Assessment of the Impact on 20 Pelagic Fish Species by the Taiwanese Small-Scale Longline Fishery in the Western North Pacific Using Ecological Risk Assessment


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
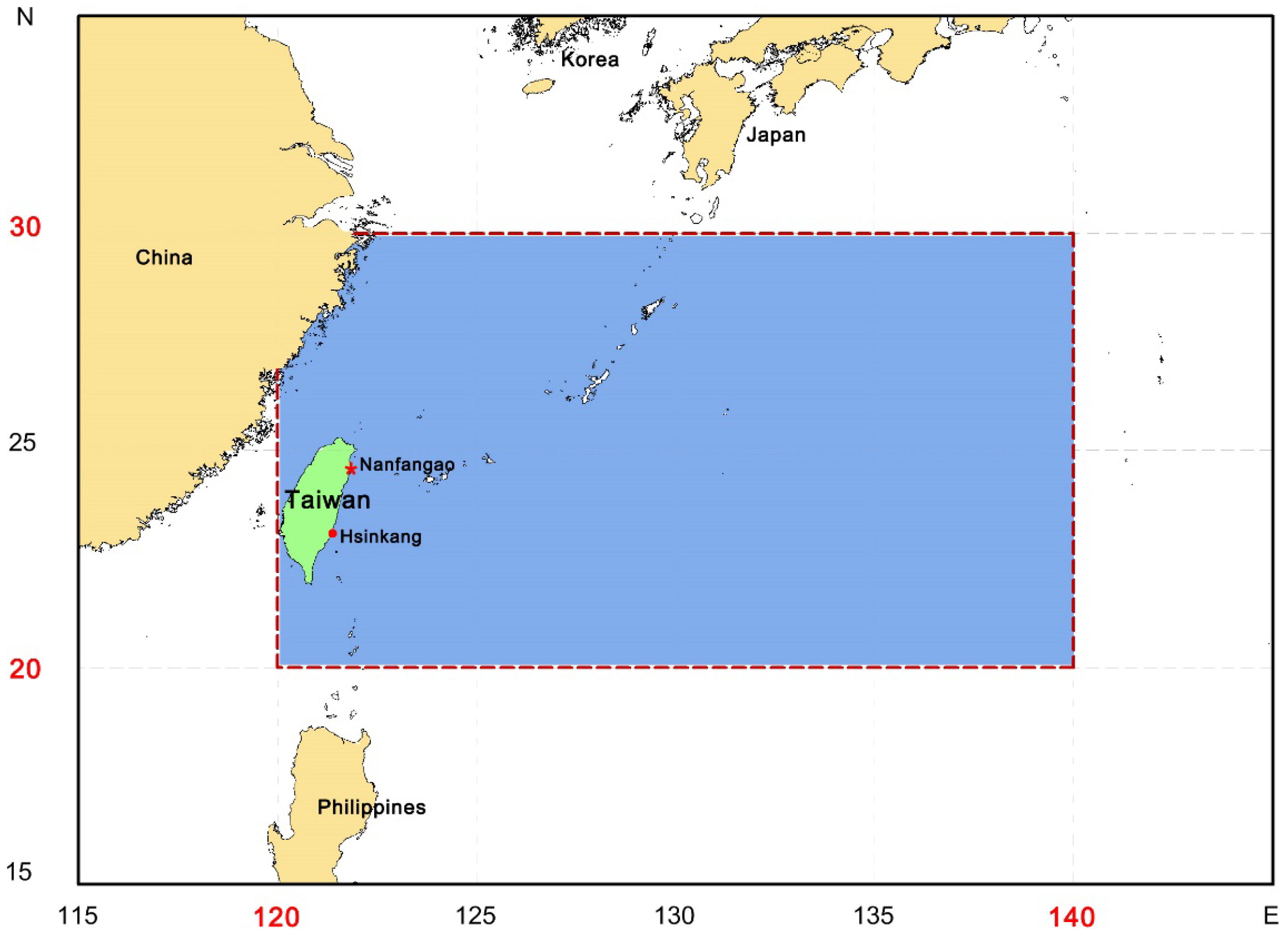
2.1. Study Area and Species
2.2. Source of Data
2.3. ERA
3. Results
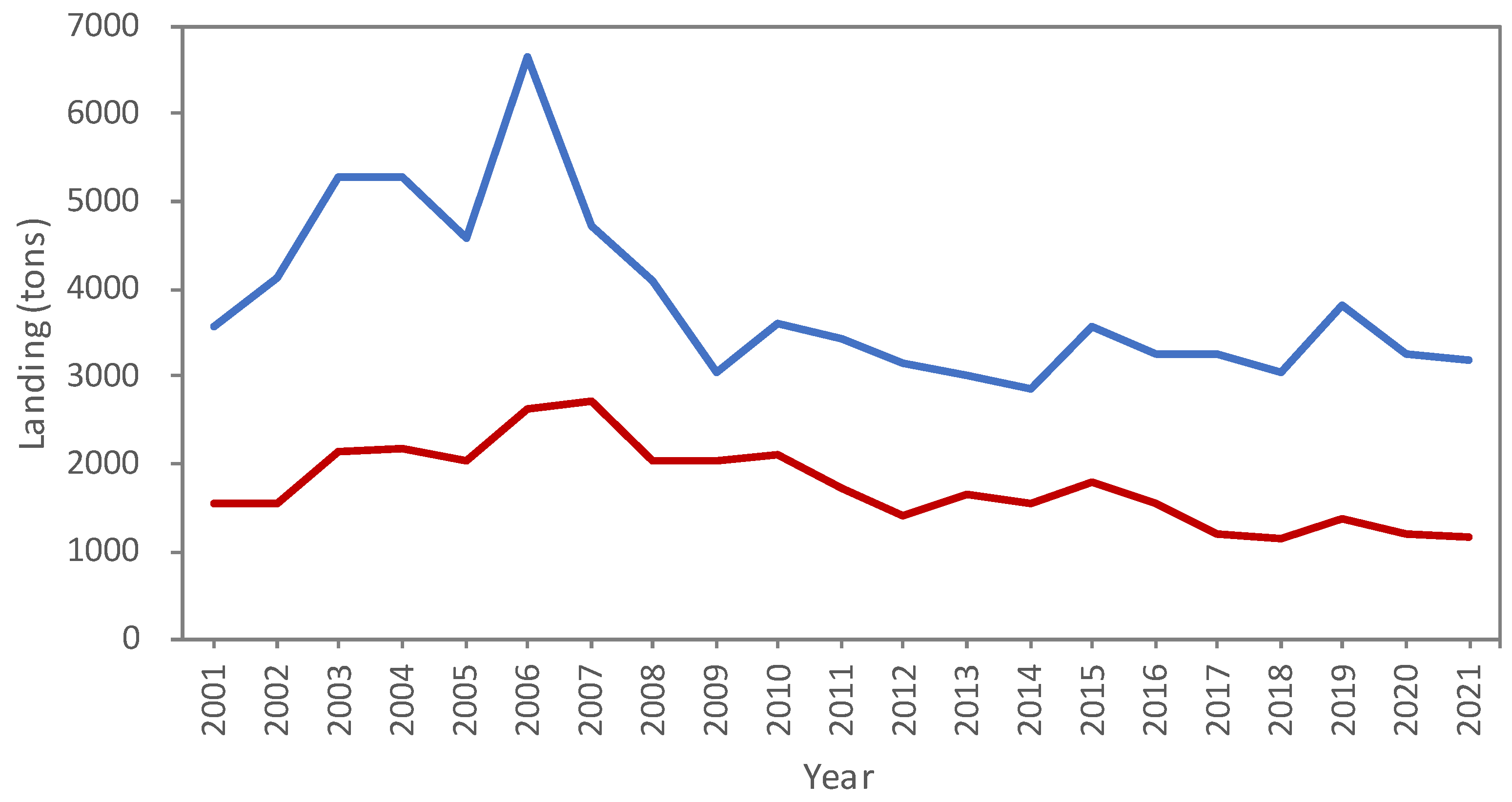
3.1. Sales Data Analysis
3.2. Life History Parameters and Biological Characteristics
3.2.1. Length
3.2.2. Age
- Maximum age (Amax): The largest Amax of sharks was 50 years for the dusky shark, the smallest Amax was 11.6 years for the scalloped hammerhead shark; the largest Amax of tunas was 26 years for the Pacific bluefin tuna, and the smallest Amax was 7.7 years for yellowfin tuna; the largest Amax of billfishes was 14 years for the blue marlin, and the smallest Amax was 6 years for striped marlin (Supplemental Table S1).
- Age at maturity (Am): The largest Am of sharks was 16.4 years for the dusky shark, the smallest Am was 4.7 years for the scalloped hammerhead shark; the largest Am of tunas was 8 years for the Pacific bluefin tuna, and the smallest Am was 2.4 years for yellowfin tuna; the largest Am of billfishes was 7.4 years for the blue marlin, and the smallest Amax was 4.8 years for striped marlin (Supplemental Tables S2 and S3).
3.2.3. Other Life History Parameters
- Fecundity/litter size: The largest litter size for sharks was the scalloped hammerhead of 30, and smallest was two for bigeye and pelagic thresher. The largest batch fecundity of tunas was the Pacific bluefin tuna of 5.8–25.2 million eggs, and the smallest was albacore of 0.94 million eggs; the largest of billfishes was the blue marlin of 6.94 million and the smallest was sailfish of 1.30 million (Supplemental Table S3).
- Reproduction mode: Carcharhinidae species, blue shark, and hammerhead sharks are viviparity, bigeye and pelagic thresher, and shortfin mako sharks are aplacental viviparity. Tunas and billfishes are oviparity.
- Trophic position (Tp): The Tp derived from the Ecopath [34] were as follows: 4.16 for Carcharhindae, 3.99 for blue shark, and blue marlin, 3.94 for swordfish, 3.89 for other billfishes, 3.78 for yellowfin tuna, and bigeye tuna, 3.75 for other tunas. The Tp of all the species was greater than 3.0, which was in high trophic position.
3.3. ERA
4. Discussion
4.1. Catch Data
4.2. Life History Parameters
4.3. ERA
4.4. Comparison with Single-Species Approach
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, K.M.; Huang, R.H.; Su, K.Y.; Joung, S.J. Vulnerability assessment of pelagic sharks in the western North Pacific by using an integrated ecological risk assessment. Animals 2021, 11, 2161. [Google Scholar] [CrossRef] [PubMed]
- Latour, R.J.; Brush, M.J.; Bonzek, C.F. Toward ecosystem-based fisheries management: Strategies for multispecies modeling and associated data requirements. Fisheries 2003, 28, 10–22. [Google Scholar] [CrossRef]
- Liu, K.M.; Chen, C.T. Demographic analysis of the scalloped hammerhead, Sphyrna lewini, in the northwestern Pacific. Fish Sci. 1999, 65, 218–223. [Google Scholar] [CrossRef]
- Liu, K.M.; Chang, Y.T.; Ni, I.H.; Jin, C.B. Spawning per recruit analysis of the pelagic thresher shark, Alopias pelagicus, in northeastern Taiwan waters. Fish. Res. 2006, 82, 56–64. [Google Scholar] [CrossRef]
- Tsai, W.P.; Liu, K.M.; Joung, S.J. Demographic analysis of the pelagic thresher shark, Alopias pelagicus in Northwest Pacific using a stochastic stage-based model. Mar. Freshw. Res. 2010, 61, 1056–1066. [Google Scholar] [CrossRef]
- Chang, J.H.; Liu, K.M. Stock assessment of the shortfin mako shark, Isurus oxyrinchus, in the Northwest Pacific Ocean using per-recruit and virtual population analyses. Fish. Res. 2009, 98, 92–103. [Google Scholar] [CrossRef]
- Tsai, W.P.; Sun, C.L.; Wang, S.P.; Liu, K.M. Evaluating the impacts of uncertainty on the estimation of biological reference points for the shortfin mako shark, Isurus oxyrinchus, in the north-western Pacific Ocean. Mar. Freshwater Res. 2011, 62, 1383–1394. [Google Scholar] [CrossRef]
- Tsai, W.P.; Sun, C.L.; Punt, A.E.; Liu, K.M. Demographic analysis of the shortfin mako shark, Isurus oxyrinchus, in the Northwestern Pacific using a two-sex stage-based matrix model. ICES J. Mar. Sci. 2014, 71, 1604–1618. [Google Scholar] [CrossRef]
- Tsai, W.P.; Liu, K.M.; Punt, A.E.; Sun, C.L. Assessing the potential biases of ignoring sexual dimorphism and mating mechanism in using a single-sex demographic model: The shortfin mako shark as a case study. ICES J. Mar. Sci. 2015, 72, 793–803. [Google Scholar] [CrossRef]
- Tsai, W.P.; Wu, J.R.; Yan, M.Z.; Liu, K.M. Assessment of biological reference points for management of the smooth hammerhead shark, Sphyrna zygaena, in the Northwest Pacific Ocean. J. Fish. Soc. Taiwan 2018, 45, 29–41. [Google Scholar] [CrossRef]
- Tsai, W.P.; Chang, Y.J.; Liu, K.M. Development and testing of a Bayesian population model for the bigeye thresher shark, Alopias superciliosus, in an area subset of the western North Pacific. Fish. Manage. Ecol. 2019, 26, 269–294. [Google Scholar] [CrossRef]
- Tsai, W.P.; Liu, K.M.; Chang, Y.J. Evaluation of biological reference points for conservation and management of the bigeye thresher shark, Alopias superciliosus, in the Northwest Pacific. Sustainability 2020, 12, 8646. [Google Scholar] [CrossRef]
- Liu, P.Y. Age, Growth, Mortality and Per-Recruit Analyses Based on Length Data for the Bigeye Tuna Thunnus obesus of Taiwanese Offshore Tuna Longline Fishery. Master’s Thesis, National Taiwan University, Taipei, Taiwan, 2005; p. 86. [Google Scholar]
- Chung, F.J. Stock assessment of Pacific bluefin tuna (Thunnus orientalis) by non-equilibrium global production model analysis. Master’s Thesis, National Taiwan University, Taipei, Taiwan, 2008; p. 32. [Google Scholar]
- Chen, K.S. Life History Traits and Yield-Per-Recruit Analysis of North Pacific Albacore (Thunnus alalunga). Ph.D. Thesis, National Taiwan University, Taipei, Taiwan, 2011; p. 166. [Google Scholar]
- Chiang, W.C. Population Dynamics and Stock Assessment of Sailfish (Istiophorus platypterus) in Waters off Eastern Taiwan. Ph.D. Thesis, National Taiwan University, Taipei, Taiwan, 2004; p. 195. [Google Scholar]
- Wang, S.P.; Sun, C.L.; Punt, A.E.; Yeh, S.Z. Application of the sex-specific age-structured assessment method for swordfish, Xiphias gladius, in the North Pacific Ocean. Fish. Res. 2007, 84, 282–300. [Google Scholar] [CrossRef]
- Fang, H.P. Estimation of Life History Parameters, Biological Reference Points, and Associated Uncertainties for Black Marlin (Makaira indica) in the Waters off Eastern Taiwan. Master’s Thesis, National Taiwan University, Taipei, Taiwan, 2008; p. 87. [Google Scholar]
- Shih, N.T. Applying Life History Traits in Providing Management Scenarios for Taiwanese Coastal and Offshore Fishery Resources. Ph.D. Thesis, National Taiwan Ocean University, Taipei, Taiwan, 2009; p. 116. [Google Scholar]
- Liu, K.M.; Chin, C.P.; Chen, C.H.; Chang, J.H. Estimating finite rate of population increase for sharks based on vital parameters. PLoS ONE 2015, 10, e0143008. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.M.; Huang, Y.W.; Hsu, H.H. Management implications for skates and rays based on analysis of life history parameters. Front. Mar. Sci. 2021, 8, 664611. [Google Scholar] [CrossRef]
- Scandol, J.; Ives, A.R.; Lockett, M.M. Development of National Guidelines to Improve the Application of Risk-Based Methods in the Scope, Implementation and Interpretation of Stock Assessments for Data-Poor Species; FRDC Project No. 2007/016; Canberra, Australia, 2009. [Google Scholar]
- Ormseth, O.A.; Spencer, P.D. An assessment of vulnerability in Alaska groundfish. Fish. Res. 2011, 112, 127–133. [Google Scholar] [CrossRef]
- Arrizabalaga, H.; de Bruyn, P.; Diaz, G.A.; Murua, H.; Chavance, P.; de Molina, A.D.; Gaertner, D.; Ariz, J.; Ruiz, J.; Kell, L.T. Productivity and susceptibility analysis for species caught in Atlantic tuna fisheries. Aquat. Living Resour. 2011, 24, 1–12. [Google Scholar] [CrossRef]
- Griffiths, S.P.; Brewer, D.T.; Heales, D.S.; Milton, D.A.; Stobutzki, I.C. Validating ecological risk assessments for fisheries: Assessing the impacts of turtle excluder devices on elasmobranch bycatch populations in an Australian trawl fishery. Mar. Freshwater Res. 2006, 57, 395–401. [Google Scholar] [CrossRef]
- Kirby, D.S. Ecological Risk Assessment for Species Caught in WCPO Tuna Fisheries: Inherent Risk as Determined by Productivity-Susceptibility Analysis; WCPFC–SC2–2006/EB WP–1; Western and Central Pacific Fisheries Commission: Kolonia, Micronesia, 2006; p. 25. [Google Scholar]
- Murua, H.; Coelho, R.; Santos, M.N.; Arrizabalaga, H.; Yokawa, K.; Romanov, E.; Zhu, J.F.; Kim, Z.G.; Bach, P.; Chavance, P.; et al. Preliminary Ecological Risk Assessment (ERA) for Shark Species Caught in Fisheries Managed by the Indian Ocean Tuna Commission (IOTC); IOTC-2012-WPEB-31; Indian Ocean Tuna Commission (IOTC): Victoria, Seychelles, 2012; p. 16. [Google Scholar]
- Murua, H.; Santiago, J.; Coelho, R.; Zudaire, I.; Neves, C.; Rosa, D.; Zudaire, I.; Semba, Y.; Geng, Z.; Bach, P.; et al. Updated Ecological Risk Assessment (ERA) for Shark Species Caught in Fisheries Managed by the Indian Ocean Tuna Commission (IOTC); IOTC-2018-SC21-14_Rev1; Indian Ocean Tuna Commission (IOTC): Victoria, Seychelles, 2018; p. 28. [Google Scholar]
- Simpfendorfer, C.A.; Cortés, E.; Heupel, M.; Brooks, E.; Babcock, E.; Baum, J.; McAuley, R.; Dudley, S.; Stevens, J.D.; Fordham, S.; et al. An Integrated Approach to Determining the Risk of Over-Exploitation for Data-Poor Pelagic Atlantic Sharks; ICCAT SCRS/2008/140; International Commission for the Conservation of Atlantic Tunas (ICCAT): Madrid, Spain, 2008; p. 15. [Google Scholar]
- Cortés, E.; Arocha, F.; Beerkircher, L.; Carvalho, F.; Domingo, A.; Heupel, M.; Holtzhausen, H.; Santos, M.N.; Ribera, M.; Simpfendorfer, C. Ecological risk assessment of pelagic sharks caught in Atlantic pelagic longline fisheries. Aquat. Living Resour. 2010, 23, 25–34. [Google Scholar] [CrossRef]
- Lin, C.Y.; Wang, S.P.; Chiang, W.C.; Griffiths, S.; Yeh, H.M. Ecological risk assessment of species impacted by fisheries in waters off eastern Taiwan. Fish. Manag. Ecol. 2020, 27, 345–356. [Google Scholar] [CrossRef]
- Hobday, A.J.; Smith, A.D.M.; Stobutzki, I.C.; Bulman, C.; Daley, R.; Dambacher, J.M.; Deng, R.A.; Dowdney, J.; Fuller, M.; Furlania, D.; et al. Ecological risk assessment for the effects of fishing. Fish. Res. 2011, 108, 372–384. [Google Scholar] [CrossRef]
- Webb, H.; Hobday, A.; Dowdney, J.; Bulman, C.; Sporcic, M.; Smith, T.; Furlani, D.; Fuller, M.; Williams, A.; Stobustzki, I. Ecological Risk Assessment for Effects of Fishing. Report for Eastern Tuna & Billfish Fishery: Longline Sub-Fishery; Australia Government Australia Fisheries Management Authority: Canberra, Australia, 2007; p. 230. [Google Scholar]
- Chin, C.P. Simulation and Analysis of the Pelagic Ecosystem of the Northwest Pacific Based on Ecopath with Ecosim. Master’s Thesis, National Taiwan Ocean University, Keelung, Taiwan, 2007; p. 127. [Google Scholar]
- Murua, H.; Arrizabalaga, H.; Huang, J.H.W.; Romanov, E.; Bach, P.; de Bruyn, P.; Chavance, P.; de Molina, A.D.; Pianet, R.; Ariz, J.; et al. Ecological Risk Assessment (ERA) for Species Caught in Fisheries Managed by the Indian Ocean Tuna Commission (IOTC): A First Attempt; Indian Ocean Tuna Commission (IOTC): Victoria, Seychelles, 2009; p. 11. [Google Scholar]
- Cortés, E.; Domingo, A.; Miller, P.; Forselledo, R.; Mas, F.; Arocha, F.; Campana, S.; Coelho, R.; Da Silva, C.; Hazin, F.H.V.; et al. Expanded Ecological Risk Assessment of Pelagic Sharks Caught in Atlantic Pelagic Longline Fisheries; ICCAT SCRS/2012/167; International Commission for the Conservation of Atlantic Tunas (ICCAT): Madrid, Spain, 2012; p. 56. [Google Scholar]
- Su, N.J. Age, Growth, Mortality, and Yield-Per-Recruit of Yellowfin Tuna Caught by the Taiwanese Offshore Tuna Longline Fishery. Master’s Thesis, National Taiwan University, Taipei, Taiwan, 2003; p. 74. [Google Scholar]
- Wang, S.P. Age and Growth Study of Swordfish, Xiphias gladius, in the Waters of Taiwan. Master’s Thesis, National Taiwan University, Taipei, Taiwan, 1999; p. 97. [Google Scholar]
- Chen, B.C. Age and Growth of the Blue Marlin, Makaira mazara, in the Western Pacific Ocean. Master’s Thesis, National Taiwan University, Taipei, Taiwan, 2001; p. 76. [Google Scholar]
- Liu, C.H. Age and Growth of Black Marlin, Makaira indica, in the Waters off Eastern Taiwan. Master’s Thesis, National Taiwan University, Taipei, Taiwan, 2006; p. 94. [Google Scholar]
- Hsu, W.S. Age and Growth of Striped Marlin (Kajikia audax) in Waters off Taiwan. Master’s Thesis, National Taiwan University, Taipei, Taiwan, 2010; p. 104. [Google Scholar]
- Shimose, T.; Tanabe, T.; Chen, K.S.; Hsu, C.C. Age Determination and Growth of Pacific Bluefin Tuna, Thunnus orientalis, off Japan and Taiwan. Fish. Res. 2009, 100, 134–139. [Google Scholar] [CrossRef]
- Campana, S.E. Accuracy, precision, and quality control in age determination, including a review of the use and abuse of age validation methods. J. Fish Biol. 2001, 59, 197–242. [Google Scholar] [CrossRef]
- Chen, C.T.; Leu, T.C.; Joung, S.J.; Lo, N.C.H. Age and growth of the scalloped hammerhead, Sphyrna lewini, in northeastern Taiwan waters. Pac. Sci. 1990, 44, 156–170. [Google Scholar]
- Anislado-Tolentino, V.; Robinson-Mendoza, C. Age and growth for the scalloped hammerhead shark, Sphyrna lewini (Griffith and Smith, 1834) along the central Pacific coast of Mexico. Cienc. Mar. 2001, 27, 501–520. [Google Scholar] [CrossRef]
- Klimley, A.P. The determinants of sexual segregation in the scalloped hammerhead shark, Sphyrna lewini. Environ. Biol. Fish. 1987, 18, 27–40. [Google Scholar] [CrossRef]
- White, W.T.; Bartron, C.; Potter, I.C. Catch composition and reproductive biology of Sphyrna lewini (Griffith & Smith) (Carcharhiniformes, Sphyrnidae) in Indonesian waters. J. Fish Biol. 2008, 72, 1675–1689. [Google Scholar]
- Harry, A.V.; Macbeth, W.G.; Gutteridge, A.N.; Simpfendorfer, C.A. The life histories of endangered hammerhead sharks (Carcharhiniformes, Sphyrnidae) from the east coast of Australia. J. Fish Biol. 2011, 78, 2026–2051. [Google Scholar] [CrossRef]
- Piercy, A.N.; Carlson, J.K.; Sulikowski, J.A.; Burgess, G.H. Age and growth of the scalloped hammerhead shark, Sphyrna lewini, in the north-west Atlantic Ocean and Gulf of Mexico. Mar. Freshw. Res. 2007, 58, 34–40. [Google Scholar] [CrossRef]
- Joung, S.J.; Chen, N.F.; Hsu, H.H.; Liu, K.M. Estimates of life history parameters of the oceanic whitetip shark, Carcharhinus longimanus, in the western North Pacific Ocean. Mar. Biol. Res. 2016, 12, 758–768. [Google Scholar] [CrossRef]
- Patrick, W.S.; Spencer, P.; Ormseth, O.; Cope, J.; Field, J.; Kobayashi, D.; Gedamke, T.; Cortés, E.; Bigelow, K.; Overholtz, W.; et al. Use of Productivity and Susceptibility Indices to Determine Stock Vulnerability, with Example Applications to Six U.S. Fisheries; NOAA Technical Memorandum NMFS-F/SPO-101; National Oceanic and Atmospheric Administration (NOAA): Washington, DC, USA, 2009; p. 104. [Google Scholar]
- Patrick, W.S.; Lawson, P.; Spencer, P.; Gedamke, T.; Link, J.; Cortés, E.; Cope, J.; Ormseth, O.; Field, J.; Bigelow, K.; et al. Using productivity and susceptibility indices to assess the vulnerability of United States fish stocks to overfishing. Fish. Bull. 2010, 108, 305–322. [Google Scholar]
- Ward, P. Empirical estimates of historical variations in the catchability and fishing power of pelagic longline gear. Rev. Fish. Biol. Fish. 2008, 18, 409–426. [Google Scholar] [CrossRef]
- Liu, K.M.; Chen, C.T.; Liao, T.H.; Joung, S.J. Age, growth, and reproduction of the pelagic thresher shark, Alopias pelagicus, in the northwestern Pacific. Copeia 1999, 1999, 68–74. [Google Scholar] [CrossRef]
- Liu, K.M.; Chiang, P.J.; Chen, C.T. Age and growth estimates of the bigeye thresher, Alopias superciliosus, in northwestern Taiwan waters. Fish. Bull. 1998, 96, 262–271. [Google Scholar]
- Joung, S.J.; Liao, Y.Y.; Liu, K.M.; Chen, C.T.; Leu, L.C. Age, growth, and reproduction of the spinner shark, Carcharhinus brevipinna, in the northeastern waters of Taiwan. Zool. Stud. 2005, 44, 102–110. [Google Scholar]
- Joung, S.J.; Chen, C.T.; Lee, H.H.; Liu, K.M. Age, growth, and reproduction of the silky sharks Carcharhinus falciformis in northeastern Taiwan waters. Fish. Res. 2008, 90, 78–85. [Google Scholar] [CrossRef]
- Joung, S.J.; Chen, J.H.; Chin, C.P.; Liu, K.M. Age and growth of the dusky shark, Carcharhinus obscurus, in the Northwest Pacific Ocean. Terr. Atmos. Oceanic Sci. 2015, 26, 153–160. [Google Scholar] [CrossRef]
- Joung, S.J.; Liao, Y.Y.; Chen, C.T. Age and growth of sandbar shark, Carcharhinus plumbeus, in northeastern Taiwan waters. Fish. Res. 2004, 70, 83–96. [Google Scholar] [CrossRef]
- Chou, Y.T. Studies on Age and Growth of Smooth Hammerhead, Sphyrna zygaena in Northeastern Taiwan Waters. Master’s Thesis, National Taiwan Ocean University, Keelung, Taiwan, 2004; p. 66. [Google Scholar]
- Huang, C.C. Age and Growth of the Blue Shark, Prionace glauca, in the Northwestern Pacific Ocean. Master’s Thesis, National Taiwan Ocean University, Keelung, Taiwan, 2006; p. 76. [Google Scholar]
- Chen, C.T.; Liu, K.M.; Chang, Y.C. Reproductive biology of the bigeye thresher shark, Alopias superciliosus (Lowe, 1839) (Chondrichthyes: Alopiidae) in the northwestern Pacific. Ichthyol. Res. 1997, 44, 227–235. [Google Scholar] [CrossRef]
- Joung, S.J.; Chen, C.T. Reproduction in the sandbar shark, Carcharhinus plumbeus, in the waters off northeastern Taiwan. Copeia 1995, 1995, 659–665. [Google Scholar] [CrossRef]
- Chen, C.T.; Leu, T.C.; Joung, S.J. Notes on reproduction in the scalloped hammerhead, Sphyrna lewini, in northeastern Taiwan waters. Fish. Bull. 1988, 86, 389–393. [Google Scholar]
- Liu, S.C. Reproductive Biology of Smooth Hammerhead, Sphyrna zygaena in Northeastern Taiwan Waters. Master’s Thesis, National Taiwan Ocean University, Keelung, Taiwan, 2002; p. 84. [Google Scholar]
- Wu, T.Y. Reproductive Biology of Blue Shark, Prionace glauca in the Northwestern Pacific Ocean. Master’s Thesis, National Taiwan Ocean University, Keelung, Taiwan, 2003; p. 109. [Google Scholar]
- Sun, C.L.; Chang, Y.J.; Tszeng, C.C.; Yeh, S.Z.; Su, N.J. Reproductive biology of blue marlin (Makaira nigricans) in the western Pacific Ocean. Fish. Bull. 2009, 107, 420–432. [Google Scholar]
- Liu, T.Y. Reproductive Biology of Black Marlin, Makaira indica, in the Waters off Taiwan and South China Sea. Master’s Thesis, National Taiwan University, Taipei, Taiwan, 2007; p. 117. [Google Scholar]
- Wang, S.P.; Sun, C.L.; Yeh, S.Z. Sex Ratios and Sexual Maturity of Swordfish (Xiphias gladius L.) in the Waters of Taiwan. Zool. Stud. 2003, 42, 529–539. [Google Scholar]
- Chang, H.Y. Reproductive Biology of Striped Marlin, Kajikia audax, in the Waters off Taiwan. Master’s Thesis, National Taiwan University, Taipei, Taiwan, 2011; p. 108. [Google Scholar]
- Chen, K.S.; Crone, P.R.; Hsu, C.C. Reproductive biology of albacore Thunnus alalunga. J. Fish. Biol. 2010, 77, 119–136. [Google Scholar] [CrossRef]
- Wang, W.J. Reproductive Biology of Yellowfin Tuna Thunnus albacares in the Western Pacific Ocean. Master’s Thesis, National Taiwan University, Taipei, Taiwan, 2005; p. 146. [Google Scholar]
- Ju, S.L. Reproductive Biology of Bigeye Tuna Thunnus obesus in the Western Pacific Ocean. Master’s Thesis, National Taiwan University, Taipei, Taiwan, 1998; p. 67. [Google Scholar]
- Chen, K.S.; Crone, P.R.; Hsu, C.C. Reproductive biology of female Pacific bluefin tuna Thunnus orientalis from south-western North Pacific Ocean. Fish. Sci. 2006, 72, 985–994. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Common Name | Species Code | IUCN Status |
|---|---|---|---|
| Alopias pelagicus | Pelagic thresher shark | PTH | EN |
| A. superciliosus | Bigeye thresher shark | BTH | VU |
| Carcharhinus brevipinna | Spinner shark | CCB | NT |
| C. falciformis | Silky shark | FAL | VU |
| C. longimanus | Oceanic whitetip shark | OCS | CR |
| C. obscurus | Dusky shark | DUS | EN |
| C. plumbeus | Sandbar shark | CCP | VU |
| Isurus oxyrinchus | Shortfin mako | SMA | EN |
| Sphyrna lewini | Scalloped hammerhead | SPL | CR |
| S. zygaena | Smooth hammerhead | SPZ | VU |
| Prionace glauca | Blue shark | BSH | NT |
| Makaira nigricans | Blue marlin | BUM | VU |
| Istiompax indica | Black marlin | BLM | DD |
| Istiophorus platypterus | Sailfish | SAF | VU |
| Xiphias gladius | Swordfish | SWO | NT |
| Kajikia audax | Striped marlin | STM | LC |
| Thunnus alalunga | Albacore tuna | ALB | LC |
| T. albacares | Yellowfin tuna | YFT | LC |
| T. obesus | Bigeye tuna | BET | VU |
| T. orientalis | Pacific bluefin tuna | BFT | NT |
| Attribute | Low Productivity High Risk (R = 1) | Medium Productivity Median Risk (R = 2) | High Productivity Low Risk (R = 3) |
|---|---|---|---|
| Age at maturity (Am) | >15 years | 5–15 years | <5 years |
| Length at maturity (Lm) | >200 cm | 40–200 cm | <40 cm |
| Maximum age (Amax) | >25 years | 10–25 years | <10 years |
| Maximum length (Lmax) | >300 cm | 100–300 cm | <100 cm |
| Fecundity (F) | <100 | 100–20,000 | >20,000 |
| Reproductive strategy (R) | viviparity or aplacental viviparity | with parental care after spawning | without parental care after spawning |
| Trophic position (TP) | >3.25 | 2.75–3.25 | <2.75 |
| Species | Rank | |||||||
|---|---|---|---|---|---|---|---|---|
| Code | Am | Lm | Amax | Lmax | F | R | TP | P |
| PTH | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1.14 |
| BTH | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1.14 |
| CCB | 2 | 1 | 2 | 2 | 1 | 1 | 1 | 1.43 |
| FAL | 2 | 1 | 1 | 2 | 1 | 1 | 1 | 1.29 |
| OCS | 2 | 2 | 2 | 2 | 1 | 1 | 1 | 1.57 |
| DUS | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1.00 |
| CCP | 2 | 2 | 2 | 2 | 1 | 1 | 1 | 1.57 |
| SMA | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1.00 |
| SPL | 3 | 1 | 2 | 1 | 1 | 1 | 1 | 1.43 |
| SPZ | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1.14 |
| BSH | 3 | 2 | 2 | 1 | 1 | 1 | 1 | 1.57 |
| BUM | 2 | 2 | 2 | 2 | 3 | 3 | 1 | 2.14 |
| BLM | 2 | 2 | 2 | 1 | 3 | 3 | 1 | 2.00 |
| SAF | 2 | 2 | 2 | 2 | 3 | 3 | 1 | 2.14 |
| SWO | 2 | 2 | 2 | 2 | 3 | 3 | 1 | 2.14 |
| STM | 3 | 2 | 3 | 2 | 3 | 3 | 1 | 2.43 |
| ALB | 3 | 2 | 2 | 2 | 3 | 3 | 1 | 2.29 |
| YFT | 3 | 2 | 3 | 2 | 3 | 3 | 1 | 2.43 |
| BET | 3 | 2 | 2 | 2 | 3 | 3 | 1 | 2.29 |
| BFT | 2 | 2 | 2 | 2 | 3 | 3 | 1 | 2.14 |
| Species Code | Catchability | Selectivity | Post-Capture Mortality | S |
|---|---|---|---|---|
| PTH | 0.0241 | 1.0000 | 0.7800 | 0.0188 |
| BTH | 0.0404 | 1.0000 | 0.7800 | 0.0315 |
| CCB | 0.0180 | 0.9970 | 0.7700 | 0.0138 |
| FAL | 0.0095 | 0.9960 | 0.8600 | 0.0082 |
| OCS | 0.0013 | 0.9350 | 0.7700 | 0.0009 |
| DUS | 0.0163 | 1.0000 | 0.9200 | 0.0150 |
| CCP | 0.0113 | 0.9480 | 0.8600 | 0.0092 |
| SMA | 0.0666 | 1.0000 | 0.9200 | 0.0613 |
| SPL | 0.0461 | 0.9680 | 0.8300 | 0.0370 |
| SPZ | 0.0133 | 0.9490 | 0.8500 | 0.0107 |
| BSH | 0.1702 | 1.0000 | 0.7900 | 0.1344 |
| BUM | 0.0611 | 0.9210 | 1.0000 | 0.0563 |
| BLM | 0.0429 | 0.9730 | 1.0000 | 0.0417 |
| SAF | 0.1368 | 0.7700 | 1.0000 | 0.1053 |
| SWF | 0.0502 | 0.9210 | 1.0000 | 0.0462 |
| STM | 0.0158 | 0.3830 | 1.0000 | 0.0061 |
| ALB | 0.0231 | 0.6460 | 1.0000 | 0.0149 |
| YFT | 0.2089 | 0.8920 | 1.0000 | 0.1864 |
| BET | 0.0352 | 0.9700 | 1.0000 | 0.0342 |
| BFT | 0.0089 | 0.6080 | 1.0000 | 0.0054 |
| Species Code | P | S | ERA | Risk Ranking |
|---|---|---|---|---|
| PTH | 1.1429 | 0.0188 | 0.9096 | 6 |
| BTH | 1.1429 | 0.0315 | 0.9184 | 5 |
| CCB | 1.4286 | 0.0138 | 0.7176 | 10 |
| FAL | 1.2857 | 0.0082 | 0.8106 | 8 |
| OCS | 1.5714 | 0.0009 | 0.6191 | 12 |
| DUS | 1.0000 | 0.0150 | 1.0028 | 2 |
| CCP | 1.5714 | 0.0092 | 0.6207 | 11 |
| SMA | 1.0000 | 0.0613 | 1.0459 | 1 |
| SPL | 1.4286 | 0.0370 | 0.7379 | 9 |
| SPZ | 1.1429 | 0.0107 | 0.9064 | 7 |
| BSH | 1.5714 | 0.1344 | 0.9138 | 4 |
| BUM | 2.1429 | 0.0563 | 0.3686 | 15 |
| BLM | 2.0000 | 0.0417 | 0.3932 | 14 |
| SAF | 2.1429 | 0.1053 | 0.5779 | 13 |
| SWO | 2.1429 | 0.0462 | 0.3317 | 16 |
| STM | 2.4286 | 0.0061 | 0.0564 | 20 |
| ALB | 2.2857 | 0.0149 | 0.1612 | 19 |
| YFT | 2.4286 | 0.1864 | 0.9330 | 3 |
| BET | 2.2857 | 0.0342 | 0.2228 | 18 |
| BFT | 2.1429 | 0.0054 | 0.2396 | 17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, K.-M.; Huang, L.-H.; Su, K.-Y. Assessment of the Impact on 20 Pelagic Fish Species by the Taiwanese Small-Scale Longline Fishery in the Western North Pacific Using Ecological Risk Assessment. Animals 2022, 12, 2124. https://doi.org/10.3390/ani12162124
Liu K-M, Huang L-H, Su K-Y. Assessment of the Impact on 20 Pelagic Fish Species by the Taiwanese Small-Scale Longline Fishery in the Western North Pacific Using Ecological Risk Assessment. Animals. 2022; 12(16):2124. https://doi.org/10.3390/ani12162124
Chicago/Turabian StyleLiu, Kwang-Ming, Lung-Hsin Huang, and Kuan-Yu Su. 2022. "Assessment of the Impact on 20 Pelagic Fish Species by the Taiwanese Small-Scale Longline Fishery in the Western North Pacific Using Ecological Risk Assessment" Animals 12, no. 16: 2124. https://doi.org/10.3390/ani12162124