | 0.06 (0.06) |
| Growth Rate | Lesion Scores | Antibody | Survival Time | Viral Load | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pre- Exposure | Post-Exposure | Proventri-culus | Intestine | Cecal Tonsil | Average | Natural2 | 7 dpe2 | Cloacal 2 dpe2 | Cloacal 4 dpe2 | Tears 6 dpe2 | ||
| Growth rate pre-exposure 1 | - −0.08 (0.27) −0.22 (0.31) | 0.09 (0.11) 0.14 (0.15) 0.01 (0.19) | 0.56 (0.16) 0.55 (0.21) 0.56 (0.39) | 0.39 (0.17) 0.60 (0.34) 0.25 (0.22) | 0.40 (0.14) 0.47 (0.17) 0.23 (0.22) | 0.25 (0.16) 0.09 (0.25) −0.18 (0.05) | 0.77 (0.88) - −0.10 (0.23) | 0.12(0.11) −0.14 (0.19) 0.21 (0.15) | −0.27 (0.21) - - | −0.27 (0.21) - - | - - - | |
| Growth rate post-exposure 1 | 0.01 (0.04) −0.04 (0.03) −0.11 (0.03) | 0.23 (0.22) - - | −0.26 (0.29) - - | −0.25 (0.29) - - | 0.07 (0.26) - - | 0.10 (0.25) - - | 0.07 (0.40) - - | 0.48 (0.65) - - | 0.81 (0.54) - - | 0.01 (0.36) - - | −0.28 (0.42) - - | |
| Proventri-culus lesion score | 0.06 (0.02) 0.05 (0.03) 0.08 (0.03) | 0.04 (0.02) 0.06 (0.03) 0.10 (0.04) | - −0.43 (0.26) 0.95 (0.44) | 0.43 (0.17) 0.21 (0.03) 0.74 (0.27) | 0.86 (0.06) 0.84 (0.09) 0.88 (0.15) | 0.23 (0.16) 0.11 (0.39) 0.11 (0.45) | −0.66 (0.21) - −0.93 (0.24) | −0.23 (0.13) −0.17 (0.22) −0.33 (0.31) | −0.14 (0.25) - - | 0.05 (0.22) - 0.29 (0.30) | −0.27 (0.25) - - | |
| Intestine lesion score | 0.11 (0.02) 0.11 (0.03) 0.09 (0.03) | −0.01 (0.02) −0.01 (0.03) 0.06 (0.04) | 0.11 (0.02) 0.15 (0.03) 0.46 (0.02) | 0.91 (0.23) 0.23 (0.03) - | 0.81 (0.13) 0.83 (0.18) - | 0.03 (0.24) −0.33 (0.42) - | −0.28 (0.37) - - | −0.25 (0.20) −0.31 (0.36) - | −0.20 (0.39) - - | 0.44 (0.32) - - | −0.48 (0.37) - - | |
| Cecal tonsil lesion score | 0.06 (0.02) 0.04 (0.34) 0.10 (0.34) | −0.02 (0.02) −0.01 (0.02) 0.04 (0.04) | 0.40 (0.02) 0.21 (0.04) 0.57 (0.02) | 0.37 (0.02) 0.23 (0.03) 0.57 (0.02) | 0.80 (0.10) −0.70 (0.38) 0.85 (0.14) | 0.07 (0.23) −0.01 (0.05) −0.53 (0.60) | −0.53 (0.34) - −0.68 (0.30) | −0.38 (0.20) −0.10 (0.03) −0.14 (0.35) | −0.48 (0.36) - - | 0.40 (0.31) - 0.32 (0.32) | 0.81 (0.49) - - | |
| Average Lesion score | 0.13 (0.02) 0.13 (0.03) 0.13 (0.03) | 0.01 (0.02) 0.03 (0.03) 0.06 (0.04) | 0.72 (0.01) 0.65 (0.02) 0.78 (0.01) | 0.65 (0.01) 0.61 (0.02) 0.74 (0.01) | 0.75 (0.01) 0.62 (0.02) 0.85 (0.01) | 0.22 (0.21) 0.26 (0.43) 0.81 (0.58) | −0.56 (0.28) - −0.68 (0.30) | −0.40 (0.17) - - | −0.37 (0.30) - - | 0.42 (0.27) - 0.62 (0.35) | 0.07 (0.32) - - | |
| Natural Antibody2 | 0.11 (0.03) 0.29 (0.06) −0.07 (0.06) | 0.05 (0.03) 0.10 (0.06) 0.07 (0.05) | 0.02 (0.02) −0.05 (0.07) 0.00 (0.04) | −0.01 (0.02) −0.05 (0.05) 0.00 (0.04) | −0.03 (0.02) 0.13 (0.06) −0.07 (0.05) | −0.01 (0.02) −0.05 (0.06) −0.06 (0.05) | 0.34 (0.30) - 0.90 (0.87) | −0.04 (0.18) −0.05 (0.40) −0.09 (0.34) | 0.43 (0.36) - - | 0.14 (0.24) - - | −0.01 (0.32) 0.13 (0.65) - | |
| Antibody 7 dpe2 | 0.05 (0.03) - −0.07 (0.06) | −0.09 (0.04) - −0.20 (0.05) | −0.12 (0.04) - −0.17 (0.05) | −0.04 (0.04) - −0.05 (0.05) | −0.07 (0.04) - 0.04 (0.09) | −0.09 (0.04) - −0.11 (0.05) | 0.09 (0.04) - - | 0.48 (0.88) - 0.04 (0.27) | 0.46 (0.42) - - | 0.21 (0.34) - 0.39 (0.31) | −0.49 (0.47) - - | |
| Survival time | 0.15 (0.02) 0.06 (0.03) 0.21 (0.03) | 0.05 (0.03) 0.08 (0.03) 0.23 (0.04) | −0.08 (0.02) 0.03 (0.03) −0.06 (0.03) | −0.03 (0.02) −0.06 (0.03) −0.03 (0.03) | −0.11 (0.02) −0.09 (0.03) −0.04 (0.03) | −0.10 (0.02) −0.01 (0.03) −0.01 (0.03) | 0.03 (0.03) 0.18 (0.04) 0.02 (0.04) | 0.17 (0.04) - 0.06 (0.04) | −0.82 (0.59) - - | 0.04 (0.22) - −0.55 (0.31) | −0.59 (0.58) - - | |
| Cloacal viral load 2 dpe2 | −0.01 (0.04) - - | −0.06 (0.06) - - | −0.06 (0.04) - - | −0.02 (0.04) - - | −0.02 (0.04) - - | −0.04 (0.04) - - | 0.01 (0.04) - - | 0.01 (0.06) - - | −0.15 (0.04) - - | 0.78 (0.39) - - | −0.12 (0.46) - - | |
| Cloacal viral load 4 dpe2 | −0.01 (0.04) - - | −0.41 (0.05) - - | −0.01 (0.04) - 0.03 (0.05) | −0.04 (0.04) - - | 0.04 (0.04) - 0.07 (0.05) | 0.03 (0.04) - 0.08 (0.05) | 0.01 (0.05) - - | 0.10 (0.07) - 0.18 (0.09) | 0.02 (0.03) - - | 0.04 (0.05) - - | 0.43 (0.37) - - | |
| Tears viral load 6 dpi 2 | - 0.01 (0.10) - | −0.01 (0.06) 0.02 (0.09) - | 0.04 (0.05) 0.02 (0.09) - | 0.01 (0.05) 0.06 (0.09) - | 0.11 (0.05) 0.04 (0.09) - | 0.08 (0.05) - - | 0.06 (0.06) −0.39 (0.58) - | −0.01 (0.05) - - | −0.13 (0.04) - - | 0.02 (0.07) - - | 0.29 (0.08) - - | |
| Velogenic Trial Phenotypes | |||||||
|---|---|---|---|---|---|---|---|
| Growth rate pre-exposure | Proventriculus lesions | Intestine lesions | Average lesions | Natural antibody 1 | Survival time | ||
| Lasota trialphenotypes | Growth rate pre-infection | 0.64 (0.72) −0.34 (0.39) | 0.85 (0.88) −0.05 (0.55) | 0.13 (1.31) - | 0.10 (0.83) −0.18 (0.71) | 0.91 (1.27) −0.58 (1.10) | −0.04 (1.36) −0.13 (0.67) |
| Growth rate pre-infection | 0.14 (1.03) −0.48 (0.48) | 0.76 (0.92) 0.56 (0.67) | −0.49 (1.38) - | 0.004 (0.87) −0.14 (0.88) | −0.84 (2.08) −0.32 (1.26) | 0.24 (1.44) −0.65 (0.78) | |
| Anti-NDV antibody 1 | 0.98 (1.29) −0.02 (0.60) | −0.30 (1.07) −0.69 (0.82) | −0.80 (1.70) - | −0.42 (1.07) 0.43 (1.11) | −0.01 (1.76) −0.19 (1.35) | −0.24 (3.18) 0.50 (1.01) | |
| Viral load, 2 dpi 1 | 0.44 (1.64) 0.03 (0.57) | −0.02 (1.41) 0.55 (0.81) | −0.08 (2.23) - | −0.02 (1.40) 0.90 (1.06) | −0.90 (2.22) 0.56 (0.98) | −0.07 (2.25) 0.16 (0.96) | |
| Viral load, 6 dpi 1 | −0.64 (1.15) −0.15 (0.68) | −0.21 (0.99) 0.71 (1.05) | −0.88 (1.64) - | −0.65 (1.00) 0.39 (1.33) | 0.80 (1.61) 0.77 (1.22) | 0.73 (4.15) −0.89 (1.26) | |
| Viral clearance | - −0.06 (0.57) | - −0.28 (0.81) | - - | - −0.50 (1.05) | - 0.16 (1.32) | - 0.69 (1.01) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walugembe, M.; Naazie, A.; Mushi, J.R.; Akwoviah, G.A.; Mollel, E.; Mang’enya, J.A.; Wang, Y.; Chouicha, N.; Kelly, T.; Msoffe, P.L.M.; et al. Genetic Analyses of Response of Local Ghanaian Tanzanian Chicken Ecotypes to a Natural Challenge with Velogenic Newcastle Disease Virus. Animals 2022, 12, 2755. https://doi.org/10.3390/ani12202755
Walugembe M, Naazie A, Mushi JR, Akwoviah GA, Mollel E, Mang’enya JA, Wang Y, Chouicha N, Kelly T, Msoffe PLM, et al. Genetic Analyses of Response of Local Ghanaian Tanzanian Chicken Ecotypes to a Natural Challenge with Velogenic Newcastle Disease Virus. Animals. 2022; 12(20):2755. https://doi.org/10.3390/ani12202755
Chicago/Turabian StyleWalugembe, Muhammed, Augustine Naazie, James R. Mushi, George A. Akwoviah, Esther Mollel, Juliana A. Mang’enya, Ying Wang, Nadira Chouicha, Terra Kelly, Peter L. M. Msoffe, and et al. 2022. "Genetic Analyses of Response of Local Ghanaian Tanzanian Chicken Ecotypes to a Natural Challenge with Velogenic Newcastle Disease Virus" Animals 12, no. 20: 2755. https://doi.org/10.3390/ani12202755