
Modulatory Impacts of Multi-Strain Probiotics on Rabbits’ Growth, Nutrient Transporters, Tight Junctions and Immune System to Fight against Listeria monocytogenes Infection

 ,
,  , , , ,
, , , ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Probiotics Preparation
2.2. Rabbits and Experimental Protocol
2.2.1. Feeding Trial
2.2.2. Experimental Infection and Clinical Trials
2.3. Growth Performance Traits and Clinical Examination
2.4. Samples Collection
2.5. Investigated Parameters
2.5.1. Quantitative DNA-Based Analysis of Intestinal Bacterial Populations
2.5.2. Expression Profiling by Reverse Transcription Quantitative Real-Time PCR (qRT-PCR) Assays

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Encoding Gene | Primer Sequence (5′-3′) | Accession No./Reference |
|---|---|---|
| Nutrient transporters | ||
| SGLT-1 | F:GATTTCCCGTATGATTACCGAG R:AAGAGGGAGACAACCACAACG | NM_001101692.1 |
| GLUT-2 | F-CAGAGGCACTGTCCACCACC R:TGTCTCCAAGCCACCCACC | NM_001277382.1 |
| CAT-1 | F:CCAGTCTATTAGGTTCCATGTTCC R: CGATTATTGGCGTTTTGGTC | XM_002721425.3 |
| FABP-2 | F: GTGGGGTTTTCCCTTTTGC R: CGCACTTTGGCCTTCACC | XM_002722970 |
| Tight junction proteins | ||
| MUC-2 | F:TATACCGCAAGCAGCCAGGT R:GCAAGCAGGACACAGACCAG | L41544.1 |
| JAM-2 | F:ATATCGCAGGTGTCCTGGAA R: GAGCATAGCACACGCCAAG | XM_017346699 |
| CLDN-1 | F: GGAGCAAAAGATGCGGATGG R: AATTGACAGGGGTCAAAGGGT | NM_001089316.1 |
| occludin | F: GCAAGAGGCCGTATCCAGAG R: AGTCCGTCTCGTAGTGGTCT | XM_008262320.1 |
| Cytokines | ||
| IL-6 | F:GCCAACCCTACAACAAGA R:AGAGCCACAACGACTGAC | NC_013678 |
| IL-8 | F:CTCTCTTGGCAACCTTCCTG R:TTGCACAGTGAGGTCCACTC | KT216053.1 |
| IL-10 | F:AAAAGCTAAAAGCCCCAGGA R:CGGGAGCTGAGGTATCAGAG | NM001082045.1 |
| IL-1β | F:TTCCGGATGTATCTCGAGCA R:GTGGATCGTGGTCGTCTTCA | NC_013670 |
| TNF-α | F:CTGCACTTCAGGGTGATCG R:CTACGTGGGCTAGAGGCTTG | XM_008262537.2 |
| IFNγ | F:TTCTTCAGCCTCACTCTCTCC R:TGTTGTCACTCTCCTCTTTCC | NM_001081991.1 |
| TGF-β1 | F: CAGTGGAAAGACCCCACATCTC R: GACGCAGGCAGCAATTATCC | NM_001082660 |
| House keeping | ||
| GAPDH | F:TGTTTGTGATGGGCGTGAA R:CCTCCACAATGCCGAAGT | NC_013676.1 |
| Listeria monocytogenes | ||
| 16S rRNA | F: CCTTTGACCACTCTGGAGACAGAGC R: AAGGAGGTGATCCAACCGCACCTTC | [39] |
| Total bacteria | ||
| 16S rRNA | F: CGGCAACGAGCGCAACCC R: CCATTGTAGCACGTGTGTAGCC | [40] |
| Genus Lactobacillus | ||
| 16S rRNA | F: AGCAGTAGGGAATCTTCCA R: CACCGCTACACATGGAG | [40] |
| Genus Bifidobacterium | ||
| 16S rRNA | F: TCGCGTCYGGTGTGAAAG R: CCACATCCAGCRTCCAC | [40] |
| Enterobacteriaceae | ||
| 16S rRNA | F: CATTGACGTTACCCGCAGAAGAAGC R: CTCTACGAGACTCAAGCTTGC | [40] |
2.5.3. Histomorphological Examination
2.6. Statistical Analysis
3. Results
3.1. Growth Performance and Clinical Observations
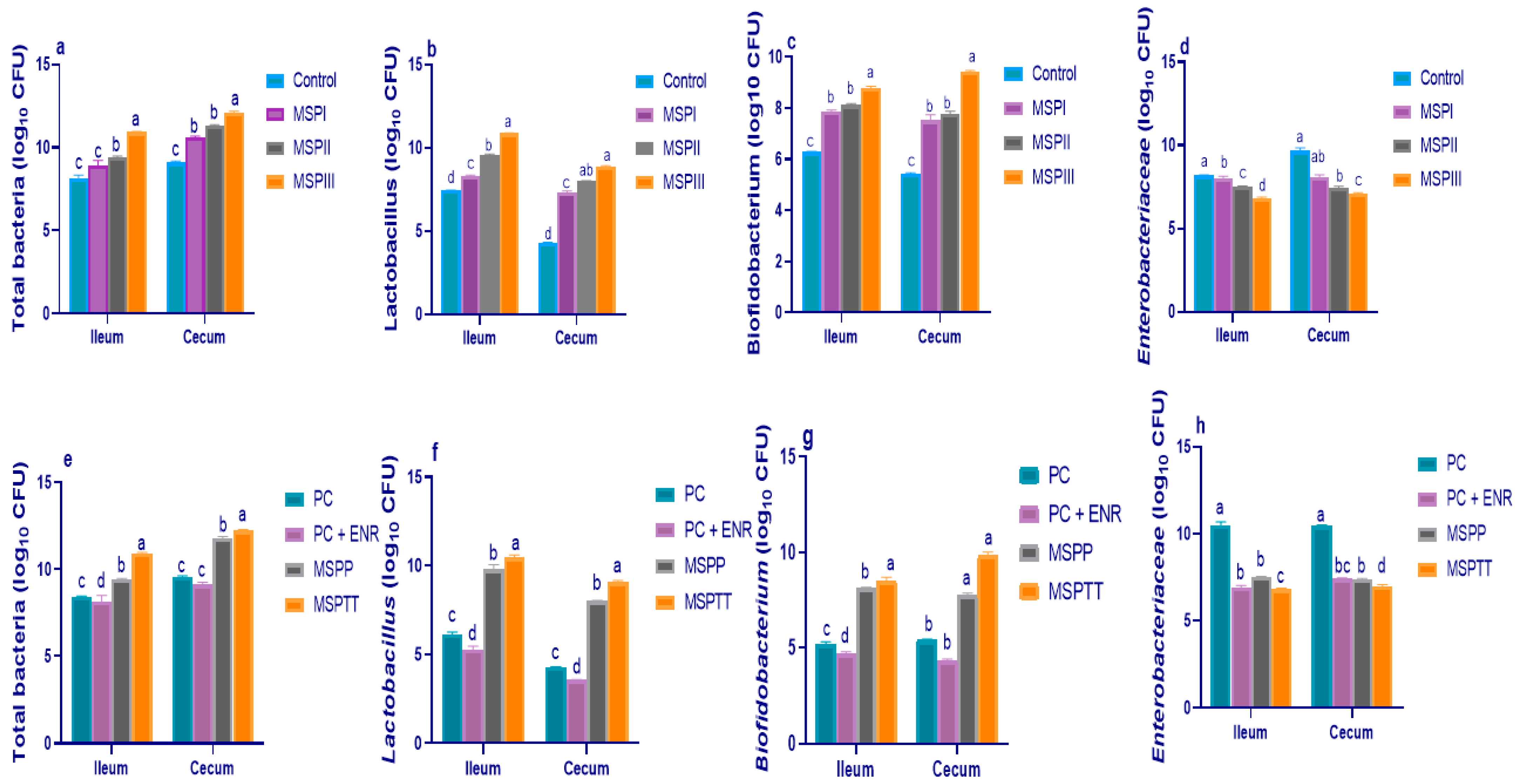
3.2. Microbial Populations of Intestinal Contents
3.3. Gene Expression Analysis
3.4. Quantification of L. monocytogenes
3.5. Histopathological Alterations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cullere, M.; Dalle Zotte, A. Rabbit meat production and consumption: State of knowledge and future perspectives. Meat Sci. 2018, 143, 137–146. [Google Scholar] [CrossRef]
- Simonová, M.P.; Lauková, A.; Žitňan, R.; Chrastinová, Ľ. Effect of rabbit-origin enterocin-producing probiotic strain Enterococcus faecium CCM7420 application on growth performance and gut morphometry in rabbits. Czech J. Anim. Sci. 2015, 60, 509–512. [Google Scholar] [CrossRef]
- Pastorelli, L.; De Salvo, C.; Mercado, J.R.; Vecchi, M.; Pizarro, T.T. Central role of the gut epithelial barrier in the pathogenesis of chronic intestinal inflammation: Lessons learned from animal models and human genetics. Front. Immunol. 2013, 4, 280. [Google Scholar] [CrossRef] [PubMed]
- Becattini, S.; Littmann, E.R.; Carter, R.A.; Kim, S.G.; Morjaria, S.M.; Ling, L.; Gyaltshen, Y.; Fontana, E.; Taur, Y.; Leiner, I.M. Commensal microbes provide first line defense against Listeria monocytogenes infection. J. Exp. Med. 2017, 214, 1973–1989. [Google Scholar] [CrossRef] [PubMed]
- Farahat, M.; Ibrahim, D.; Kishawy, A.; Abdallah, H.; Hernandez-Santana, A.; Attia, G. Effect of cereal type and plant extract addition on the growth performance, intestinal morphology, caecal microflora, and gut barriers gene expression of broiler chickens. Animal 2020, 15, 100056. [Google Scholar] [CrossRef] [PubMed]
- Michelland, R.J.; Combes, S.; Monteils, V.; Cauquil, L.; Gidenne, T.; Fortun-Lamothe, L. Molecular analysis of the bacterial community in digestive tract of rabbit. Anaerobe 2010, 16, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, D.; Ismail, T.A.; Khalifa, E.; El-Kader, A.; Shaimaa, A.; Mohamed, D.I.; Mohamed, D.T.; Shahin, S.E.; El-Hamid, A.; Marwa, I. Supplementing Garlic Nanohydrogel Optimized Growth, Gastrointestinal Integrity and Economics and Ameliorated Necrotic Enteritis in Broiler Chickens Using a Clostridium perfringens Challenge Model. Animals 2021, 11, 2027. [Google Scholar] [CrossRef]
- Kamada, N.; Chen, G.Y.; Inohara, N.; Núñez, G. Control of pathogens and pathobionts by the gut microbiota. Nat. Immunol. 2013, 14, 685–690. [Google Scholar] [CrossRef]
- Awad, N.F.; Abd El-Hamid, M.I.; Hashem, Y.; Erfan, A.; Abdelrahman, B.; Mahmoud, H.I. Impact of single and mixed infections with Escherichia coli and Mycoplasma gallisepticum on Newcastle disease virus vaccine performance in broiler chickens: An in vivo perspective. J. Appl. Microbiol. 2019, 127, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Johansson, J.; Freitag, N.E. Regulation of Listeria monocytogenes virulence. Microbiol. Spectr. 2019, 7, 1–20. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, F.; Chai, J.; Wang, J. Effect of lactic acid bacteria on Listeria monocytogenes infection and innate immunity in rabbits. Czech J. Anim. Sci. 2020, 65, 23–30. [Google Scholar] [CrossRef]
- De Cesare, A.; Parisi, A.; Mioni, R.; Comin, D.; Lucchi, A.; Manfreda, G. Listeria monocytogenes circulating in rabbit meat products and slaughterhouses in Italy: Prevalence data and comparison among typing results. Foodborne Pathog. Dis. 2017, 14, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Stratakos, A.C.; Ijaz, U.Z.; Ward, P.; Linton, M.; Kelly, C.; Pinkerton, L.; Scates, P.; McBride, J.; Pet, I.; Criste, A. In vitro and in vivo characterisation of Listeria monocytogenes outbreak isolates. Food Control 2020, 107, 106784. [Google Scholar] [CrossRef]
- Hegazy, W.A.; Salem, I.M.; Alotaibi, H.F.; Khafagy, E.-S.; Ibrahim, D. Terazosin Interferes with Quorum Sensing and Type Three Secretion System and Diminishes the Bacterial Espionage to Mitigate the Salmonella Typhimurium Pathogenesis. Antibiotics 2022, 11, 465. [Google Scholar] [CrossRef] [PubMed]
- Radoshevich, L.; Cossart, P. Listeria monocytogenes: Towards a complete picture of its physiology and pathogenesis. Nat. Rev. Microbiol. 2018, 16, 32–46. [Google Scholar] [CrossRef]
- Mesev, E.V.; Ledesma, R.A.; Ploss, A. Decoding type I and III interferon signalling during viral infection. Nat. Microbiol. 2019, 4, 914–924. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hamid, M.I.; Awad, N.F.; Hashem, Y.M.; Abdel-Rahman, M.A.; Abdelaziz, A.M.; Mohammed, I.A.; Abo-Shama, U.H. In vitro evaluation of various antimicrobials against field Mycoplasma gallisepticum and Mycoplasma synoviae isolates in Egypt. Poult. Sci. 2019, 98, 6281–6288. [Google Scholar] [CrossRef] [PubMed]
- Ammar, A.; Attia, A.; Abd El-Hamid, M.; El-Shorbagy, I.; Abd El-Kader, S. Genetic basis of resistance waves among methicillin resistant Staphylococcus aureus isolates recovered from milk and meat products in Egypt. Cell. Mol. Biol. 2016, 62, 7–15. [Google Scholar]
- Ammar, A.; El-Hamid, M.; Eid, S.E.; El Oksh, A.S. Insight into antimicrobial resistance and virulence genes of emergent multidrug resistant avian pathogenic Escherichia coli in Egypt: How closely related are they. Rev. Med. Vet 2015, 166, 304–314. [Google Scholar]
- Awad, N.F.; Hashem, Y.M.; Elshater, N.S.; Khalifa, E.; Hamed, R.I.; Nossieur, H.H.; Abd-Allah, E.M.; Elazab, S.T.; Nassan, M.A.; Abd El-Hamid, M.I. Therapeutic potentials of aivlosin and/or zinc oxide nanoparticles against Mycoplasma gallisepticum and/or Ornithobacterium rhinotracheale with a special reference to the effect of zinc oxide nanoparticles on aivlosin tissue residues: An in vivo approach. Poult. Sci. 2022, 101, 101884. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hamid, M.I.; El-Sayed, M.; Ali, A.R.; Abdallah, H.; Arnaout, M.I.; El-Mowalid, G.A. Marjoram extract down-regulates the expression of Pasteurella multocida adhesion, colonization and toxin genes: A potential mechanism for its antimicrobial activity. Comp. Immunol. Microbiol. Infect. Dis. 2019, 62, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Ammar, A.M.; El-Naenaeey, E.-S.Y.; El-Malt, R.M.; El-Gedawy, A.A.; Khalifa, E.; Elnahriry, S.S.; Abd El-Hamid, M.I. Prevalence, antimicrobial susceptibility, virulence and genotyping of Campylobacter jejuni with a special reference to the anti-virulence potential of Eugenol and beta-resorcylic acid on some multi-drug resistant isolates in Egypt. Animals 2020, 11, 3. [Google Scholar] [CrossRef]
- Amalaradjou, M.A.R.; Bhunia, A.K. Modern approaches in probiotics research to control foodborne pathogens. Adv. Food Nutr. Res. 2012, 67, 185–239. [Google Scholar] [PubMed]
- Suez, J.; Zmora, N.; Segal, E.; Elinav, E. The pros, cons, and many unknowns of probiotics. Nat. Med. 2019, 25, 716–729. [Google Scholar] [CrossRef]
- Mancini, S.; Paci, G. Probiotics in Rabbit Farming: Growth Performance, Health Status, and Meat Quality. Animals 2021, 11, 3388. [Google Scholar] [CrossRef]
- Rajoka, M.S.R.; Shi, J.; Zhu, J.; Shao, D.; Huang, Q.; Yang, H.; Jin, M. Capacity of lactic acid bacteria in immunity enhancement and cancer prevention. Appl. Microbiol. Biotechnol. 2017, 101, 35–45. [Google Scholar] [CrossRef]
- Ibrahim, D.; Moustafa, A.; Shahin, S.; Sherief, W.; Farag, M.; Nassan, M. Impact of fermented or enzymatically fermented dried olive pomace on growth, expression of digestive enzymes and glucose transporters genes, oxidative stability of frozen meat and economic efficiency of broiler chickens. Front. Vet. Sci. 2021, 8, 442. [Google Scholar] [CrossRef]
- Omar, A.E.; Al-Khalaifah, H.S.; Ismail, T.A.; El-Aziz, A.; Reda, M.; El-Mandrawy, S.A.; Shalaby, S.I.; Ibrahim, D. Performance, serum biochemical and immunological parameters, and digestive enzyme and intestinal barrier-related gene expression of broiler chickens fed fermented fava bean by-products as a substitute for conventional feed. Front. Vet. Sci. 2021, 8, 740. [Google Scholar] [CrossRef]
- Al-Khalaifah, H.S.; Shahin, S.E.; Omar, A.E.; Mohammed, H.A.; Mahmoud, H.I.; Ibrahim, D. Effects of graded levels of microbial fermented or enzymatically treated dried brewer’s grains on growth, digestive and nutrient transporter genes expression and cost effectiveness in broiler chickens. BMC Vet. Res. 2020, 16, 424. [Google Scholar] [CrossRef]
- De Blas, C.; Wiseman, J. The Nutrition of the Rabbit; CAB International: Wallingford, UK, 1998; p. 3441. [Google Scholar]
- López, M.B.; Briones, V.; Fernández-Garayzábal, J.F.; Vázquez-Boland, J.A.; García, J.A.; del Mar Blanco, M.; Suárez, G.; Domínguez, L. Serological response in rabbits to Listeria monocytogenes after oral or intragastric inoculation. FEMS Immunol. Med. Microbiol. 1993, 7, 131–134. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, F.; Chai, J.; Wang, J. Lactobacillus acidophilus reduces Listeria monocytogenes infection by inhibiting mitogen-activated protein kinase genes in growing rabbits. Rev. Bras. Zootec 2020, 49. [Google Scholar] [CrossRef]
- Ibrahim, D.; Abdelfattah-Hassan, A.; Arisha, A.H.; Abd El-Aziz, R.M.; Sherief, W.R.; Adil, S.H.; El Sayed, R.; Metwally, A.E. Impact of feeding anaerobically fermented feed supplemented with acidifiers on its quality and growth performance, intestinal villi and enteric pathogens of mulard ducks. Livest. Sci. 2020, 242, 104299. [Google Scholar] [CrossRef]
- Ibrahim, D.; Sewid, A.H.; Arisha, A.H.; Abd El-Fattah, A.H.; Abdelaziz, A.M.; Al-Jabr, O.A.; Kishawy, A.T. Influence of Glycyrrhiza glabra Extract on Growth, Gene Expression of Gut Integrity, and Campylobacter jejuni Colonization in Broiler Chickens. Front. Vet. Sci. 2020, 7, 612063. [Google Scholar] [CrossRef] [PubMed]
- Blasco, A.; Ouhayoun, J. Harmonization of criteria and terminology in rabbit meat research. Revised proposal. World Rabbit Sci. 1996, 4, 93–99. [Google Scholar] [CrossRef]
- Ibrahim, D.; El Sayed, R.; Abdelfattah-Hassan, A.; Morshedy, A. Creatine or guanidinoacetic acid? Which is more effective at enhancing growth, tissue creatine stores, quality of meat, and genes controlling growth/myogenesis in Mulard ducks. J. Appl. Anim. Res. 2019, 47, 159–166. [Google Scholar] [CrossRef]
- Ibrahim, D.; Al-Khalaifah, H.S.; Abdelfattah-Hassan, A.; Eldoumani, H.; Khater, S.I.; Arisha, A.H.; Mohamed, S.A.; Ismail, T.A.; Tolba, S.A. Promising Role of Growth Hormone-Boosting Peptide in Regulating the Expression of Muscle-Specific Genes and Related MicroRNAs in Broiler Chickens. Animals 2021, 11, 1906. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Lantz, P.-G.; Tjerneld, F.; Borch, E.; Hahn-Hägerdal, B.; Rådström, P. Enhanced sensitivity in PCR detection of Listeria monocytogenes in soft cheese through use of an aqueous two-phase system as a sample preparation method. Appl. Environ. Microbiol. 1994, 60, 3416–3418. [Google Scholar] [CrossRef]
- Liu, L.; Zeng, D.; Yang, M.; Wen, B.; Lai, J.; Zhou, Y.; Sun, H.; Xiong, L.; Wang, J.; Lin, Y. Probiotic Clostridium butyricum improves the growth performance, immune function, and gut microbiota of weaning rex rabbits. Probiotics Antimicrob. Proteins 2019, 11, 1278–1292. [Google Scholar] [CrossRef]
- Ibrahim, D.; Abdelfattah-Hassan, A.; Badawi, M.; Ismail, T.A.; Bendary, M.M.; Abdelaziz, A.M.; Mosbah, R.A.; Mohamed, D.I.; Arisha, A.H.; El-Hamid, M.I.A. Thymol nanoemulsion promoted broiler chicken’s growth, gastrointestinal barrier and bacterial community and conferred protection against Salmonella Typhimurium. Sci. Rep. 2021, 11, 7742. [Google Scholar] [CrossRef]
- Alasmari, W.A.; Abdelfattah-Hassan, A.; El-Ghazali, H.M.; Abdo, S.A.; Ibrahim, D.; ElSawy, N.A.; El-Shetry, E.S.; Saleh, A.A.; Abourehab, M.A.; Mahfouz, H. Exosomes Derived from BM-MSCs Mitigate the Development of Chronic Kidney Damage Post-Menopause via Interfering with Fibrosis and Apoptosis. Biomolecules 2022, 12, 663. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Wu, F.; Hao, G.; Qi, Q.; Li, R.; Li, N.; Wei, L.; Chai, T. Bacillus subtilis improves immunity and disease resistance in rabbits. Front. Immunol. 2017, 8, 354. [Google Scholar] [CrossRef]
- Shim, Y.; Shinde, P.; Choi, J.; Kim, J.; Seo, D.; Pak, J.; Chae, B.; Kwon, I. Evaluation of multi-microbial probiotics produced by submerged liquid and solid substrate fermentation methods in broilers. Asian-Australas. J. Anim. Sci. 2010, 23, 521–529. [Google Scholar] [CrossRef]
- Sen, S.; Ingale, S.; Kim, J.; Kim, K.; Kim, Y.; Khong, C.; Lohakare, J.; Kim, E.; Kim, H.; Kwon, I. Effect of supplementation of Bacillus subtilis LS 1-2 grown on citrus-juice waste and corn-soybean meal substrate on growth performance, nutrient retention, caecal microbiology and small intestinal morphology of broilers. Asian-Australas. J. Anim. Sci. 2011, 24, 1120–1127. [Google Scholar] [CrossRef]
- Chen, W.; Wang, J.; Yan, L.; Huang, Y. Evaluation of probiotics in diets with different nutrient densities on growth performance, blood characteristics, relative organ weight and breast meat characteristics in broilers. Br. Poult. Sci. 2013, 54, 635–641. [Google Scholar] [CrossRef]
- Sanders, M.E.; Lenoir-Wijnkoop, I.; Salminen, S.; Merenstein, D.J.; Gibson, G.R.; Petschow, B.W.; Nieuwdorp, M.; Tancredi, D.J.; Cifelli, C.J.; Jacques, P. Probiotics and prebiotics: Prospects for public health and nutritional recommendations. Ann. N. Y. Acad. Sci. 2014, 1309, 19–29. [Google Scholar] [CrossRef]
- Phuoc, T.L.; Jamikorn, U. Effects of probiotic supplement (Bacillus subtilis and Lactobacillus acidophilus) on feed efficiency, growth performance, and microbial population of weaning rabbits. Asian-Australas. J. Anim. Sci. 2017, 30, 198–205. [Google Scholar] [CrossRef]
- Giang, H.H.; Viet, T.Q.; Ogle, B.; Lindberg, J.E. Growth performance, digestibility, gut environment and health status in weaned piglets fed a diet supplemented with potentially probiotic complexes of lactic acid bacteria. Livest. Sci. 2010, 129, 95–103. [Google Scholar] [CrossRef]
- Neveling, D.; Van Emmenes, L.; Ahire, J.; Pieterse, E.; Smith, C.; Dicks, L. Safety assessment of antibiotic and probiotic feed additives for Gallus gallus domesticus. Sci. Rep. 2017, 7, 12767. [Google Scholar] [CrossRef]
- Cross, M.L. Microbes versus microbes: Immune signals generated by probiotic lactobacilli and their role in protection against microbial pathogens. FEMS Immunol. Med. Microbiol. 2002, 34, 245–253. [Google Scholar] [CrossRef]
- O’Hara, A.M.; Shanahan, F. The gut flora as a forgotten organ. EMBO Rep. 2006, 7, 688–693. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Polk, D.B. Probiotics and Probiotic-Derived Functional Factors—Mechanistic Insights Into Applications for Intestinal Homeostasis. Front. Immunol. 2020, 11, 1428. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zeng, Y.; Wang, S.; Liu, H.; Zhang, D.; Zhang, W.; Wang, Y.; Ji, H. Swine-derived probiotic Lactobacillus plantarum inhibits growth and adhesion of enterotoxigenic Escherichia coli and mediates host defense. Front. Microbiol. 2018, 9, 1364. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, C.; Donders, G.; Palmeira-de-Oliveira, R.; Queiroz, J.; Tomaz, C.; Martinez-de-Oliveira, J.; Palmeira-de-Oliveira, A. Bacteriocin production of the probiotic Lactobacillus acidophilus KS400. Amb Express 2018, 8, 153. [Google Scholar] [CrossRef] [PubMed]
- Pu, J.; Chen, D.; Tian, G.; He, J.; Zheng, P.; Mao, X.; Yu, J.; Huang, Z.; Zhu, L.; Luo, J. Protective effects of benzoic acid, Bacillus coagulans, and oregano oil on intestinal injury caused by enterotoxigenic Escherichia coli in weaned piglets. BioMed Res. Int. 2018, 2018, 1829632. [Google Scholar] [CrossRef]
- Gorboulev, V.; Schürmann, A.; Vallon, V.; Kipp, H.; Jaschke, A.; Klessen, D.; Friedrich, A.; Scherneck, S.; Rieg, T.; Cunard, R. Na+-D-glucose cotransporter SGLT1 is pivotal for intestinal glucose absorption and glucose-dependent incretin secretion. Diabetes 2012, 61, 187–196. [Google Scholar] [CrossRef]
- Wright, E.; Hirayama, B.; Loo, D. Active sugar transport in health and disease. J. Intern. Med. 2007, 261, 32–43. [Google Scholar] [CrossRef]
- Boudry, G.; Cheeseman, C.I.; Perdue, M.H. Psychological stress impairs Na+-dependent glucose absorption and increases GLUT2 expression in the rat jejunal brush-border membrane. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2007, 292, R862–R867. [Google Scholar] [CrossRef]
- Zhao, J.; Bao, J.; Wang, X.; Jiao, H.; Song, Z.; Lin, H. Altered gene and protein expression of glucose transporter1 underlies dexamethasone inhibition of insulin-stimulated glucose uptake in chicken muscles. J. Anim. Sci. 2012, 90, 4337–4345. [Google Scholar] [CrossRef]
- Ibrahim, D.; Moustafa, A.; Metwally, A.S.; Nassan, M.A.; Abdallah, K.; Eldemery, F.; Tufarelli, V.; Laudadio, V.; Kishawy, A.T. Potential Application of Cornelian Cherry Extract on Broiler Chickens: Growth, Expression of Antioxidant Biomarker and Glucose Transport Genes, and Oxidative Stability of Frozen Meat. Animals 2021, 11, 1038. [Google Scholar] [CrossRef]
- Musso, G.; Gambino, R.; Cassader, M. Interactions between gut microbiota and host metabolism predisposing to obesity and diabetes. Annu. Rev. Med. 2011, 62, 361–380. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Mahender, M.; Prakash, M.G.; Raghunandan, T.; Reddy, K.K. Productive performance of broiler rabbits fed diets supplemented with probiotic and enzymes under two systems of housing. Indian J. Anim. Res. 2014, 48, 355–361. [Google Scholar] [CrossRef]
- Biswas, A.; Dev, K.; Tyagi, P.K.; Mandal, A. The effect of multi-strain probiotics as feed additives on performance, immunity, expression of nutrient transporter genes and gut morphometry in broiler chickens. Anim. Biosci. 2022, 35, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Castanys-Muñoz, E.; Martin, M.J.; Vazquez, E. Building a beneficial microbiome from birth. Adv. Nutr. 2016, 7, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Boirivant, M.; Strober, W. The mechanism of action of probiotics. Curr. Opin. Gastroenterol. 2007, 23, 679–692. [Google Scholar] [CrossRef]
- Ewuola, E.; Amadi, C.; Imam, T. Performance evaluation and nutrient digestibility of rabbits fed dietary prebiotics, probiotics and symbiotics. Int. J. Appl. Agric. Apic. Res. 2011, 7, 107–117. [Google Scholar]
- Li, J.; Sung, C.Y.J.; Lee, N.; Ni, Y.; Pihlajamäki, J.; Panagiotou, G.; El-Nezami, H. Probiotics modulated gut microbiota suppresses hepatocellular carcinoma growth in mice. Proc. Natl. Acad. Sci. USA 2016, 113, E1306–E1315. [Google Scholar] [CrossRef] [PubMed]
- Galdeano, C.M.; Cazorla, S.I.; Dumit, J.M.L.; Vélez, E.; Perdigón, G. Beneficial effects of probiotic consumption on the immune system. Ann. Nutr. Metab. 2019, 74, 115–124. [Google Scholar]
- Seyidoglu, N.; Peker, S. Effects of different doses of probiotic yeast Saccharomyces cerevisiae on the duodenal mucosa in rabbits. Indian J. Anim. Res. 2015, 49, 602–606. [Google Scholar] [CrossRef]
- Oso, A.; Idowu, O.; Haastrup, A.; Ajibade, A.; Olowonefa, K.; Aluko, A.; Ogunade, I.; Osho, S.; Bamgbose, A. Growth performance, apparent nutrient digestibility, caecal fermentation, ileal morphology and caecal microflora of growing rabbits fed diets containing probiotics and prebiotics. Livest. Sci. 2013, 157, 184–190. [Google Scholar] [CrossRef]
- Røssland, E.; Langsrud, T.; Granum, P.E.; Sørhaug, T. Production of antimicrobial metabolites by strains of Lactobacillus or Lactococcus co-cultured with Bacillus cereus in milk. Int. J. Food Microbiol. 2005, 98, 193–200. [Google Scholar] [CrossRef]
- Becattini, S.; Taur, Y.; Pamer, E.G. Antibiotic-induced changes in the intestinal microbiota and disease. Trends Mol. Med. 2016, 22, 458–478. [Google Scholar] [CrossRef] [PubMed]
- Ohh, S.; Shinde, P.; Choi, J.; Jin, Z.; Hahn, T.; Lim, H.; Kim, G.; Park, Y.; Hahm, K.; Chae, B. Effects of potato (Solanum tuberosum I. cv. golden valley) protein on performance, nutrient metabolizability, and cecal microflora in broilers. Arch. Fur Geflügelkunde 2010, 74, 30–35. [Google Scholar]
- Tang, J.; Mohamad Ghazali, F.; Saleha, A.; Nishibuchi, M.; Son, R. Comparison of thermophilic Campylobacter spp. occurrence in two types of retail chicken samples. Int. Food Res. J. 2009, 16, 277–288. [Google Scholar]
- Proctor, A.; Phillips, G.J. Differential effects of bacitracin methylene disalicylate (BMD) on the distal colon and cecal microbiota of young broiler chickens. Front. Vet. Sci. 2019, 6, 114. [Google Scholar] [CrossRef] [PubMed]
- Koltes, D.A.; Lester, H.D.; Frost, M.; Aldridge, D.; Christensen, K.D.; Scanes, C.G. Effects of bacitracin methylene disalicylate and diet change on gastrointestinal integrity and endotoxin permeability in the duodenum of broiler chicken. BMC Res. Notes 2017, 10, 470. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.; Gueimonde, M.; Salminen, S. Probiotics in adhesion of pathogens: Mechanisms of action. In Bioactive Foods in Promoting Health; Academic Press: New York, NY, USA, 2010. [Google Scholar]
- Corthay, A. A three-cell model for activation of naive T helper cells. Scand. J. Immunol. 2006, 64, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Takahashi, K.; Tohno, M.; Miura, Y.; Kamada, T.; Ikegami, S.; Kitazawa, H. Immunomodulation in gut-associated lymphoid tissue of neonatal chicks by immunobiotic diets. Poult. Sci. 2009, 88, 2532–2538. [Google Scholar] [CrossRef]
- Liu, N.; Deng, X.; Liang, C.; Cai, H. Effect of broccoli residues fermented with probiotics on the growth performance and health status of broilers challenged with Clostridium perfringens. Braz. J. Poult. Sci. 2018, 20, 625–632. [Google Scholar] [CrossRef]
- Villena, J.; Chiba, E.; Vizoso-Pinto, M.G.; Tomosada, Y.; Takahashi, T.; Ishizuka, T.; Aso, H.; Salva, S.; Alvarez, S.; Kitazawa, H. Immunobiotic Lactobacillus rhamnosus strains differentially modulate antiviral immune response in porcine intestinal epithelial and antigen presenting cells. BMC Microbiol. 2014, 14, 126. [Google Scholar] [CrossRef] [PubMed]
- Kopp, M.; Goldstein, M.; Dietschek, A.; Sofke, J.; Heinzmann, A.; Urbanek, R. Lactobacillus GG has in vitro effects on enhanced interleukin-10 and interferon-γ release of mononuclear cells but no in vivo effects in supplemented mothers and their neonates. Clin. Exp. Allergy 2008, 38, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Wan, Z.; Wang, L.; Chen, Z.; Ma, X.; Yang, X.; Zhang, J.; Jiang, Z. In vitro evaluation of swine-derived Lactobacillus reuteri: Probiotic properties and effects on intestinal porcine epithelial cells challenged with enterotoxigenic Escherichia coli K88. J. Microbiol. Biotechnol. 2016, 26, 1018–1025. [Google Scholar]
- Farkas, O.; Mátis, G.; Pászti-Gere, E.; Palócz, O.; Kulcsár, A.; Petrilla, J.; Csikó, G.; Neogrády, Z.; Gálfi, P. Effects of Lactobacillus plantarum 2142 and sodium n-butyrate in lipopolysaccharide-triggered inflammation: Comparison of a porcine intestinal epithelial cell line and primary hepatocyte monocultures with a porcine enterohepatic co-culture system. J. Anim. Sci. 2014, 92, 3835–3845. [Google Scholar] [CrossRef]
- Azad, M.; Kalam, A.; Sarker, M.; Wan, D. Immunomodulatory effects of probiotics on cytokine profiles. BioMed Res. Int. 2018, 2018, 8063647. [Google Scholar] [CrossRef] [PubMed]
- Kayamuro, H.; Yoshioka, Y.; Abe, Y.; Arita, S.; Katayama, K.; Nomura, T.; Yoshikawa, T.; Kubota-Koketsu, R.; Ikuta, K.; Okamoto, S. Interleukin-1 family cytokines as mucosal vaccine adjuvants for induction of protective immunity against influenza virus. J. Virol. 2010, 84, 12703–12712. [Google Scholar] [CrossRef]
- Drolia, R.; Tenguria, S.; Durkes, A.C.; Turner, J.R.; Bhunia, A.K. Listeria adhesion protein induces intestinal epithelial barrier dysfunction for bacterial translocation. Cell Host Microbe 2018, 23, 470–484.e7. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, H.; Sahebani, N.; Mirabolfathy, M.; Faravardeh, L.; Mahdavi, V. The effect of salicylic acid and Bacillus subtilis on cucumber root and stem rot, caused by Fusarium oxysporum f. sp. radicis cucumerinum. Iran. J. Plant Pathol. 2011, 46, 85–87. [Google Scholar]
- Gómez-Llorente, C.; Munoz, S.; Gil, A. Role of Toll-like receptors in the development of immunotolerance mediated by probiotics. Proc. Nutr. Soc. 2010, 69, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Harty, J.T.; Bevant, M.J. Specific immunity to Listeria monocytogenes in the absence of IFNγ. Immunity 1995, 3, 109–117. [Google Scholar] [CrossRef]
- Wei, X.; Zhang, Y.; Zhou, H.; Tian, F.; Ni, Y. Antimicrobial activities and in vitro properties of cold-adapted Lactobacillus strains isolated from the intestinal tract of cold water fishes of high latitude water areas in Xinjiang, China. BMC Microbiol. 2019, 19, 247. [Google Scholar] [CrossRef]
- D’Arienzo, R.; Maurano, F.; Mazzarella, G.; Luongo, D.; Stefanile, R.; Ricca, E.; Rossi, M. Bacillus subtilis spores reduce susceptibility to Citrobacter rodentium-mediated enteropathy in a mouse model. Res. Microbiol. 2006, 157, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Johnson-Henry, K.C.; Nadjafi, M.; Avitzur, Y.; Mitchell, D.J.; Ngan, B.-Y.; Galindo-Mata, E.; Jones, N.L.; Sherman, P.M. Amelioration of the effects of Citrobacter rodentium infection in mice by pretreatment with probiotics. J. Infect. Dis. 2005, 191, 2106–2117. [Google Scholar] [CrossRef]
- Dos Santos, L.M.; Santos, M.M.; de Souza Silva, H.P.; Arantes, R.M.E.; Nicoli, J.R.; Vieira, L.Q. Monoassociation with probiotic Lactobacillus delbrueckii UFV-H2b20 stimulates the immune system and protects germfree mice against Listeria monocytogenes infection. Med. Microbiol. Immunol. 2011, 200, 29–38. [Google Scholar] [CrossRef]
- Foye, O.T.; Huang, I.-F.; Chiou, C.C.; Walker, W.A.; Shi, H.N. Early administration of probiotic Lactobacillus acidophilus and/or prebiotic inulin attenuates pathogen-mediated intestinal inflammation and Smad 7 cell signaling. FEMS Immunol. Med. Microbiol. 2012, 65, 467–480. [Google Scholar] [CrossRef]
- Oh, S.; Lillehoj, H.S.; Lee, Y.; Bravo, D.; Lillehoj, E.P. Dietary antibiotic growth promoters down-regulate intestinal inflammatory cytokine expression in chickens challenged with LPS or co-infected with Eimeria maxima and Clostridium perfringens. Front. Vet. Sci. 2019, 6, 420. [Google Scholar] [CrossRef]
- Gadde, U.D.; Oh, S.; Lee, Y.; Davis, E.; Zimmerman, N.; Rehberger, T.; Lillehoj, H.S. Dietary Bacillus subtilis-based direct-fed microbials alleviate LPS-induced intestinal immunological stress and improve intestinal barrier gene expression in commercial broiler chickens. Res. Vet. Sci. 2017, 114, 236–243. [Google Scholar] [CrossRef]
- Alandiyjany, M.N.; Abdelaziz, A.S.; Abdelfattah-Hassan, A.; Hegazy, W.A.; Hassan, A.A.; Elazab, S.T.; Mohamed, E.A.; El-Shetry, E.S.; Saleh, A.A.; ElSawy, N.A. Novel In Vivo Assessment of Antimicrobial Efficacy of Ciprofloxacin Loaded Mesoporous Silica Nanoparticles against Salmonella typhimurium Infection. Pharmaceuticals 2022, 15, 357. [Google Scholar] [CrossRef] [PubMed]
- Marchiando, A.M.; Graham, W.V.; Turner, J.R. Epithelial barriers in homeostasis and disease. Annu. Rev. Pathol. Mech. Dis. 2010, 5, 119–144. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Shen, L.; Weber, C.R.; Raleigh, D.R.; Yu, D.; Turner, J.R. Tight junction pore and leak pathways: A dynamic duo. Annu. Rev. Physiol. 2011, 73, 283–309. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, D.; Eldemery, F.; Metwally, A.S.; Abd-Allah, E.M.; Mohamed, D.T.; Ismail, T.A.; Hamed, T.A.; Al Sadik, G.M.; Neamat-Allah, A.N.; El-Hamid, A. Dietary Eugenol Nanoemulsion Potentiated Performance of Broiler Chickens: Orchestration of Digestive Enzymes, Intestinal Barrier Functions and Cytokines Related Gene Expression With a Consequence of Attenuating the Severity of E. coli O78 Infection. Front. Vet. Sci. 2022, 9, 847580. [Google Scholar] [CrossRef] [PubMed]
- Slifer, Z.M.; Blikslager, A.T. The integral role of tight junction proteins in the repair of injured intestinal epithelium. Int. J. Mol. Sci. 2020, 21, 972. [Google Scholar] [CrossRef] [PubMed]
- Karl, J.P.; Margolis, L.M.; Madslien, E.H.; Murphy, N.E.; Castellani, J.W.; Gundersen, Y.; Hoke, A.V.; Levangie, M.W.; Kumar, R.; Chakraborty, N. Changes in intestinal microbiota composition and metabolism coincide with increased intestinal permeability in young adults under prolonged physiological stress. Am. J. Physiol.-Gastrointest. Liver Physiol. 2017, 312, G559–G571. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Zhao, D.-H.; Jiang, M. VSL# 3 probiotics regulate the intestinal epithelial barrier in vivo and in vitro via the p38 and ERK signaling pathways. Int. J. Mol. Med. 2012, 29, 202–208. [Google Scholar] [PubMed]
- Stavropoulou, E.; Bezirtzoglou, E. Probiotics as a weapon in the fight against COVID-19. Front. Nutr. 2020, 7, 614986. [Google Scholar] [CrossRef]
- Alkushi, A.G.; Elazab, S.T.; Abdelfattah-Hassan, A.; Mahfouz, H.; Salem, G.A.; Sheraiba, N.I.; Mohamed, E.A.; Attia, M.S.; El-Shetry, E.S.; Saleh, A.A. Multi-Strain-Probiotic-Loaded Nanoparticles Reduced Colon Inflammation and Orchestrated the Expressions of Tight Junction, NLRP3 Inflammasome and Caspase-1 Genes in DSS-Induced Colitis Model. Pharmaceutics 2022, 14, 1183. [Google Scholar] [CrossRef]
- Anderson, R.C.; Cookson, A.L.; McNabb, W.C.; Park, Z.; McCann, M.J.; Kelly, W.J.; Roy, N.C. Lactobacillus plantarum MB452 enhances the function of the intestinal barrier by increasing the expression levels of genes involved in tight junction formation. BMC Microbiol. 2010, 10, 316. [Google Scholar] [CrossRef]
- Blackwood, B.P.; Yuan, C.Y.; Wood, D.R.; Nicolas, J.D.; Grothaus, J.S.; Hunter, C.J. Probiotic Lactobacillus species strengthen intestinal barrier function and tight junction integrity in experimental necrotizing enterocolitis. J. Probiotics Health 2017, 5, 159. [Google Scholar] [CrossRef]
- Wu, Y.; Zhu, C.; Chen, Z.; Chen, Z.; Zhang, W.; Ma, X.; Wang, L.; Yang, X.; Jiang, Z. Protective effects of Lactobacillus plantarum on epithelial barrier disruption caused by enterotoxigenic Escherichia coli in intestinal porcine epithelial cells. Vet. Immunol. Immunopathol. 2016, 172, 55–63. [Google Scholar] [CrossRef]
- Wang, J.; Lin, L.; Jiang, Q.; Huang, W.; Liu, N. Probiotics and clay detoxifier protected growth performance and intestinal barrier of lambs fed diet contaminated with aflatoxin B-1. Indian J. Anim. Sci. 2019, 89, 658–662. [Google Scholar]
- Drolia, R.; Amalaradjou, M.A.R.; Ryan, V.; Tenguria, S.; Liu, D.; Bai, X.; Xu, L.; Singh, A.K.; Cox, A.D.; Bernal-Crespo, V. Receptor-targeted engineered probiotics mitigate lethal Listeria infection. Nat. Commun. 2020, 11, 6344. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Zhang, Z.; Hang, X.; Jiang, Y.L. plantarum prevents enteroinvasive Escherichia coli-induced tight junction proteins changes in intestinal epithelial cells. BMC Microbiol. 2009, 9, 63. [Google Scholar] [CrossRef] [PubMed]
- Bhat, M.I.; Kapila, S.; Kapila, R. Lactobacillus fermentum (MTCC-5898) supplementation renders prophylactic action against Escherichia coli impaired intestinal barrier function through tight junction modulation. LWT 2020, 123, 109118. [Google Scholar] [CrossRef]
- de LeBlanc, A.d.M.; Dogi, C.A.; Galdeano, C.M.; Carmuega, E.; Weill, R.; Perdigón, G. Effect of the administration of a fermented milk containing Lactobacillus casei DN-114001 on intestinal microbiota and gut associated immune cells of nursing mice and after weaning until immune maturity. BMC Immunol. 2008, 9, 27. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Kim, S.-H.; Whang, K.-Y.; Kim, Y.-J.; Oh, S.-J. Inhibition of Escherichia coli O157: H7 attachment by interactions between lactic acid bacteria and intestinal epithelial cells. J. Microbiol. Biotechnol. 2008, 18, 1278–1285. [Google Scholar]
- Mack, D.R.; Ahrné, S.; Hyde, L.; Wei, S.; Hollingsworth, M.A. Extracellular MUC3 mucin secretion follows adherence of Lactobacillus strains to intestinal epithelial cells in vitro. Gut 2003, 52, 827–833. [Google Scholar] [CrossRef]
- Otte, J.-M.; Podolsky, D.K. Functional modulation of enterocytes by gram-positive and gram-negative microorganisms. Am. J. Physiol.-Gastrointest. Liver Physiol. 2004, 286, G613–G626. [Google Scholar] [CrossRef]
- Feng, Y.; Huang, Y.; Wang, Y.; Wang, P.; Song, H.; Wang, F. Antibiotics induced intestinal tight junction barrier dysfunction is associated with microbiota dysbiosis, activated NLRP3 inflammasome and autophagy. PLoS ONE 2019, 14, e0218384. [Google Scholar] [CrossRef]
- Regan, T.; MacSharry, J.; Brint, E. Tracing innate immune defences along the path of Listeria monocytogenes infection. Immunol. Cell Biol. 2014, 92, 563–569. [Google Scholar] [CrossRef]
- Ruppé, E.; Burdet, C.; Grall, N.; De Lastours, V.; Lescure, F.-X.; Andremont, A.; Armand-Lefèvre, L. Impact of antibiotics on the intestinal microbiota needs to be re-defined to optimize antibiotic usage. Clin. Microbiol. Infect. 2018, 24, 3–5. [Google Scholar] [CrossRef]
- O’Toole, P.W.; Marchesi, J.R.; Hill, C. Next-generation probiotics: The spectrum from probiotics to live biotherapeutics. Nat. Microbiol. 2017, 2, 17057. [Google Scholar] [CrossRef] [PubMed]
- Scaffaro, R.; Lopresti, F.; Marino, A.; Nostro, A. Antimicrobial additives for poly (lactic acid) materials and their applications: Current state and perspectives. Appl. Microbiol. Biotechnol. 2018, 102, 7739–7756. [Google Scholar] [CrossRef] [PubMed]
- Ehsani, A.; Rezaeiyan, A.; Hashemi, M.; Aminzare, M.; Jannat, B.; Afshari, A. Antibacterial activity and sensory properties of Heracleum persicum essential oil, nisin, and Lactobacillus acidophilus against Listeria monocytogenes in cheese. Vet. World 2019, 12, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Rocha, K.R.; Perini, H.F.; de Souza, C.M.; Schueler, J.; Tosoni, N.F.; Furlaneto, M.C.; Furlaneto-Maia, L. Inhibitory effect of bacteriocins from enterococci on developing and preformed biofilms of Listeria monocytogenes, Listeria ivanovii and Listeria innocua. World J. Microbiol. Biotechnol. 2019, 35, 96. [Google Scholar] [CrossRef] [PubMed]
- Watson, G.; Evans, M. Listeriosis in a rabbit. Vet. Pathol. 1985, 22, 191–193. [Google Scholar] [CrossRef] [PubMed]
- Olaoye, S.; Adedire, O.; Fajobi, A.; Farinu, A.; Ademolue, O.; Ogundipe, W. Impact of probiotic feed fermentation on the performance characteristics and histology of post-weaned rabbits. In Proceedings of the 25th Annual Conference of Animal Science Association of Nigeria, Abuja, Nigeria, 1–4 November 2020. [Google Scholar]








| Ingredient | % |
|---|---|
| Yellow corn | 10 |
| Barley grains | 16.3 |
| Soybean meal, 44% | 15.70 |
| Berseem hay | 33.2 |
| Wheat bran | 19 |
| Molasses | 3.00 |
| Premix * | 0.3 |
| Calcium dibasic phosphate | 1.5 |
| Common salt | 0.5 |
| Antitoxin | 0.3 |
| Anticoccidial | 0.2 |
| Nutrient level | |
| DE, Kcal/Kg | 2555.60 |
| CP | 16.33 |
| EE | 2.33 |
| CF | 12.56 |
| Ca | 1.09 |
| Phosphorus | 0.59 |
| Parameter | Experimental Groups | p Value | SEM | |||
|---|---|---|---|---|---|---|
| Control | MSPI | MSPII | MSPIII | |||
| BW, g | 1818 d | 2006 c | 2147 b | 2176 a | <0.02 | 12.14 |
| BWG, g | 1086 d | 1204 c | 1344 b | 1372 a | <0.001 | 13.16 |
| FI, g | 2700 c | 2991 b | 3084 a | 2942 b | 0.03 | 14.83 |
| FCR | 2.49 a | 2.48 a | 2.29 b | 2.14 c | 0.01 | <0.001 |
| Parameter | Experimental Groups | p Value | SEM | |||
|---|---|---|---|---|---|---|
| PC | PC + ENR | MSPP | MSPTT | |||
| BW, g | 2110 d | 2664 b | 2431c | 2742 a | <0.001 | 34.93 |
| BWG, g | 1308 d | 1862 b | 1627 c | 1938 a | <0.001 | 33.34 |
| FI, g | 4832 d | 5756 a | 5214 c | 5654 b | 0.04 | 15.37 |
| FCR | 3.69 a | 3.09 c | 3.20 b | 2.92 d | 0.02 | 0.09 |
| Mortality % | 24.00 d | 7.80 b | 20.00 c | 6.00 a | <0.001 | 0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd El-Hamid, M.I.; Ibrahim, D.; Hamed, R.I.; Nossieur, H.H.; Elbanna, M.H.; Baz, H.; Abd-Allah, E.M.; El Oksh, A.S.A.; Ibrahim, G.A.; Khalifa, E.; et al. Modulatory Impacts of Multi-Strain Probiotics on Rabbits’ Growth, Nutrient Transporters, Tight Junctions and Immune System to Fight against Listeria monocytogenes Infection. Animals 2022, 12, 2082. https://doi.org/10.3390/ani12162082
Abd El-Hamid MI, Ibrahim D, Hamed RI, Nossieur HH, Elbanna MH, Baz H, Abd-Allah EM, El Oksh ASA, Ibrahim GA, Khalifa E, et al. Modulatory Impacts of Multi-Strain Probiotics on Rabbits’ Growth, Nutrient Transporters, Tight Junctions and Immune System to Fight against Listeria monocytogenes Infection. Animals. 2022; 12(16):2082. https://doi.org/10.3390/ani12162082
Chicago/Turabian StyleAbd El-Hamid, Marwa I., Doaa Ibrahim, Rehab I. Hamed, Heba H. Nossieur, Mariam Hassan Elbanna, Heba Baz, Ehab. M. Abd-Allah, Amal S. A. El Oksh, Ghada A. Ibrahim, Eman Khalifa, and et al. 2022. "Modulatory Impacts of Multi-Strain Probiotics on Rabbits’ Growth, Nutrient Transporters, Tight Junctions and Immune System to Fight against Listeria monocytogenes Infection" Animals 12, no. 16: 2082. https://doi.org/10.3390/ani12162082