
Physiological and Biochemical Characteristics of Rainbow Trout with Severe, Moderate and Asymptomatic Course of Vibrio anguillarum Infection
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal and Water Collection
- I—relative weight of organ
- O—organ mass, g
- F—fish mass, g

2.2. Fish Pathology Characterization (Diagnostic Criteria)
2.3. Hematological Parameters
2.4. Expression of Immune-Related Genes
2.5. Grouping Fish according to the Severity of the Disease
2.6. Identification of the Pathogen Using 16S rRNA Gene Amplicon Sequencing
2.7. Plasma Proteome Analysis
3. Results
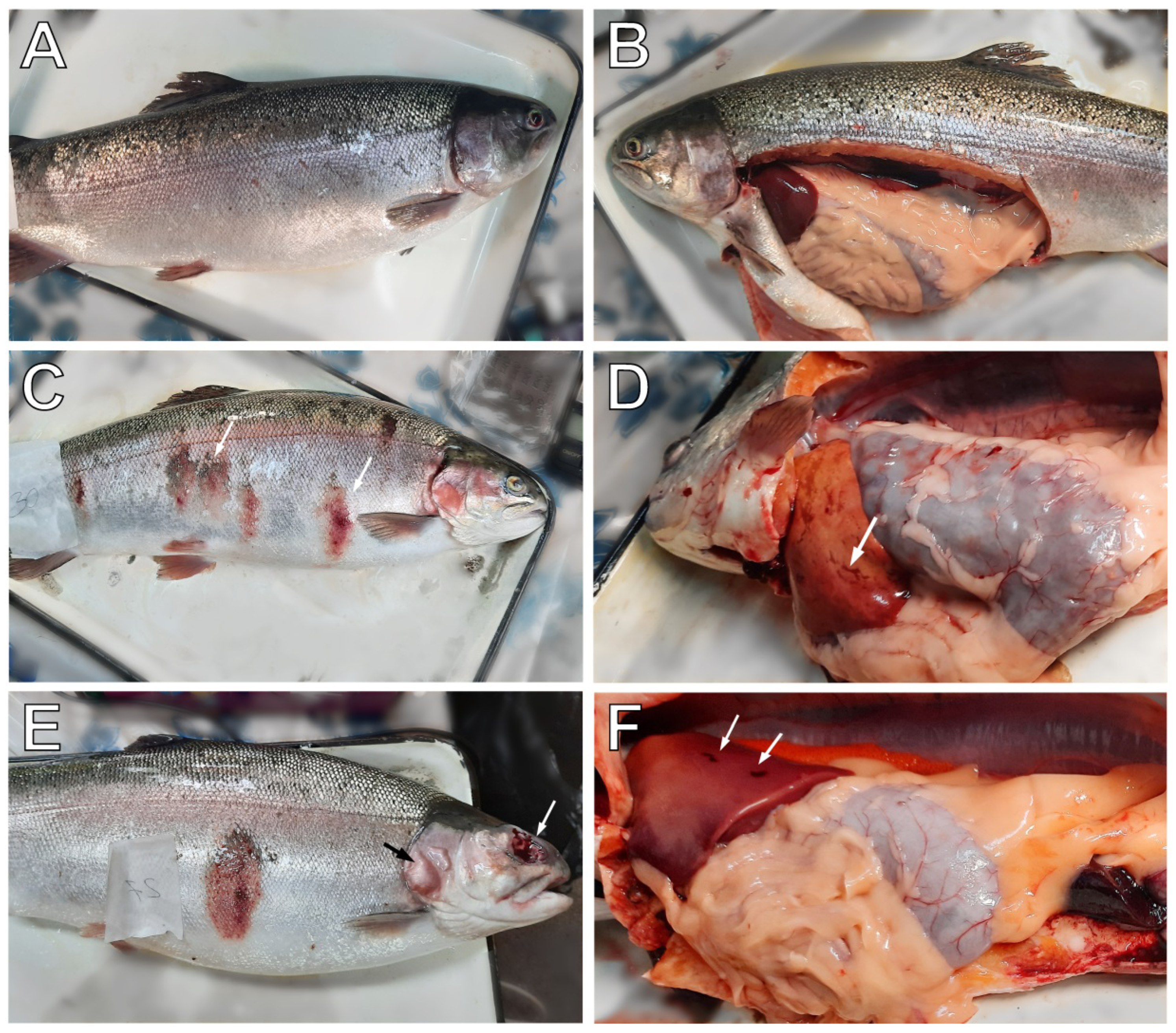
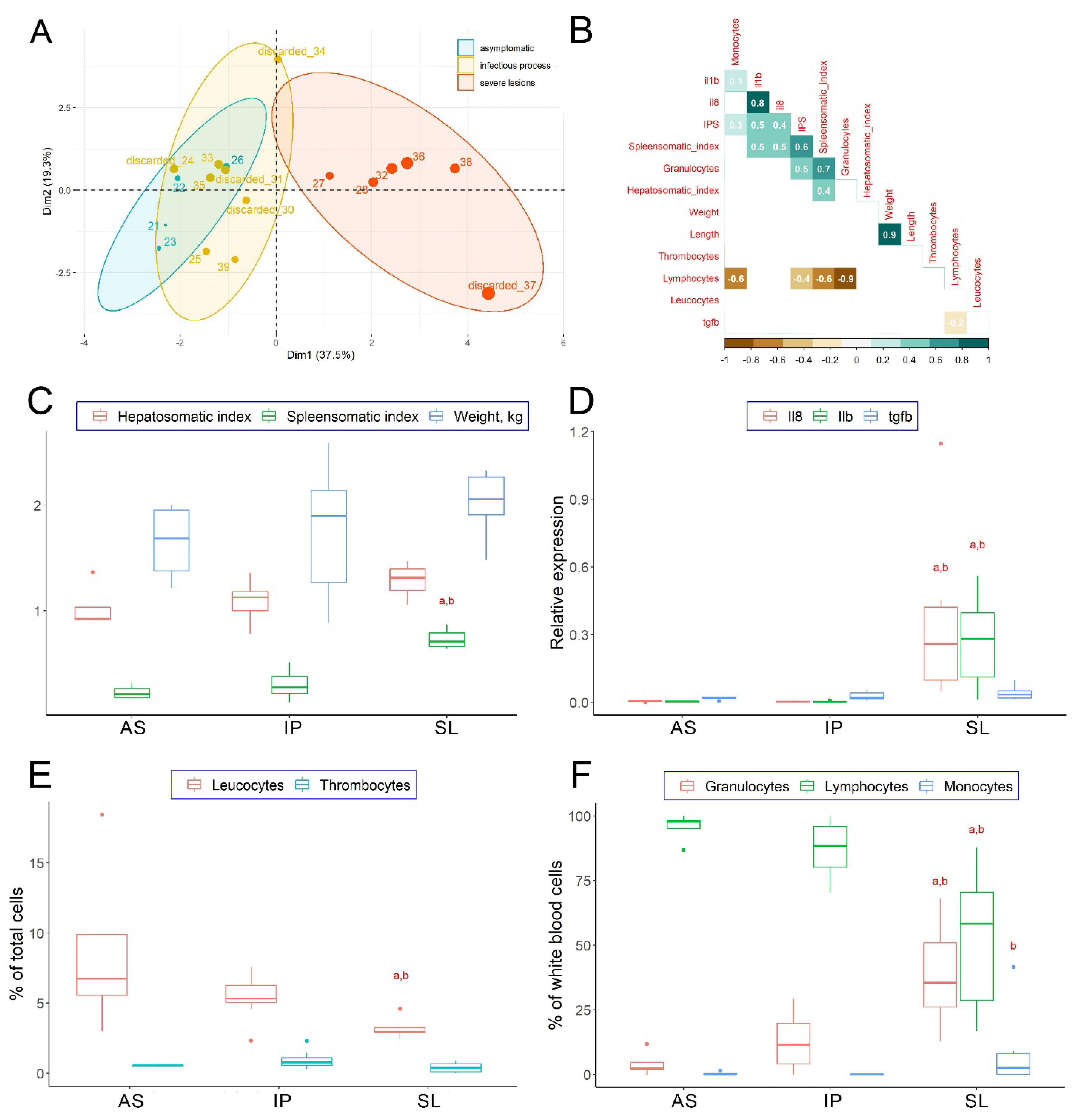
3.1. Clinical Examination of Collected Fish
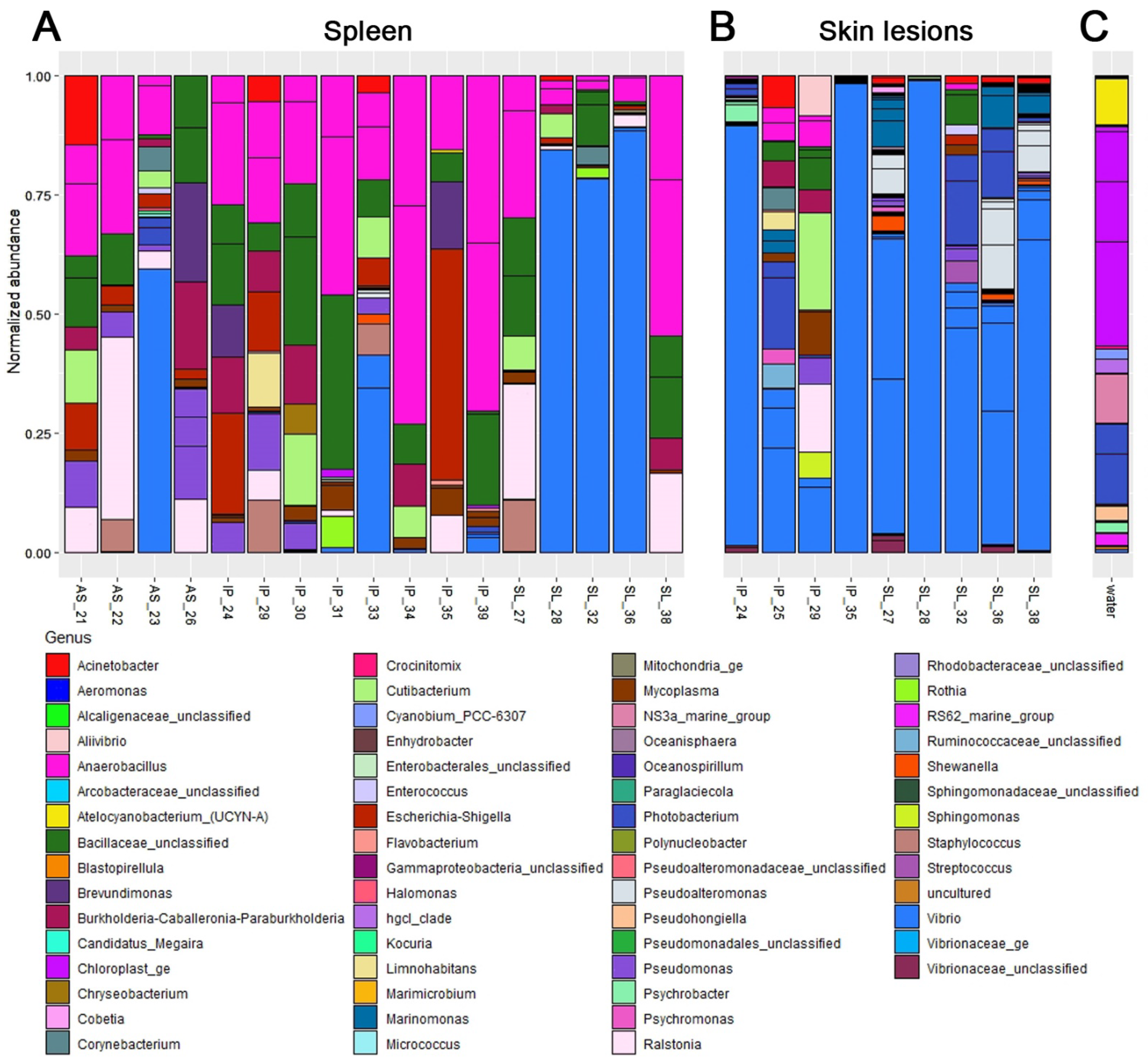
3.2. Molecular Identification of Pathogen
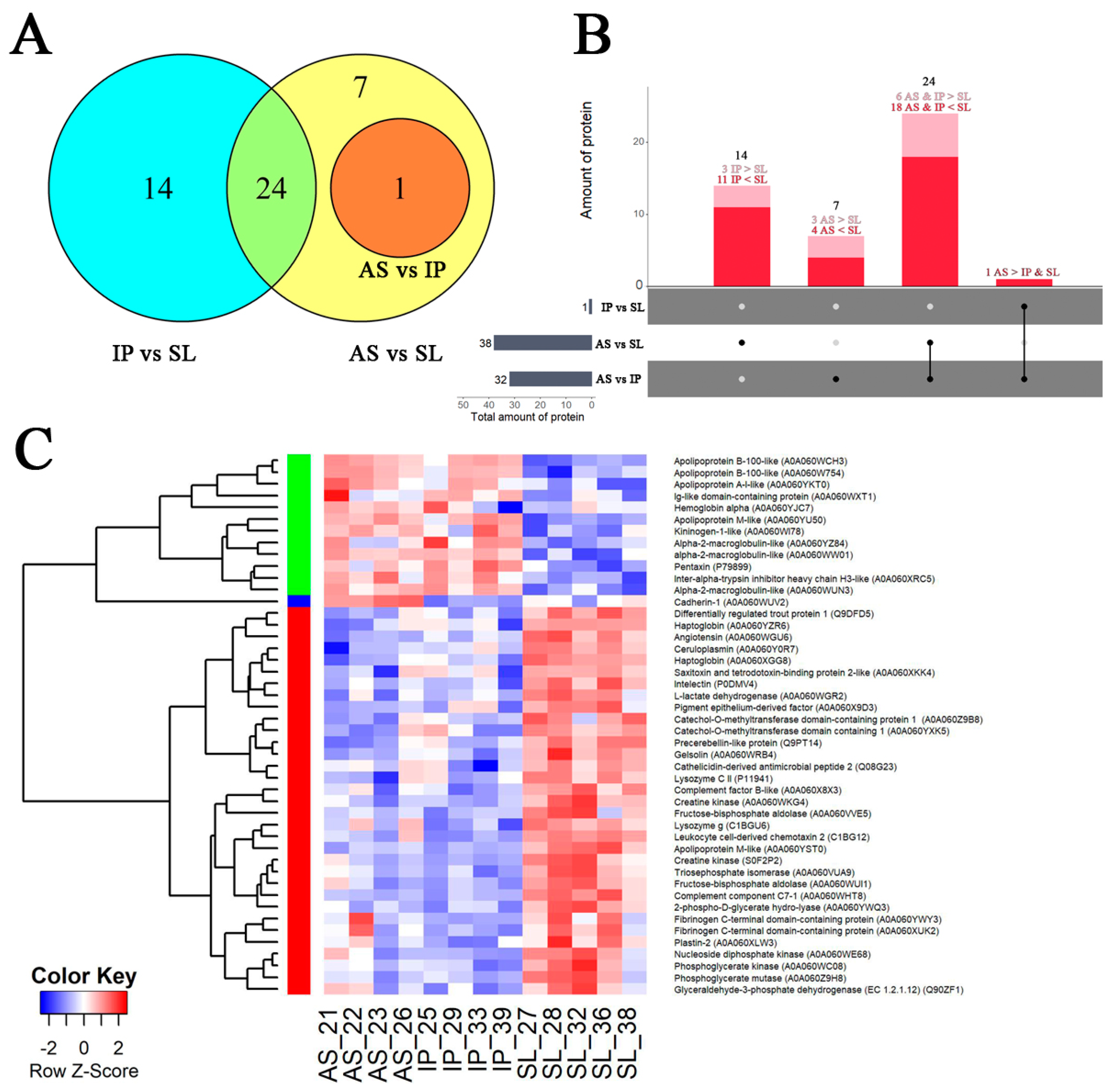
3.3. Proteomic Analysis
4. Discussion
4.1. Molecular Mechanisms of Individual Resistance to Natural V. anguillarum
4.2. Molecular Characterization of Severely Affected Fish
4.2.1. Serum Markers of Tissue Damage
4.2.2. Antigen Recognition and Humoral Immunity Regulation
4.2.3. Blood Coagulation System
4.2.4. Anti-Inflammatory Reactions
4.3. Several Remarks for Experimental Design and Following Studies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AS | asymptomatic trout |
| IP | moderately symptomatic trout |
| SL | trout with severe lesions |
| DEPs | differential expression proteins |
| GO | gene ontology |
| IPS | integral score of ichthyopathology |
| OTU | operational taxonomic unit |
| LFQ | label-free quantification |
| PCA | principal component analysis |
| APPs | acute phase proteins |
| LECT2 | leukocyte cell-derived chemotaxin 2 |
| ITIH3 | inter-alpha-trypsin inhibitor heavy chain H3-like protein |
| V. anguillarum | Vibrio anguillarum |
| DRTP1 | differentially regulated trout protein |
| COMT | catechol-O-methyltransferase domain-containing protein |
| PEDG | pigment epithelial derived factor |
References
- The State of World Fisheries and Aquaculture 2020; FAO: Rome, Italy, 2020.
- Moiseenko, T.; Sharov, A. Large Russian Lakes Ladoga, Onega, and Imandra under Strong Pollution and in the Period of Revitalization: A Review. Geosciences 2019, 9, 492. [Google Scholar] [CrossRef] [Green Version]
- Ina-Salwany, M.Y.; Al-Saari, N.; Mohamad, A.; Mursidi, F.; Mohd-Aris, A.; Amal, M.N.A.; Kasai, H.; Mino, S.; Sawabe, T.; Zamri-Saad, M. Vibriosis in Fish: A Review on Disease Development and Prevention. J. Aquat. Anim. Health 2019, 31, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.-S.; Xin, Y.-R.; Yang, X.-D.; Zeng, C.; Zhao, H.-K.; Liu, M.-Q.; Zhang, M.-Z.; Daniel, J.G.; Li, J.-F.; Wen, H.-S. Transcriptional Profiles of Genes Related to Stress and Immune Response in Rainbow Trout (Oncorhynchus mykiss) Symptomatically or Asymptomatically Infected with Vibrio anguillarum. Front. Immunol. 2021, 12, 639489. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-N.; Zhao, Y.-T.; Cao, S.-L.; Wang, H.; Zhang, J.-J. Integrated transcriptomic and proteomic analyses of grass carp intestines after vaccination with a double-targeted DNA vaccine of Vibrio mimicus. Fish Shellfish Immunol. 2020, 98, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Karami, A.M.; Ødegård, J.; Marana, M.H.; Zuo, S.; Jaafar, R.; Mathiessen, H.; von Gersdorff Jørgensen, L.; Kania, P.W.; Dalsgaard, I.; Nielsen, T.; et al. A Major QTL for Resistance to Vibrio anguillarum in Rainbow Trout. Front. Genet. 2020, 11, 607558. [Google Scholar] [CrossRef]
- Yang, J.; Liu, B.; Li, X.; Li, G.; Wen, H.; Qi, X.; Li, Y.; He, F. Immune correlates of NF-κB and TNFα promoter DNA methylation in Japanese flounder (Paralichthys olivaceus) muscle and immune parameters change response to Vibrio anguillarum infection. Fish Shellfish Immunol. 2021, 119, 578–586. [Google Scholar] [CrossRef]
- Qi, L.; Chen, Y.; Shi, K.; Ma, H.; Wei, S.; Sha, Z. Combining of transcriptomic and proteomic data to mine immune-related genes and proteins in the liver of Cynoglossus semilaevis challenged with Vibrio anguillarum. Comp. Biochem. Physiol. Part D Genom. Proteom. 2021, 39, 100864. [Google Scholar] [CrossRef]
- Kutyrev, I.; Cleveland, B.; Leeds, T.; Wiens, G.D. Proinflammatory cytokine and cytokine receptor gene expression kinetics following challenge with Flavobacterium psychrophilum in resistant and susceptible lines of rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2016, 58, 542–553. [Google Scholar] [CrossRef] [Green Version]
- Ransom, D.P.; Lannan, C.N.; Rohovec, J.S.; Fryer, J.L. Comparison of histopathology caused by Vibrio anguillarum and Vibrio ordalii in three species of Pacific salmon*. J. Fish Dis. 1984, 7, 107–115. [Google Scholar] [CrossRef]
- Gerwick, L.; Corleysmith, G.; Bayne, C. Gene transcript changes in individual rainbow trout livers following an inflammatory stimulus. Fish Shellfish Immunol. 2007, 22, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ma, A.; Sun, Z. Genetic parameters of seven immune factors in turbot (Scophthalmus maximus) infected with Vibrio anguillarum. J. Fish Dis. 2021, 44, 263–271. [Google Scholar] [CrossRef]
- Semple, S.L.; Heath, G.; Filice, C.T.; Heath, D.D.; Dixon, B. The impact of outbreeding on the immune function and disease status of eight hybrid Chinook salmon crosses after Vibrio anguillarum challenge. Aquac. Res. 2022, 53, 957–973. [Google Scholar] [CrossRef]
- Witeska, M.; Kondera, E.; Ługowska, K.; Bojarski, B. Hematological methods in fish—Not only for beginners. Aquaculture 2022, 547, 737498. [Google Scholar] [CrossRef]
- Izergina, E.E.; Izergin, I.L.; Izergin, L.I. Atlas Kletok Krovi Lososevyx ryb Materikovogo Poberezh’ya Severnoj Chasti Oxotskogo Morya [Atlas of Blood Cells Salmon Continental Coast of the Northern Part of the Okhotsk Sea]; Kordis: Magadan, Russia, 2014; ISBN 9785896782278. [Google Scholar]
- Ivanova, N.T. Atlas of Fish Blood Cells: Comparative Morphology and Classification of the Formed Elements of Fish Blood; Legkaya and Pitshevaya Prom.: Moscow, Russia, 1983. [Google Scholar]
- Amin, A.B.; Mortensen, L.; Poppe, T.T. Histology Atlas, Normal Structure of Salmonids: A Colour Atlas, English, German, French and Spanish Legends; Akvapatologisk Laboratorium AS: Bodø, Norway, 1992; ISBN 9788299240611. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Salem, M.; Silverstein, J.; Rexroad, C.E.; Yao, J. Effect of starvation on global gene expression and proteolysis in rainbow trout (Oncorhynchus mykiss). BMC Genom. 2007, 8, 328. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.-H.; Austin, B. Cytokine expression in leucocytes and gut cells of rainbow trout, Oncorhynchus mykiss Walbaum, induced by probiotics. Vet. Immunol. Immunopathol. 2006, 114, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Tongsri, P.; Meng, K.; Liu, X.; Wu, Z.; Yin, G.; Wang, Q.; Liu, M.; Xu, Z. The predominant role of mucosal immunoglobulin IgT in the gills of rainbow trout (Oncorhynchus mykiss) after infection with Flavobacterium columnare. Fish Shellfish Immunol. 2020, 99, 654–662. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Frans, I.; Michiels, C.W.; Bossier, P.; Willems, K.A.; Lievens, B.; Rediers, H. Vibrio anguillarum as a fish pathogen: Virulence factors, diagnosis and prevention. J. Fish Dis. 2011, 34, 643–661. [Google Scholar] [CrossRef] [PubMed]
- Ceylan, M.; Duman, M.; İnan, S.; Özyiğit, Ö.; Satıcıoğlu, İ.B.; Altun, S. Listonella (Vibrio) anguillarum ile deneysel enfeksiyon yapılan gökkuşağı alabalıklarında histopatolojik değişikliklerin incelenmesi. Uludağ Üniversitesi Veteriner Fakültesi Dergisi, 2 June 2019. [Google Scholar] [CrossRef]
- Lamas, J.; Santos, Y.; Bruno, D.; Toranzo, A.E.; Anadon, R. A Comparison of Pathological Changes Caused by Vibrio anguillarum and Its Extracellular Products in Rainbow Trout (Oncorhynchus mykiss). Fish Pathol. 1994, 29, 79–89. [Google Scholar] [CrossRef] [Green Version]
- Tsai, H.-T.; Lee, T.-H.; Yang, S.-F.; Lin, L.-Y.; Tee, Y.-T.; Wang, P.-H. Markedly elevated soluble E-cadherin in plasma of patient with pelvic inflammatory disease. Fertil. Steril. 2013, 99, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Fu, Q.; Su, B.; Zhou, S.; Liu, F.; Song, L.; Zhang, M.; Ren, Y.; Dong, X.; Tan, F.; et al. Transcriptomic profiling revealed the signatures of intestinal barrier alteration and pathogen entry in turbot (Scophthalmus maximus) following Vibrio anguillarum challenge. Dev. Comp. Immunol. 2016, 65, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Freedman, S.J. The role of alpha 2-macroglobulin in furunculosis: A comparison of rainbow trout and brook trout. Comp. Biochem. Physiol. B 1991, 98, 549–553. [Google Scholar] [CrossRef]
- Ellis, A.E. Innate host defense mechanisms of fish against viruses and bacteria. Dev. Comp. Immunol. 2001, 25, 827–839. [Google Scholar] [CrossRef]
- Peng, T.C.; Liao, K.W.; Lai, H.L.; Chao, Y.F.C.; Chang, F.M.; Harn, H.-J.; Lee, R.P. The physiological changes of cumulative hemorrhagic shock in conscious rats. J. Biomed. Sci. 2006, 13, 385. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Wei, Z.; Jiang, H.; Cheng, L.; Chen, Q.; Chen, M.; Yan, J.; Sun, Z. Lactate dehydrogenase is associated with 28-day mortality in patients with sepsis: A retrospective observational study. J. Surg. Res. 2018, 228, 314–321. [Google Scholar] [CrossRef]
- Braceland, M.; Bickerdike, R.; Tinsley, J.; Cockerill, D.; Mcloughlin, M.F.; Graham, D.A.; Burchmore, R.J.; Weir, W.; Wallace, C.; Eckersall, P.D. The serum proteome of Atlantic salmon, Salmo salar, during pancreas disease (PD) following infection with salmonid alphavirus subtype 3 (SAV3). J. Proteom. 2013, 94, 423–436. [Google Scholar] [CrossRef] [Green Version]
- Gerwick, L.; Reynolds, W.S.; Bayne, C.J. A precerebellin-like protein is part of the acute phase response in rainbow trout, Oncorhynchus mykiss. Dev. Comp. Immunol. 2000, 24, 597–607. [Google Scholar] [CrossRef]
- Gerwick, L.; Steinhauer, R.; Lapatra, S.; Sandell, T.; Ortuno, J.; Hajiseyedjavadi, N.; Bayne, C.J. The acute phase response of rainbow trout (Oncorhynchus mykiss) plasma proteins to viral, bacterial and fungal inflammatory agents. Fish Shellfish Immunol. 2002, 12, 229–242. [Google Scholar] [CrossRef]
- Bakke, F.K.; Monte, M.M.; Stead, D.A.; Causey, D.R.; Douglas, A.; Macqueen, D.J.; Dooley, H. Plasma Proteome Responses in Salmonid Fish Following Immunization. Front. Immunol. 2020, 11, 581070. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.-C.; Huang, S.-C. Differential Effects of Statins on Inflammatory Interleukin-8 and Antimicrobial Peptide Human Β-Defensin 2 Responses in Salmonella-Infected Intestinal Epithelial Cells. Int. J. Mol. Sci. 2018, 19, 1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.-H.; Austin, B. Haemolysins in Vibrio species. J. Appl. Microbiol. 2005, 98, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.; McCulloh, R.J. Hemopexin and haptoglobin: Allies against heme toxicity from hemoglobin not contenders. Front. Physiol. 2015, 6, 187. [Google Scholar] [CrossRef] [PubMed]
- Yuzaki, M. Cbln and C1q family proteins—New transneuronal cytokines. Cell. Mol. Life Sci. 2008, 65, 1698–1705. [Google Scholar] [CrossRef] [PubMed]
- Drouin, M.; Saenz, J.; Chiffoleau, E. C-Type Lectin-Like Receptors: Head or Tail in Cell Death Immunity. Front. Immunol. 2020, 11, 251. [Google Scholar] [CrossRef] [Green Version]
- Kokkinos, P.A.; Kazantzi, A.; Sfyroera, G.; Zarkadis, I.K. Molecular cloning of leukocyte cell-derived chemotaxin 2 in rainbow trout. Fish Shellfish Immunol. 2005, 18, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Nakao, M.; Tsujikura, M.; Ichiki, S.; Vo, T.K.; Somamoto, T. The complement system in teleost fish: Progress of post-homolog-hunting researches. Dev. Comp. Immunol. 2011, 35, 1296–1308. [Google Scholar] [CrossRef]
- Hollebecq, M.-G.; Faivre, B.; Bourmaud, C.; Michel, C. Spontaneous bactericidal and complement activities in serum of rainbow trout (Oncorhynchus mykiss) genetically selected for resistance or susceptibility to furunculosis. Fish Shellfish Immunol. 1995, 5, 407–426. [Google Scholar] [CrossRef]
- Castro, R.; Tafalla, C. Overview of fish immunity. In Mucosal Health in Aquaculture; Elsevier: Amsterdam, The Netherlands, 2015; pp. 3–54. [Google Scholar]
- Chang, C.-I.; Zhang, Y.-A.; Zou, J.; Nie, P.; Secombes, C.J. Two Cathelicidin Genes Are Present in both Rainbow Trout (Oncorhynchus mykiss) and Atlantic Salmon (Salmo salar). Antimicrob. Agents Chemother. 2006, 50, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Wei, W.; Xu, H.; Wang, Q.; Zhang, X.; Chang, K.; Wu, C.; Zhang, Y. Identification of differentially expressed genes in large yellow croaker (Pseudosciaena crocea) induced by attenuated live Vibrio anguillarum. Aquaculture 2009, 291, 124–129. [Google Scholar] [CrossRef]
- Pridgeon, J.W.; Klesius, P.H. Apolipoprotein A1 in channel catfish: Transcriptional analysis, antimicrobial activity, and efficacy as plasmid DNA immunostimulant against Aeromonas hydrophila infection. Fish Shellfish Immunol. 2013, 35, 1129–1137. [Google Scholar] [CrossRef] [PubMed]
- Khansari, A.R.; Balasch, J.C.; Vallejos-Vidal, E.; Parra, D.; Reyes-López, F.E.; Tort, L. Comparative Immune- and Stress-Related Transcript Response Induced by Air Exposure and Vibrio anguillarum Bacterin in Rainbow Trout (Oncorhynchus mykiss) and Gilthead Seabream (Sparus aurata) Mucosal Surfaces. Front. Immunol. 2018, 9, 856. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurpe, S.R.; Sukhovskaya, I.V.; Borvinskaya, E.V.; Morozov, A.A.; Parshukov, A.N.; Malysheva, I.E.; Vasileva, A.V.; Chechkova, N.A.; Kuchko, T.Y. Physiological and Biochemical Characteristics of Rainbow Trout with Severe, Moderate and Asymptomatic Course of Vibrio anguillarum Infection. Animals 2022, 12, 2642. https://doi.org/10.3390/ani12192642
Kurpe SR, Sukhovskaya IV, Borvinskaya EV, Morozov AA, Parshukov AN, Malysheva IE, Vasileva AV, Chechkova NA, Kuchko TY. Physiological and Biochemical Characteristics of Rainbow Trout with Severe, Moderate and Asymptomatic Course of Vibrio anguillarum Infection. Animals. 2022; 12(19):2642. https://doi.org/10.3390/ani12192642
Chicago/Turabian StyleKurpe, Stanislav Rimaso, Irina Viktorovna Sukhovskaya, Ekaterina Vitalyevna Borvinskaya, Alexey Anatolievich Morozov, Aleksey Nikolaevich Parshukov, Irina Evgenyevna Malysheva, Alina Valeryevna Vasileva, Natalia Alexandrovna Chechkova, and Tamara Yurevna Kuchko. 2022. "Physiological and Biochemical Characteristics of Rainbow Trout with Severe, Moderate and Asymptomatic Course of Vibrio anguillarum Infection" Animals 12, no. 19: 2642. https://doi.org/10.3390/ani12192642