
Essential and Toxic Mineral Content and Fatty Acid Profile of Colostrum in Dairy Sheep

 , ,
, ,  ,
,  ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Practices
2.2. Animals and Experimental Design
2.3. Milk Sampling
2.4. Colostrum Sampling
2.5. Determination of IgG, Protein, Fat Content, and Fatty Acid Profile
2.6. Determination of Minerals in Colostrum and Milk
2.7. Statistical Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Argüello, A.; Castro, N.; Capote, J.; Tyler, J.W.; Holloway, N.M. Effect of colostrum administration practices on serum IgG in goat kids. Livest. Prod. Sci. 2004, 90, 235–239. [Google Scholar] [CrossRef]
- Castro, N.; Capote, J.; Bruckmaier, R.M.; Arguello, A. Management effects on colostrogenesis in small ruminants: A review. J. Appl. Anim. Res. 2011, 39, 85–93. [Google Scholar] [CrossRef]
- Hurley, W.L.; Theil, P.K. Perspectives on immunoglobulins in colostrum and milk. Nutrients. 2011, 3, 442–474. [Google Scholar] [CrossRef] [Green Version]
- Sfacteria, A.; Lanteri, G.; Agricola, S.; Ferraro, S.; Macrì, B.; Mazzullo, G. Miodistrofia enzootica degli agnelli: Indagini clinico-patologiche in un allevamento siciliano. Large Anim. Rev. 2009, 15, 211–214. [Google Scholar]
- Rossi, L.; Valdez Lumbreras, A.E.; Vagni, S.; Dell’Anno, M.; Bontempo, V. Nutritional and functional properties of colostrum in puppies and kittens. Animals 2021, 11, 3260. [Google Scholar] [CrossRef] [PubMed]
- Chandwe, K.; Kelly, P. Colostrum therapy for human gastrointestinal health and disease. Nutrients 2021, 13, 1956. [Google Scholar] [CrossRef] [PubMed]
- Mehra, R.; Garhwal, R.; Sangwan, K.; Guiné, R.; Lemos, E.T.; Buttar, H.S.; Visen, P.; Kumar, N.; Bhardwaj, A.; Kumar, H. Insights into the research trends on bovine colostrum: Beneficial health perspectives with special reference to manufacturing of functional foods and feed supplements. Nutrients 2022, 14, 659. [Google Scholar] [CrossRef] [PubMed]
- Davison, G.; Marchbank, T.; March, D.S.; Thatcher, R.; Playford, R.J. Zinc carnosine works with bovine colostrum in truncating heavy exercise-induced increase in gut permeability in healthy volunteers. Am. J. Clin. Nutr. 2016, 104, 526–536. [Google Scholar] [CrossRef] [Green Version]
- Page, P.; Sherwin, G.; Sampson, R.; Phillips, K.; Lovatt, F. Ewe colostrum quality on commercial Welsh sheep farms. Livestock 2022, 27, 40–46. [Google Scholar] [CrossRef]
- Regione Autonoma della Sardegna. Scheda Identificativa dei Prodotti Agroalimentari Tradizionali della Sardegna ai Sensi Dell’articolo 8, D.lgs. n. 173/98, Articolo 2 D.M. n. 350/99. Available online: https://www.regione.sardegna.it/documenti/1_38_20160920145750.pdf (accessed on 20 July 2022).
- Pulina, G.; Atzori, A.S.; Dimauro, C.; Ibba, I.; Gaias, G.F.; Correddu, F. The milk fingerprint of Sardinian dairy sheep: Quality and yield of milk used for Pecorino Romano PDO. cheese production on population-based 5-year survey. Ital. J. Anim. Sci. 2021, 20, 171–180. [Google Scholar] [CrossRef]
- Su, C.-K.; Chiang, B.H. Extraction of immunoglobulin-G from colostral whey by reverse micelles. J. Dairy Sci. 2003, 86, 1639–1645. [Google Scholar] [CrossRef]
- Zarrilli, A.; Micera, E.; Lacarpia, N.; Lombardi, P.; Pero, M.E.; Pelagalli, A.; D’Angelo, D.; Mattia, M.; Avallone, L. Evaluation of ewe colostrum quality by estimation of enzyme activity levels. Revue Médecine Vétérinaire 2003, 154, 521–523. [Google Scholar]
- AOAC International. Official Methods of Analysis, 16th ed.; AOAC Int.: Arlington, VA, USA, 1995. [Google Scholar]
- AOAC International. Official Methods of Analysis, 17th ed.; AOAC Int.: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Tsiplakou, E.; Mitsiopoulou, C.; Mavrommatis, A.; Karaiskou, C.; Chronopoulou, E.G.; Mavridis, G.; Sotirakoglou, K.; Labrou, N.E.; Zervas, G. Effect of under- and overfeeding on sheep and goat milk and plasma enzymes activities related to oxidation. J. Anim. Physiol. Anim. Nutr. 2017, 102, 288. [Google Scholar]
- Nudda, A.; McGuire, M.A.; Battacone, G.; Pulina, G. Seasonal variation in conjugated linoleic acid and vaccenic acid in milk fat of sheep and its transfer to cheese and ricotta. J. Dairy Sci. 2005, 88, 1311–1319. [Google Scholar] [CrossRef] [Green Version]
- FIL-IDF. International Dairy Federation. Milk fat. In Preparation of Fatty Acid Methyl Esters; International Dairy Federation: Brussels, Belgium, 1999. [Google Scholar]
- Nudda, A.; Battacone, G.; Atzori, A.S.; Dimauro, C.; Rassu, P.G.; Nicolussi, P.; Bonelli, P.; Pulina, G. Effect of extruded linseed supplementation on blood metabolic profile and milk performance of Saanen goats. Animal 2013, 7, 1464–1471. [Google Scholar] [CrossRef]
- Schennink, A.; Heck, M.L.; Bovenhuis, H.; Visker, M.H.P.W.; van Valenberg, H.J.F.; van Arendonk, J.A.M. Milk fatty acid unsaturation: Genetic parameters and effects of stearoyl-CoA desaturase (SCD1) and acyl CoA: Diacylglycerol acyltransferase 1 (DGAT1). Int. J. Dairy Sci. 2008, 91, 2135–2143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessler, E.C.; Bruckmaier, R.M.; Gross, J.J. Immunoglobulin G content and colostrum composition of different goat and sheep breeds in Switzerland and Germany. J. Dairy Sci. 2019, 102, 5542–5549. [Google Scholar] [CrossRef]
- Kessler, E.C.; Bruckmaier, R.M.; Gross, J.J. Short communication: Comparative estimation of colostrum quality by Brix refractometry in bovine, caprine, and ovine colostrum. J. Dairy Sci. 2021, 104, 2438–2444. [Google Scholar] [CrossRef] [PubMed]
- Turquino, C.F.; Flaiban, K.K.M.C.; Lisbôa, J.A.N. Passive transfer of immunity in meat lambs reared in the tropics on extensive management. Pesquisa Veterinária Brasileira 2011, 31, 3. [Google Scholar] [CrossRef] [Green Version]
- Alves, A.C.; Alves, N.G.; Ascari, I.J.; Junqueir, F.B.; Coutinho, A.S.; Lima, R.R.; Pérez, J.R.O.; De Paula, S.O.; Furusho-Garcia, I.F.; Abreu, L.R. Colostrum composition of Santa Inês sheep and passive transfer of immunity to lambs. J. Dairy Sci. 2015, 98, 3706–3716. [Google Scholar] [PubMed] [Green Version]
- Salman, S.; Dinse, D.; Khol-Parisini, A.; Schafft, H.; Lahrssen-Wiederholt, M.; Schreiner, M.; Scharek-Tedin, L.; Zentek, J. Colostrum and milk selenium, antioxidative capacity and immune status of dairy cows fed sodium selenite or selenium yeast. Arch. Anim. Nutr. 2013, 67, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Kachuee, R.; Abdi-Benemar, H.; Mansoori, Y.; Sánchez-Aparicio, P.; Seifdavati, J.; Elghandour, M.; Guillén, R.J.; Salem, A. Effects of sodium selenite, L-selenomethionine, and selenium nanoparticles during late pregnancy on selenium, zinc, copper, and iron concentrations in Khalkhali goats and their kids. Biol. Trace Elem. Res. 2019, 191, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Guiso, M.F.; Battacone, G.; Canu, L.; Deroma, M.; Langasco, I.; Sanna, G.; Pulina, G.; Nudda, A. Seasonal variation of selenium, manganese, zinc, copper and toxic metals concentration in milk of sarda dairy ewes. Sheep and goat management and reproduction-session 054. In Book of Abstract of the 72 Annual Meeting of the European Federation of Animal Science; EAAP: Davos, Switzerland, 2021; p. 514. [Google Scholar]
- Van Dael, P.; Shen, L.; Van Renterghem, R.; Deelstra, H. Selenium content of sheep’s milk and its distribution in protein fractions. Eur. Food Res. Technol. 1993, 196, 536–539. [Google Scholar]
- Beretta, C. (Ed.) Tossicologia Veterinaria; Casa Editrice Ambrosiana: Milan, Italy, 1998; pp. 183–184. [Google Scholar]
- Stewart, W.C.; Bobe, G.; Vorachek, W.R.; Stang, B.V.; Pirelli, G.J.; Mosher, W.D.; Hall, J.A. Organic and inorganic selenium: IV. Passive transfer of immunoglobulin from ewe to lamb. J. Anim. Sci. 2013, 91, 1791–1800. [Google Scholar] [CrossRef] [PubMed]
- Wallace, L.G.; Bobe, G.; Vorachek, W.R.; Dolan, B.P.; Estill, C.T.; Pirelli, G.J.; Hall, J.A. Effects of feeding pregnant beef cows selenium-enriched alfalfa hay on selenium status and antibody titers in their newborn calves. J. Anim. Sci. 2017, 95, 2408–2420. [Google Scholar] [CrossRef]
- Melnikov, P.; da Cruz Montes Moura, A.J.; Batista Palhares, D.; Martinbianco de Figueiredo, C.S. Zinc and copper in colostrum. Indian Pediatr. 2007, 44, 355–357. [Google Scholar]
- De la Fuente, M.A.; Olano, A.; Juàrez, M. Distribution of calcium, magnesium, phosphorus, zinc, manganese, copper and iron between the soluble and colloidal phases of ewe’s and goat’s milk. Lait 1997, 77, 515–520. [Google Scholar] [CrossRef] [Green Version]
- Mills, C.F.; Dalgarno, A.C.; Williams, R.B.; Quarterman, J. Zinc deficiency and the zinc requirements of calves and lambs. Br. J. Nutr. 1967, 21, 751–768. [Google Scholar] [CrossRef] [Green Version]
- Goran, G.V.; Crivineanu, V.; Rotaru, E.; Tudoreanu, L.; Hanganu, A. Dynamics of some mineral elements in sheep colostrum. Bul. UASVM Vet. Med. 2010, 67, 81–87. [Google Scholar]
- Roshanzamir, H.; Rezaei, J.; Fazaeli, H. Colostrum and milk performance, and blood immunity indices and minerals of Holstein cows receiving organic Mn, Zn and Cu sources. Anim. Nutr. 2020, 6, 61–68. [Google Scholar] [CrossRef]
- Senthilkumar, P.; Nagalakshmi, D.; Ramana Reddy, Y.; Sudhakar, K. Effect of different level and source of copper supplementation on immune response and copper dependent enzyme activity in lambs. Trop. Anim. Health Prod. 2009, 41, 645–653. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, 49, 5–24. [Google Scholar]
- Abba, B.; Ali, H.A.; Chamba, G.; Sanda, F.S.; Modu, S. Determination of some heavy metals and proximate composition of camel, cow, goat and sheep milk. ChemSearch J. 2021, 12, 50–54. [Google Scholar]
- Amaro, M.A.; Sanchez, P.J.; Moreno, R.; Zurera, G. Nickel content in raw cow’s, ewe’s and goat’s milk. Lait 1998, 78, 699–706. [Google Scholar] [CrossRef] [Green Version]
- Fereníka, M.; Ebringer, L. Modulatory effects of selenium and zinc on the immune system. Folia Microbiol. 2003, 48, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Srivastava, A.K.; Bhatia, A.K. Studies on immune response in copper toxicity in sheep. J. Immunol. Immunopathol. 2002, 4, 93–96. [Google Scholar]
- Cekic, S.D.; Demir, A.; Baskan, K.S.; Tütem, E.; Apak, R. Determination of total antioxidant capacity of milk by CUPRAC and ABTS methods with separate characterisation of milk protein fractions. J. Dairy Res. 2015, 82, 177–184. [Google Scholar] [CrossRef]
- Chen, J.; Lindmark-Mansson, H.; Gorton, L.; Åkesson, B. Antioxidant capacity of cow milk as assayed by spectrophotometric and amperometric methods. Int. Dairy J. 2003, 13, 927–935. [Google Scholar] [CrossRef]
- Stobiecka, M.; Król, J.; Brodziak, A. Antioxidant activity of milk and dairy products. Animals 2022, 12, 245. [Google Scholar] [CrossRef] [PubMed]
- Annichiarico, G.; Taibi, L. La Nutrizione Minerale e Vitaminica ed i Fabbisogni Idrici. In L’Alimentazione Degli Ovini Da Latte; Pulina, G., Ed.; Avenue media: Bologna, Italy, 2001; pp. 67–68. [Google Scholar]
- Wilms, J.N.; Hare, K.S.; Fischer-Tlustos, A.J.; Vahmani, P.; Dugan, M.; Leal, L.N.; Steele, M.A. Fatty acid profile characterization in colostrum, transition milk, and mature milk of primi- and multiparous cows during the first week of lactation. J. Dairy Sci. 2022, 105, 4692–4710. [Google Scholar] [CrossRef]
- Cesarani, A.; Gaspa, G.; Correddu, F.; Cellesi, M.; Dimauro, C.; Macciotta, N.P.P. Genomic selection of milk fatty acid composition in Sarda dairy sheep: Effect of different phenotypes and relationship matrices on heritability and breeding value accuracy. J. Dairy Sci. 2019, 102, 3189–3203. [Google Scholar] [CrossRef] [Green Version]
- Correddu, F.; Cesarani, A.; Dimauro, C.; Gaspa, G.; Macciotta, N.N.P. Principal component and multivariate factor analysis of detailed sheep milk fatty acid profile. J. Dairy Sci. 2021, 104, 5079–5094. [Google Scholar] [CrossRef] [PubMed]
- Ruxton, C.; Reed, S.; Simpson, M.; Millington, K. The health benefits of omega-3 polyunsaturated fatty acids: A review of the evidence. J. Hum. Nutr. Diet. 2007, 20, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Basak, S.; Mallick, R.; Banerjee, A.; Pathak, S.; Duttaroy, A.K. Maternal supply of both arachidonic and docosahexaenoic acids is required for optimal neurodevelopment. Nutrients 2021, 16, 2061. [Google Scholar] [CrossRef] [PubMed]
- Kudo, N.; Nakagawa, Y.; Waku, K.; Kawashima, Y.; Kozuka, H. Prevention by zinc of cadmium inhibition of stearoyl-CoA desaturase in rat-liver. Toxicology 1991, 68, 133–142. [Google Scholar] [CrossRef]
- Pigeon, C.; Legrand, P.; Leroyer, P.; Bouriel, M.; Turlin, B.; Brissot, P.; Loréal, O. Stearoyl coenzyme a desaturase 1 expression and activity are increased in the liver during iron overload. Biochim. Biophys. Acta 2001, 1535, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Karlengen, I.J.; Taugbøl, O.; Salbu, B.; Aastveit, A.H.; Harstad, O.M. Effect of different levels of supplied cobalt on the fatty acid composition of bovine milk. Br. J. Nutr. 2013, 109, 834–843. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Colostrum Composition | |||||
|---|---|---|---|---|---|
| Farm | TP (%) | IgG (g dm−3) | Fat (%) | FRAP (µmol Ascorbic Acid dm−3) | ABTS (% Inhibition) |
| A | 17 | 40 | 5.3 c | 3.5 | 20 b |
| B | 16 | 40 | 7.6 abc | 3.4 | 20 b |
| C | 14 | 30 | 7.7 abc | nd | nd |
| D | 15 | 40 | 9.6 a | 2.3 | 20 b |
| E | 15 | 40 | 6.9 abc | 2.4 | 20 b |
| F | 19 | 50 | 5.5 bc | 3.1 | 20 b |
| G | 14.5 | 40 | 9.1 ab | 2.2 | 40 a |
| H | 16 | 40 | 10 a | 3.4 | 50 a |
| Mean of all samples | 16 | 40 | 7.8 | 2.9 | 30 |
| SD | 5 | 20 | 3.2 | 1.4 | 10 |
| Min | 7.1 | 7.9 | 2.5 | 0.7 | 10 |
| Max | 29 | 105 | 18 | 8.5 | 58 |
| Pvalue | 0.511 | 0.379 | <0.001 | 0.120 | <0.001 |
| SEM | 0.54 | 2.20 | 0.36 | 0.15 | 1.53 |
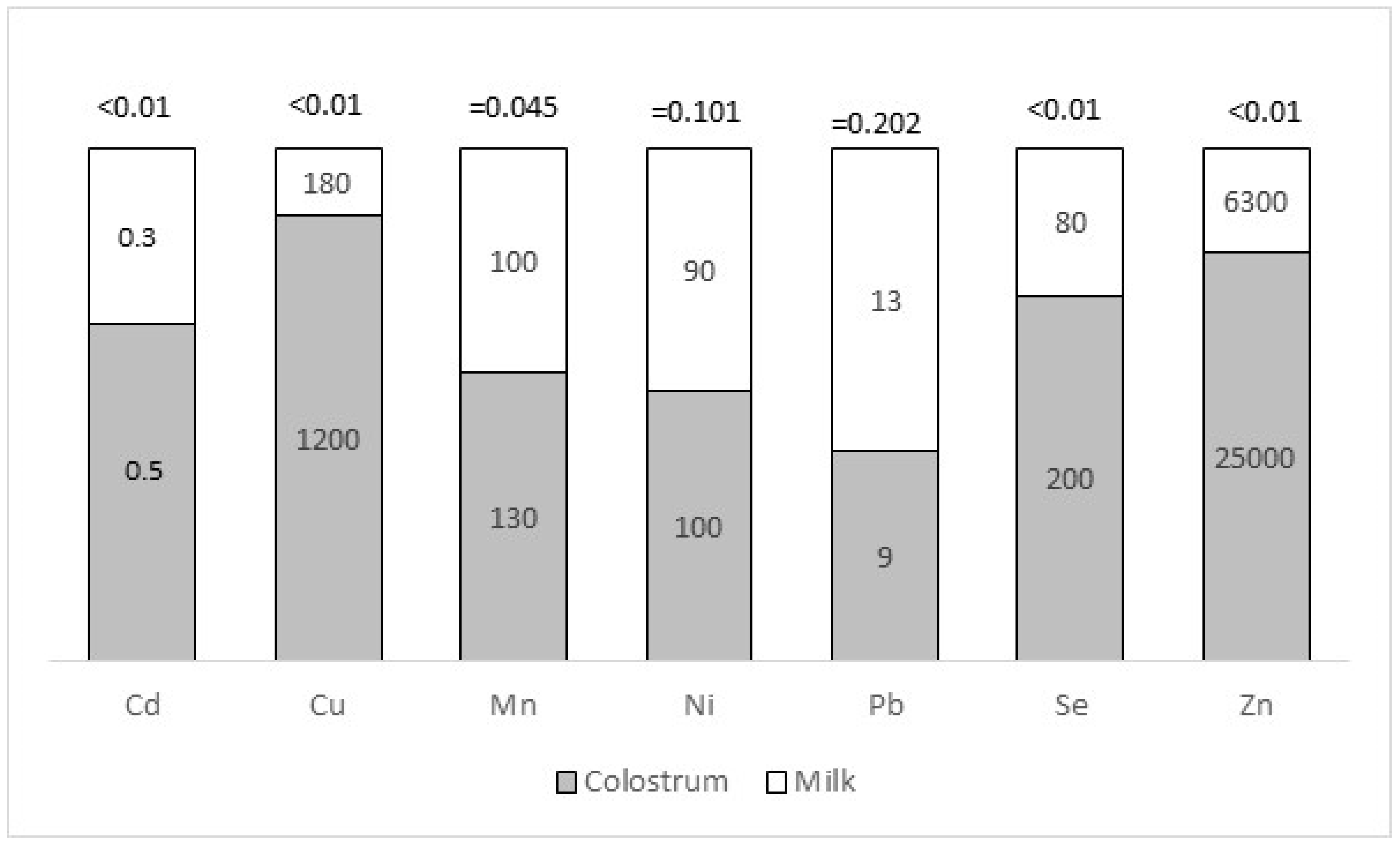
| Minerals | |||||||
|---|---|---|---|---|---|---|---|
| Farm | Cd | Cu | Mn | Ni | Pb | Se | Zn |
| A | 0.4 ab | 660 d | 140 b | 90 ab | 10 | 300 ab | 26,000 |
| B | 0.8 a | 1200 abc | 70 c | 110 ab | 12 | 200 ab | 32,000 |
| C | 0.5 ab | 1600 a | 240 a | 80 b | 7 | 200 ab | 23,000 |
| D | 0.6 ab | 880 bcd | 250 a | 120 a | 11 | 300 ab | 23,000 |
| E | 0.4 ab | 1400 ab | 74 bc | 80 b | 7 | 200 ab | 18,000 |
| F | 0.6 ab | 670 cd | 74 bc | 80 b | 7 | 100 b | 27,000 |
| G | 0.6 ab | 1300 abc | 100 bc | 110 ab | 9 | 350 a | 25,000 |
| H | 0.3 b | 1600 a | 60 c | 100 ab | 11 | 200 ab | 22,000 |
| Mean | 0.5 | 1200 | 130 | 100 | 9 | 200 | 25,000 |
| SD | 0.3 | 600 | 90 | 30 | 5 | 200 | 11,000 |
| Min | 0.1 | 130 | 33 | 39 | 3 | 20 | 5000 |
| Max | 1.5 | 2800 | 400 | 190 | 32 | 1000 | 57,000 |
| p-value | 0.0083 | <0.0001 | <0.0001 | 0.0016 | 0.0417 | 0.0282 | 0.217 |
| Se | Cu | Mn | Zn | Ni | Pb | Cd | TP | IgG | Fat | FRAP | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cu | 0.053 | 1 | |||||||||
| Mn | 0.220 | −0.124 | 1 | ||||||||
| Zn | 0.469 ** | −0.115 | 0.114 | 1 | |||||||
| Ni | 0.428 ** | −0.087 | 0.336 ** | 0.486 ** | 1 | ||||||
| Pb | 0.127 | −0.020 | −0.003 | 0.135 | 0.433 ** | 1 | |||||
| Cd | 0.041 | 0.017 | −0.035 | 0.154 | 0.263 * | 0.059 | 1 | ||||
| NT | 0.492 ** | −0.208 | 0.113 | 0.680 ** | 0.359 ** | 0.039 | 0.051 | 1 | |||
| IgG | 0.447 ** | −0.254 * | −0.005 | 0.555 ** | 0.250 * | 0.015 | 0.107 | 0.912 ** | 1 | ||
| Fat | −0.177 | 0.247 * | 0.029 | −0.063 | 0.058 | 0.014 | −0.021 | −0.331 * | 0.412 ** | 1 | |
| FRAP | 0.135 | −0.111 | −0.038 | 0.310 ** | −0.028 | −0.126 | −0.058 | 0.285 * | 0.256 * | −0.035 | 1 |
| ABTS | 0.051 | 0.39 ** | −0.219 | −0.072 | −0.048 | 0.087 | −0.099 | −0.063 | −0.182 | 0.44 ** | −0.025 |
| Type | p-Value | ||||
|---|---|---|---|---|---|
| Colostrum | Milk | SEM | Type | Farm | |
| Fat, % | 8 | 5.8 | 0.4922 | 0.0081 | 0.0021 |
| FA (% on Total FAs) | |||||
| Short chain FA | |||||
| C4:0 | 1.8 | 2.4 | 0.0656 | <0.0001 | 0.0017 |
| C6:0 | 0.8 | 2.0 | 0.0417 | <0.0001 | 0.0006 |
| C7:0 | 0.05 | 0.08 | 0.0048 | 0.0005 | <0.0001 |
| C8:0 | 0.6 | 2.1 | 0.0464 | <0.0001 | 0.0063 |
| C9:0 | 0.10 | 0.15 | 0.0095 | 0.0441 | <0.0001 |
| C10:0 | 1.8 | 7 | 0.1744 | <0.0001 | 0.0031 |
| C10:1 | 0.06 | 0.09 | 0.0052 | 0.0001 | 0.0226 |
| Medium chain FA | |||||
| C11:0 | 0.12 | 0.32 | 0.0126 | <0.0001 | 0.0041 |
| C12:0 | 2.0 | 4.2 | 0.1112 | <0.0001 | 0.0097 |
| isoC13:0 | 0.01 | 0.02 | 0.0008 | <0.0001 | 0.0441 |
| anteisoC13:0 | 0.03 | 0.04 | 0.0023 | 0.0064 | 0.0009 |
| isoC14:0 | 0.06 | 0.12 | 0.0039 | <0.0001 | 0.3183 |
| C14:0 | 11 | 11 | 0.4014 | 0.9561 | 0.0001 |
| isoC15:0 | 0.18 | 0.31 | 0.0071 | <0.0001 | 0.0398 |
| anteisoC15:0 | 0.20 | 0.54 | 0.0116 | <0.0001 | 0.1539 |
| C14:1c9 | 0.4 | 0.17 | 0.0353 | <0.0001 | <0.0001 |
| C15:0 | 0.6 | 1.1 | 0.0251 | <0.0001 | 0.0002 |
| C15:1 | 0.03 | 0.08 | 0.003 | <0.0001 | <0.0001 |
| isoC16:0 | 0.20 | 0.35 | 0.0083 | <0.0001 | 0.1687 |
| C16:0 | 29 | 24 | 0.8182 | <0.0001 | <0.0001 |
| isoC17:0 | 0.42 | 0.51 | 0.0132 | <0.0001 | <0.0001 |
| anteisoC17:0 | 0.47 | 0.48 | 0.0155 | 0.7088 | <0.0001 |
| C16:1c9 | 1.6 | 0.8 | 0.1189 | <0.0001 | 0.0001 |
| C17:0 | 0.8 | 0.72 | 0.0264 | 0.0002 | 0.0004 |
| isoC18:0 | 0.12 | 0.07 | 0.005 | <0.0001 | 0.0012 |
| C17:1c9 | 0.4 | 0.19 | 0.0148 | <0.0001 | 0.0006 |
| Long chain FA | |||||
| C18:0 | 7 | 10 | 0.3547 | <0.0001 | 0.0162 |
| C18:1t4-8 | 0.21 | 0.28 | 0.0082 | <0.0001 | 0.0037 |
| C18:1t9 | 0.21 | 0.25 | 0.009 | 0.0004 | <0.0001 |
| C18:1t10 | 0.3 | 0.4 | 0.0273 | <0.0001 | 0.0367 |
| C18:1t11 | 0.7 | 1.6 | 0.0752 | <0.0001 | 0.0059 |
| C18:1t12 | 0.23 | 0.4 | 0.0162 | <0.0001 | 0.008 |
| C18:1t13:t14 | 0.3 | 1.1 | 0.0391 | <0.0001 | 0.0306 |
| C18:1c9 | 30 | 17 | 1.0669 | <0.0001 | <0.0001 |
| C18:2n6 (LA) | 2.5 | 2.1 | 0.1102 | 0.0001 | <0.0001 |
| C20:0 | 0.22 | 0.26 | 0.0078 | <0.0001 | <0.0001 |
| C18:3n6 | 0.05 | 0.05 | 0.0037 | 0.0631 | 0.0044 |
| C20:1c9 | 0.03 | 0.02 | 0.0012 | <0.0001 | 0.0002 |
| C18:3n3 (ALA) | 0.4 | 0.7 | 0.0349 | <0.0001 | <0.0001 |
| CLAc9t11 | 0.7 | 0.8 | 0.0422 | 0.0043 | <0.0001 |
| CLAt10c12 | 0.03 | 0.05 | 0.0031 | <0.0001 | 0.0003 |
| CLAt12t14 | 0.01 | 0.04 | 0.0021 | <0.0001 | 0.0065 |
| CLAt11t13 | 0.03 | 0.06 | 0.0034 | <0.0001 | 0.042 |
| CLAt9t11 | 0.02 | 0.03 | 0.002 | <0.0001 | 0.0098 |
| C18:4n3 | 0.01 | 0.01 | 0.0005 | 0.0002 | 0.0229 |
| C20:2n6 | 0.03 | 0.02 | 0.0011 | <0.0001 | 0.3811 |
| C20:3n9 | 0.07 | 0.10 | 0.0051 | <0.0001 | 0.4713 |
| C22:0 | 0.07 | 0.15 | 0.0048 | <0.0001 | 0.0035 |
| C20:3n6 | 0.04 | 0.03 | 0.0014 | <0.0001 | 0.0006 |
| C20:4n6 (ARA) | 0.31 | 0.14 | 0.0116 | <0.0001 | <0.0001 |
| C23:0 | 0.02 | 0.08 | 0.0025 | <0.0001 | 0.0252 |
| EPA | 0.07 | 0.05 | 0.0033 | <0.0001 | <0.0001 |
| DPA | 0.17 | 0.09 | 0.0084 | <0.0001 | <0.0001 |
| DHA | 0.06 | 0.03 | 0.0034 | <0.0001 | <0.0001 |
| Groups of FA | |||||
| SCFA | 5 | 14 | 0.2799 | <0.0001 | 0.0015 |
| MCFA | 48 | 46 | 1.3171 | 0.25 | <0.0001 |
| LCFA | 47 | 40 | 1.4586 | 0.0003 | <0.0001 |
| SFA | 58 | 69 | 1.1462 | <0.0001 | <0.0001 |
| MUFA | 37 | 25 | 1.0343 | <0.0001 | <0.0001 |
| PUFA | 6 | 6.1 | 0.1953 | 0.152 | 0.0103 |
| UFA | 42 | 31 | 1.1463 | <0.0001 | <0.0001 |
| OCFA | 1.7 | 2.4 | 0.0455 | <0.0001 | 0.0014 |
| BCFA | 1.7 | 2.4 | 0.0458 | <0.0001 | 0.0034 |
| OBCFA | 3.4 | 4.9 | 0.0769 | <0.0001 | 0.0023 |
| PUFA6 | 3.0 | 2.5 | 0.1208 | <0.0001 | <0.0001 |
| PUFA3 | 0.8 | 0.9 | 0.0464 | 0.0078 | <0.0001 |
| n6/n3 | 4 | 3 | 0.3021 | <0.0001 | <0.0001 |
| n3/n6 | 0.27 | 0.4 | 0.0213 | <0.0001 | <0.0001 |
| CLA | 0.8 | 1.1 | 0.0479 | <0.0001 | <0.0001 |
| TFA | 3.4 | 6 | 0.2146 | <0.0001 | 0.0012 |
| TFA (without VA) | 2.7 | 5 | 0.1519 | <0.0001 | 0.0013 |
| Indexes | |||||
| AI | 1.9 | 2.3 | 0.1329 | 0.0071 | <0.0001 |
| TI | 1.9 | 2.1 | 0.1237 | 0.1187 | <0.0001 |
| h/H | 0.9 | 0.60 | 0.049 | <0.0001 | <0.0001 |
| DI C10:1 | 4 | 1.3 | 0.2513 | <0.0001 | <0.0001 |
| DI C14:1 | 3 | 1.5 | 0.178 | <0.0001 | 0.0018 |
| DI C16:1 | 5 | 2.9 | 0.2219 | <0.0001 | 0.0081 |
| DI C18:1 | 80 | 63 | 0.7329 | <0.0001 | 0.1824 |
| DI CLA | 51 | 35 | 0.9838 | <0.0001 | 0.5566 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guiso, M.F.; Battacone, G.; Canu, L.; Deroma, M.; Langasco, I.; Sanna, G.; Tsiplakou, E.; Pulina, G.; Nudda, A. Essential and Toxic Mineral Content and Fatty Acid Profile of Colostrum in Dairy Sheep. Animals 2022, 12, 2730. https://doi.org/10.3390/ani12202730
Guiso MF, Battacone G, Canu L, Deroma M, Langasco I, Sanna G, Tsiplakou E, Pulina G, Nudda A. Essential and Toxic Mineral Content and Fatty Acid Profile of Colostrum in Dairy Sheep. Animals. 2022; 12(20):2730. https://doi.org/10.3390/ani12202730
Chicago/Turabian StyleGuiso, Maria Francesca, Gianni Battacone, Linda Canu, Mario Deroma, Ilaria Langasco, Gavino Sanna, Eleni Tsiplakou, Giuseppe Pulina, and Anna Nudda. 2022. "Essential and Toxic Mineral Content and Fatty Acid Profile of Colostrum in Dairy Sheep" Animals 12, no. 20: 2730. https://doi.org/10.3390/ani12202730