
The Distribution Characteristics of Aerosol Bacteria in Different Types of Pig Houses
by
 ,
,
Huan Cui
1,2,†, 
Cheng Zhang
1,3,†,
Juxiang Liu
3,
Shishan Dong
3,
Kui Zhao
2,
Ligong Chen
3,
Zhaoliang Chen
3,
Yucheng Sun
1 and
Zhendong Guo
1,* 1
Changchun Veterinary Research Institute, Chinese Academy of Agriculture Sciences, Changchun 130122, China
2
College of Animal Medicine, Jilin University, Changchun 130062, China
3
College of Veterinary Medicine, Hebei Agricultural University, Baoding 071000, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Animals 2022, 12(12), 1540; https://doi.org/10.3390/ani12121540
Submission received: 4 May 2022
/
Revised: 8 June 2022
/
Accepted: 8 June 2022
/
Published: 14 June 2022
(This article belongs to the Section Veterinary Clinical Studies)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Microbial aerosols from pig houses can be released into the environment, posing a serious threat to biosafety and public health. At present, there are few studies on the structural characteristics of aerosol bacteria in piggeries at different growth stages. It is important to understand the characteristics of aerosol bacteria in pig houses to solve the problems of air pollution and disease control in pig houses at different growth stages. In this study, bacterial aerosol concentrations and bacterial communities were compared in pig houses at different growth stages in Hebei Province, China. It was found that bacterial concentrations, community richness, and diversity in the air increased with the age of pigs. There are many pathogenic bacteria in the microbial aerosols of piggery. Our study highlights the importance of more comprehensive research and analysis of microbial aerosols in pig houses. Precautions for air pollution should be instituted in pig houses, including wearing masks, rigorous disinfection, and hygiene procedures.
Abstract
With the development of modern pig raising technology, the increasing density of animals in pig houses leads to the accumulation of microbial aerosols in pig houses. It is an important prerequisite to grasp the characteristics of bacteria in aerosols in different pig houses to solve the problems of air pollution and disease prevention and control in different pig houses. This work investigated the effects of growth stages on bacterial aerosol concentrations and bacterial communities in pig houses. Three traditional types of closed pig houses were studied: farrowing (FAR) houses, weaning (WEA) houses, and fattening (FAT) houses. The Andersen six-stage sampler and high-volume air sampler were used to assess the concentrations and size distribution of airborne bacteria, and 16S rRNA gene sequencing was used to identify the bacterial communities. We found that the airborne bacterial concentration, community richness, and diversity index increased with pig age. We found that Acinetobacter, Erysipelothrix, Streptococcus, Moraxella, and Aerococcus in the microbial aerosols of pig houses have the potential risk of causing disease. These differences lead us to believe that disinfection strategies for pig houses should involve a situational focus on environmental aerosol composition on a case-by-case basis.
1. Introduction
With the development of modern pig raising technology, the increasing density of animals in pig houses has resulted in poor air quality in pig houses [1]. The accumulation of microbial aerosols in pig houses poses a serious public health threat [1,2,3]. Microbial aerosols play an important role in the transmission of pathogens [4]. Previous studies have shown that exposure to high concentrations of microbial aerosols is associated with some diseases, such as cardiovascular disease [5,6], atherosclerosis [7], skin lesions [8], and respiratory diseases [9,10]. Pig farmers are at a higher risk of developing respiratory symptoms than those in office or normal human living environments [11,12,13].
Previous studies have shown that the pig gut microbiome varies significantly at different growth stages, while the air microbiota in pig houses is mainly derived from feces [14,15]. However, in different growth stages, how the aerosol bacteria in pig houses change, which of these bacteria are pathogenic bacteria, and the particle size distribution have not been identified. Therefore, it is necessary to study the structure and particle size distribution of microorganisms in pigsties at different stages. Moreover, previous studies have shown that bacteria play a dominant role in microbial aerosols in pig houses [16], and the most dominant phyla in pig houses are Firmicutes and Proteobacteria [17,18]. Potential pathogenic bacteria (such as Acinetobacter, Erysipelothrix, Streptococcus, etc.) in microbial aerosols of pig house bacteria have been widely studied by researchers [19,20,21,22]. Because of technical limitations, culture-based methods are not able to comprehensively analyze the diversity of airborne microorganisms in pig houses. The method of 16S ribosomal RNA (rRNA) gene sequence analysis has been widely used to describe the diversity of bacterial communities [23].
Pigs are an important group of commercial species raised in Hebei Province, China. However, because of the lack of related large-scale pig breeding enterprises, most pig farms are still based on traditional closed breeding, and the structure of the pig house is relatively simple. Furthermore, there is no automatic excrement cleaning or automatic control of the environment in pig house-related equipment. During the winter, there is an increase in respiratory illness among farmworkers, as microbial aerosol concentrations tend to increase with particulate concentrations because of reduced ventilation in pig houses [24,25].
In this study, the concentrations of aerosol bacteria and bacterial communities, as well as bacterial community diversity and community composition of three types of pig houses (FAR, WEA, and FAT), were compared in three typical closed pig farms in Hebei Province. Our data provide a novel basis for the threat to public health of pig house aerosol bacteria.
2. Materials and Methods
2.1. Characteristics of Research Objects
During the winter of 2019, samples were taken from three representative pig farms in Hebei Province, China, all of which belonged to the same swine industry corporation. Pig farms were anonymized according to the company’s requirements. Each pig farm had four types of pig houses: a gestating stable, a farrowing (FAR) stable, a weaning (WEA) stable, and a fattening (FAT) stable. According to the company’s requirements for the safety of pregnant sows, we did not sample gestation houses. The FAR stables consisted of post-farrowing sows and preweaning pig houses, the WEA stables raised postweaning pigs, and the FAT stables raised fattening pigs. For safe production, the FAR stables were positioned in a separate area, while the WEA and FAT stables were located in another. Nursery pigs could be transported from FAR stables to WEA stables through special transport channels.
For the three types of pig houses (breed: Danish landrace), the stocking density was 3.7–4.2 m2/head for parturient sows, 0.5–0.8 m2/head for nursery pigs, and 1–1.5 m2/head for finishing pigs. Indoor structures of pig houses included sheet steel roofs and brick walls using thermal insulation material, with ventilation and a water curtain to maintain a relatively constant temperature and humidity. Each pig house occupied an area of approximately 60 × 15 m2 and was equipped with a double-row structure. Every 5 m, there was a corral in the pig house, and each corral was equipped with a deep pit with slats to collect pig manure, which was cleaned every 15 days. There were no disease outbreaks on any of the farms during our investigation, and samples were collected ten days after fecal cleaning.
2.2. Air Sampling and Cultivation of Isolated Bacteria
An Andersen six-stage sampler (TE-20-800, TISCH, Cleves, OH, USA) was used to identify the size distribution characteristics of culturable bacteria in airborne microorganisms. For each pig house, one sampling point was set in the middle of the passage of the pig house with a height of 0.5 m (the breathing height of pigs) above the ground. The sampling rate was 28.3 L/min for 3 min [2]. The sampler was routinely sterilized with 75% ethanol and dried before sampling. Luria-Bertani (LB) agar medium (Solarbio Science & Technology Co., Ltd., Beijing, China) was used to collect culturable bacteria. The culture plates were then incubated at 37 °C for 24 h. The particle size ranges of the air sampler were 0.65–1.1 μm, 1.1–2.1 μm, 2.1–3.3 μm, 3.3–4.7 μm, 4.7–7.0 μm, and ≥7.0 μm for stages 6, 5, 4, 3, 2, and 1, respectively. Each sample was evaluated in triplicate, and bacterial colonies were counted and calculated according to a previous method to obtain the number of bacterial colonies per cubic meter of air [26].
2.3. Collection of Microbial Aerosols
The method for the collection of microbial aerosols was performed according to our previous study [23]. In brief, a high-volume air sampler (HH02-LS120 of Beijing Huaruihean Technology Co., Ltd., Beijing, China) with 20.32 × 25.4 cm2 Tissuquartz filters (PALL, Port Washington, NY, USA) was used to collect microbial aerosols in the pig house. The sampler was placed in the middle passage of the pig house at a height of 0.5 m above the ground, with a sampling efficiency of 1000 L/min and a sampling time of 12 h [27]. A total of 27 samples were collected from three farms (9 samples per farm), and 3 replications were collected from each pig house on the same farm and mixed together for analysis. Each membrane was cut into an average of 8 pieces with sterile surgical scissors, with a weight error of ±1 mg for each part. Ultrapure water (ST876, Yaji, Shanghai, China) was added to elute the particles. Homogenization was carried out with a homogenizer, and centrifugation was performed at 25,000× g for 10 min at 4 °C. All the filters were sterilized by baking in a muffle furnace at 500 °C for 48 h before sampling. The filters were stored in a mobile refrigerator and transported to the laboratory after collection. Samples were stored at −80 °C until analysis.
2.4. 16S rRNA Gene Sequencing
The total genome DNA extraction method and PCR conditions of microbial aerosol samples were performed as described in previous studies [27,28,29]. A NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA) was used to detect the concentration and quality of extracted DNA (OD260/OD280). PCR amplification was performed after qualified detection. The unqualified samples were extracted again until they passed the test. For bacterial diversity analysis, primers 515F (5′-GTGCCAGCMGCCGCGGTAA-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′) were used to amplify the V3/V4 region of the 16S rRNA gene [30]. For 16S rRNA high-throughput sequencing, the PCR products from each sample were transferred to Beijing Novogene Bioinformatics Technology Co., Ltd. (Novogene, Beijing, China). All original sequences were deposited in GenBank under accession number PRJNA815738.
2.5. Statistical Analysis
QIIME software was employed to remove the poor-quality sequences and obtain high-quality data to control the sequence quality. OTUs (operational taxonomic units) were clustered using UPARSE software (v7.0.1001), which had 97% identity for 16S rRNA sequences [31]. Species annotation was performed on the OTU sequences, and the Mothur method and the SSUrRNA database [32] of SILVA132 were used for species annotation analysis to obtain taxonomic information at each taxonomic level (phylum and genus) and for community composition analysis. The observed-species, Shannon, and Chao1 indices in each sample were calculated with QIIME (version 2.0). Nonmetric multidimensional scaling (NMDS) based on an unweighted Unifrac distance matrix was used to estimate beta diversity. Differences in microbial community structure between groups were also analyzed by principal component analysis (PCA) [33,34]. All data were subjected to one-way analysis of variance (one-way ANOVA) using GraphPad Prism 8 software to determine statistically significant differences (p < 0.05). All analyses were conducted in triplicate and represented at least three separate experiments. The results are presented as means ± standard deviation (SD).
3. Results
3.1. Size Distribution of Culturable Airborne Bacteria
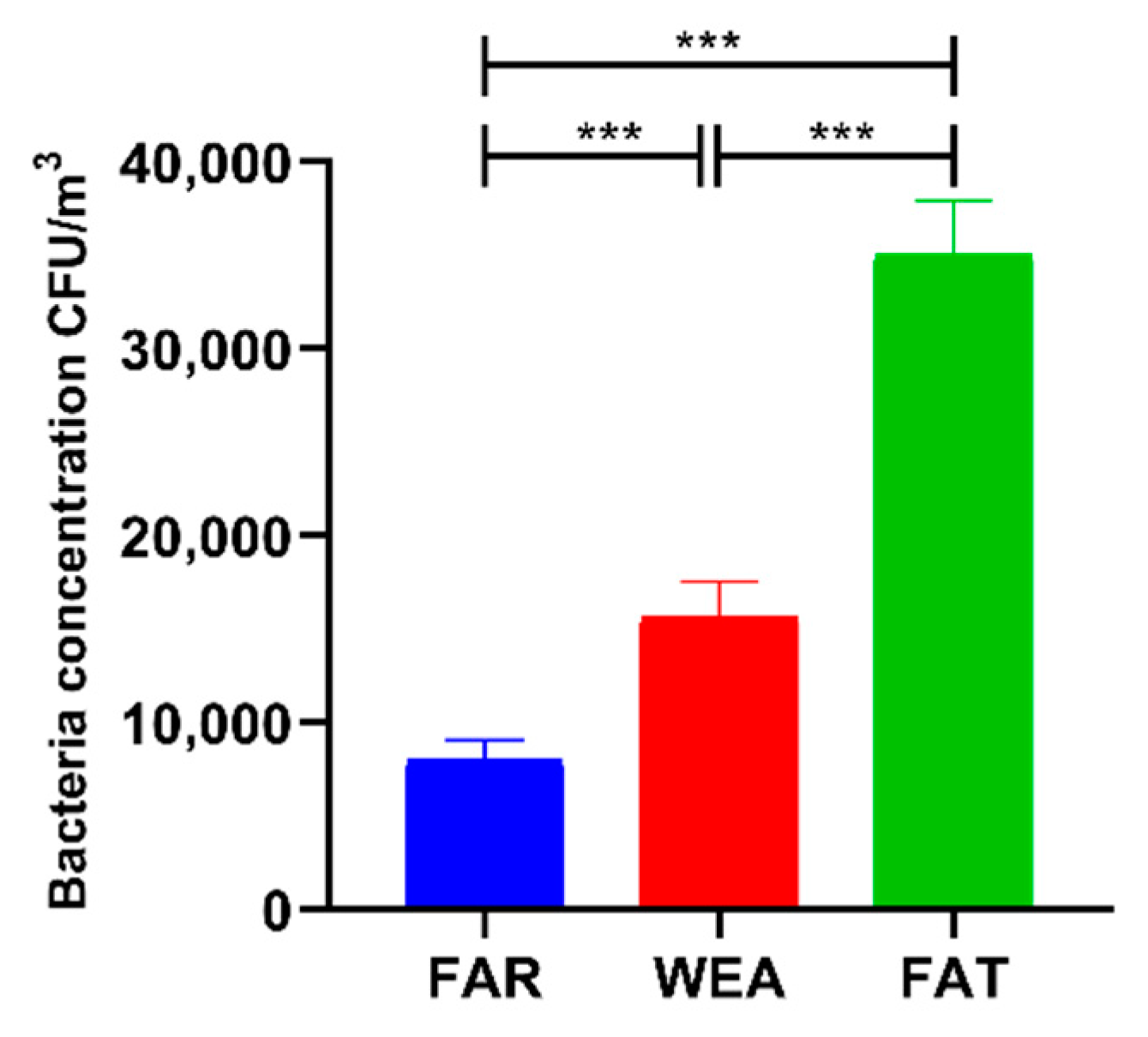
The concentrations of culturable bacteria in the air of the three pig house types (FAR, WEA, and FAT) are shown in Figure 1. The concentrations of airborne bacteria in the FAR, WEA, and FAT stables were 8.092 ± 1.089 × 103 CFU/m3, 15.669 ± 1.899 × 103 CFU/m3, and 35.027 ± 2.859 × 103 CFU/m3, respectively. Bacterial aerosol concentrations were significantly higher in the WEA and FAT stables than in the FAR stables (p < 0.001) and significantly higher in the FAT stables than in the WEA stables (p < 0.001).
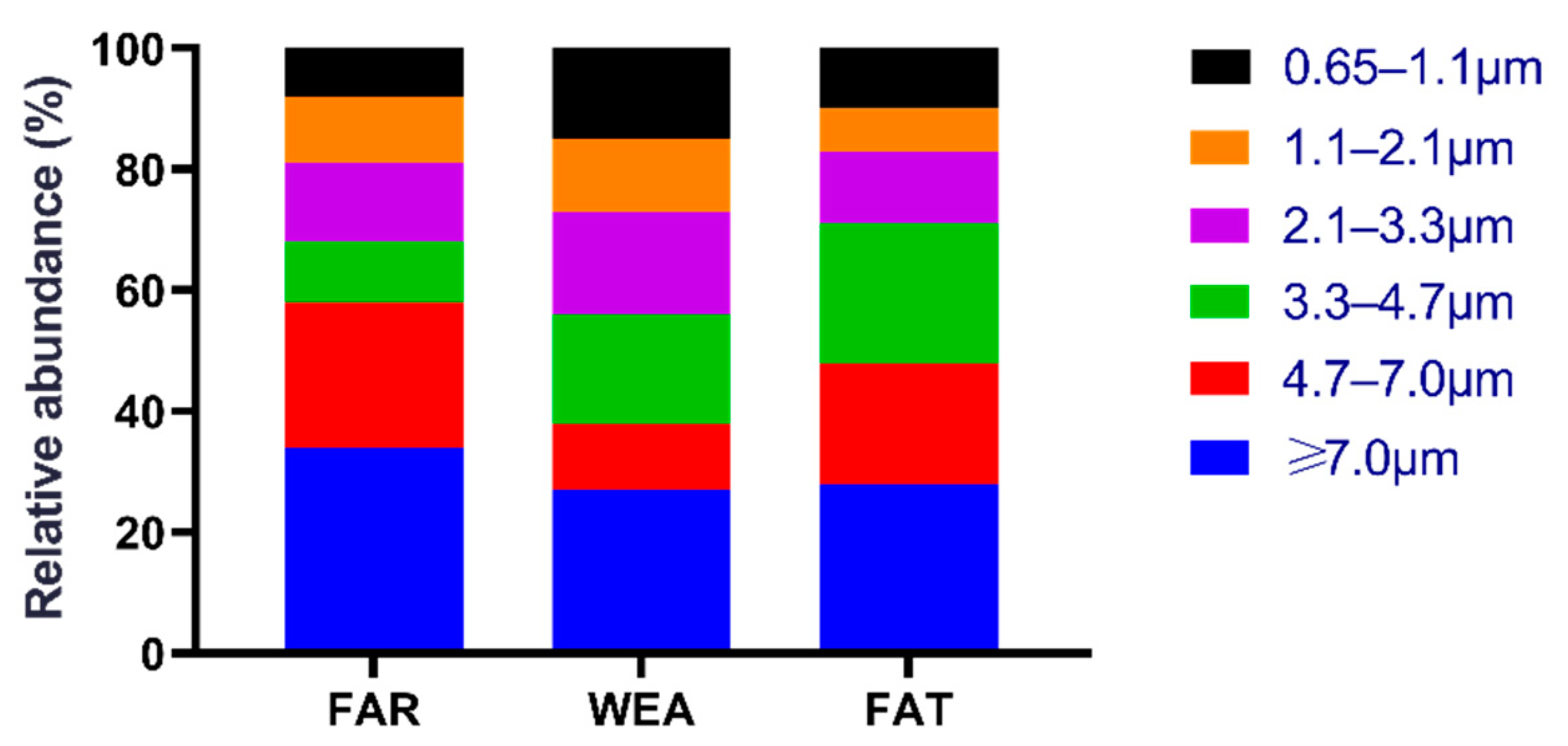
Figure 2 shows the size distribution of culturable airborne bacteria in the three types of pig houses (FAR, WEA, and FAT). In the FAR stables, bacteria with a particle size greater than 3.3 µm (Andersen six-stage sampler, stages 1–3) accounted for 68.2%, while bacteria with a particle size range of 0.6 to 3.3 µm (Andersen six-stage sampler, stages 4–6) accounted for 31.8%. In the WEA stables, a total of 56.2% of airborne bacteria had a particle size greater than 3.3 µm, and 43.8% had a particle size range of 0.6 to 3.3 µm. The majority of bacteria in the FAT stables were found in stage 1 (≥7.0 μm) (28.2%), and stage 5 (1.1–2.1 μm) (7.5%) was the least common.
3.2. Sequencing Analysis
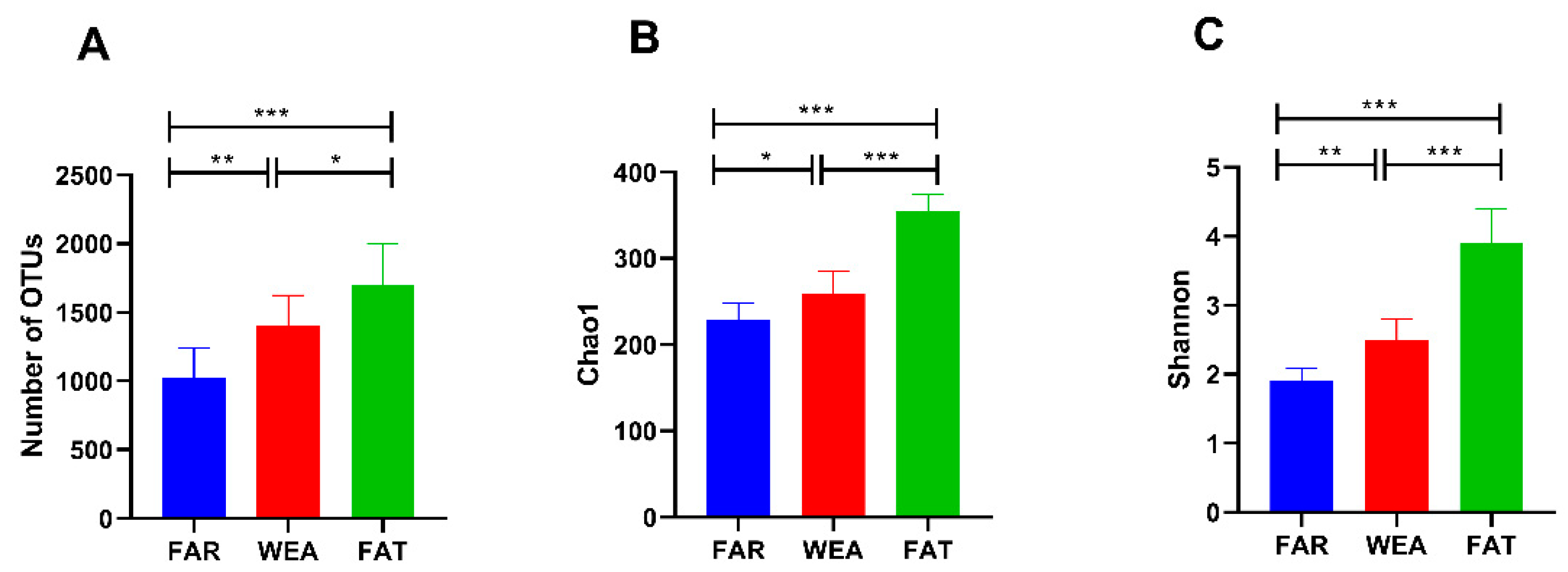
Nine air samples from three pig house types (FAR, WEA, and FAT) were sequenced for the bacterial V3/V4 region. After filtering out the poor-quality sequences, 85,524 effective reads were obtained. The sequences were clustered into 3362 operational taxonomic units (OTUs) with 97% identity, among which the OUTs of the FAR stables were 1.026 ± 0.215 × 103 and the OUTs of the WEA stables were 1.405 ± 0.219 × 103. In the FAT stables, the OUTs were 1.703 ± 0.295 × 103 (Figure 3A). Compared with the FAR stables, the WEA stables showed a significantly increased number of OUTs (p < 0.01). The number of OUTs in the FAT stables was significantly higher than that in the FAR and the WEA stables (p < 0.05). At the same time, the dilution curves of all samples were infinitely close to saturation, indicating that most bacteria in the samples were detected, ensuring the reliability and stability of subsequent analysis.
3.3. Diversity of Airborne Bacterial Communities
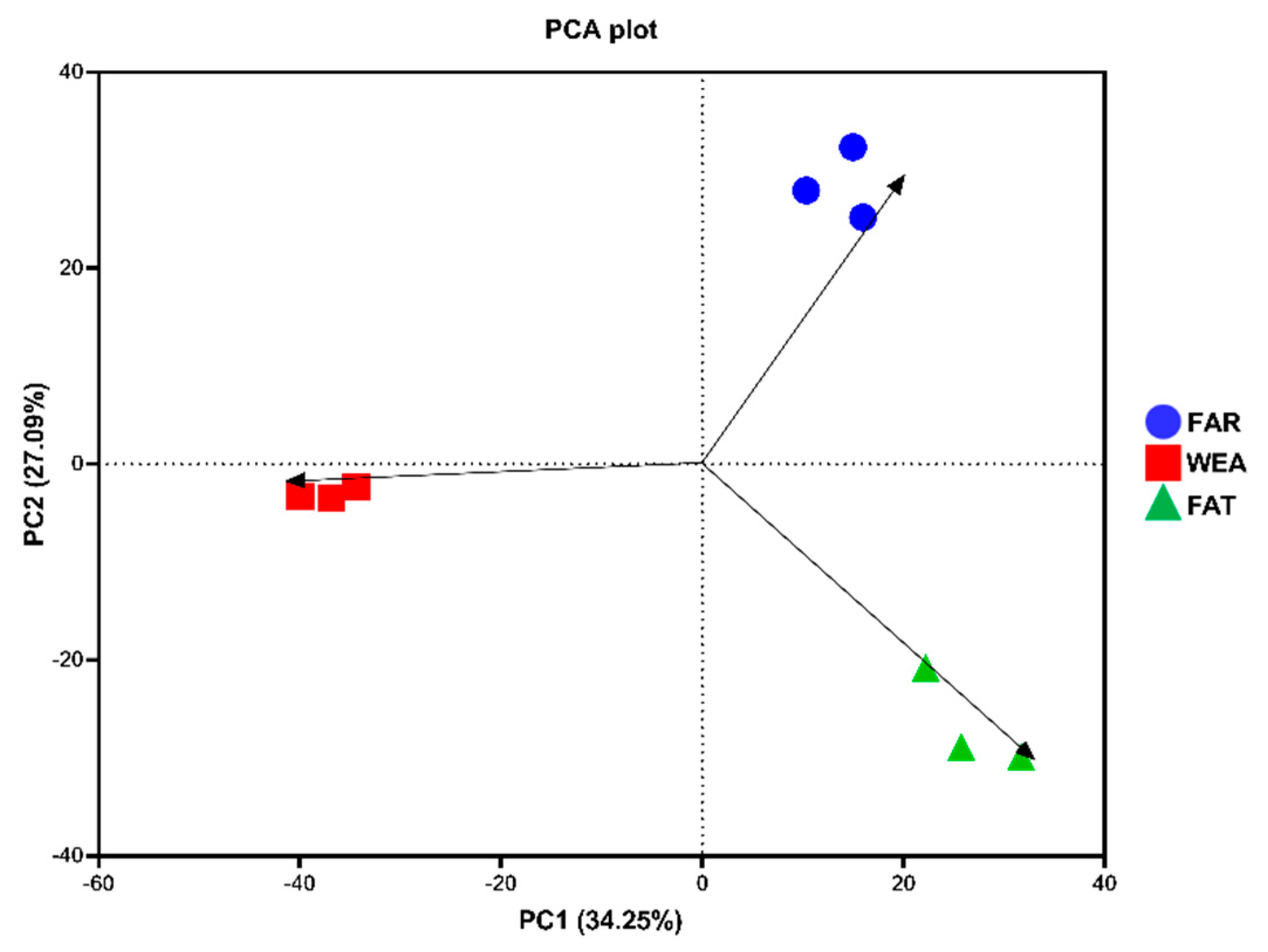
The α-diversity and β-diversity of bacterial communities in the FAR, WEA, and FAT stables are shown in Figure 3B,C and Figure 4. The Chao1 index and Shannon index were evaluated in the α-diversity analysis (Figure 3B,C). Compared with the WEA and FAR stables, the Chao1 index and Shannon index of the FAT stables were significantly higher (p < 0.001) (Figure 3B,C). The Chao1 index and Shannon index of the WEA stables were significantly higher than those of the FAR stables (p < 0.05) (Figure 3B,C). Figure 4 shows the principal component analysis (PCA) of β-diversity. Principal component analysis (PCA) extracted the two axes that best reflected the differences between samples, and each point represented a sample. The closer these points were on the PCA plot, the more similar the community composition of the samples was. The PCA results showed (Figure 4) that there were certain differences in the bacterial communities of the FAR, WEA, and FAT stables. The multiple response permutation programs (MRPP) test showed significant differences among the three types of pig houses (p < 0.05).
3.4. Composition of the Airborne Bacterial Community
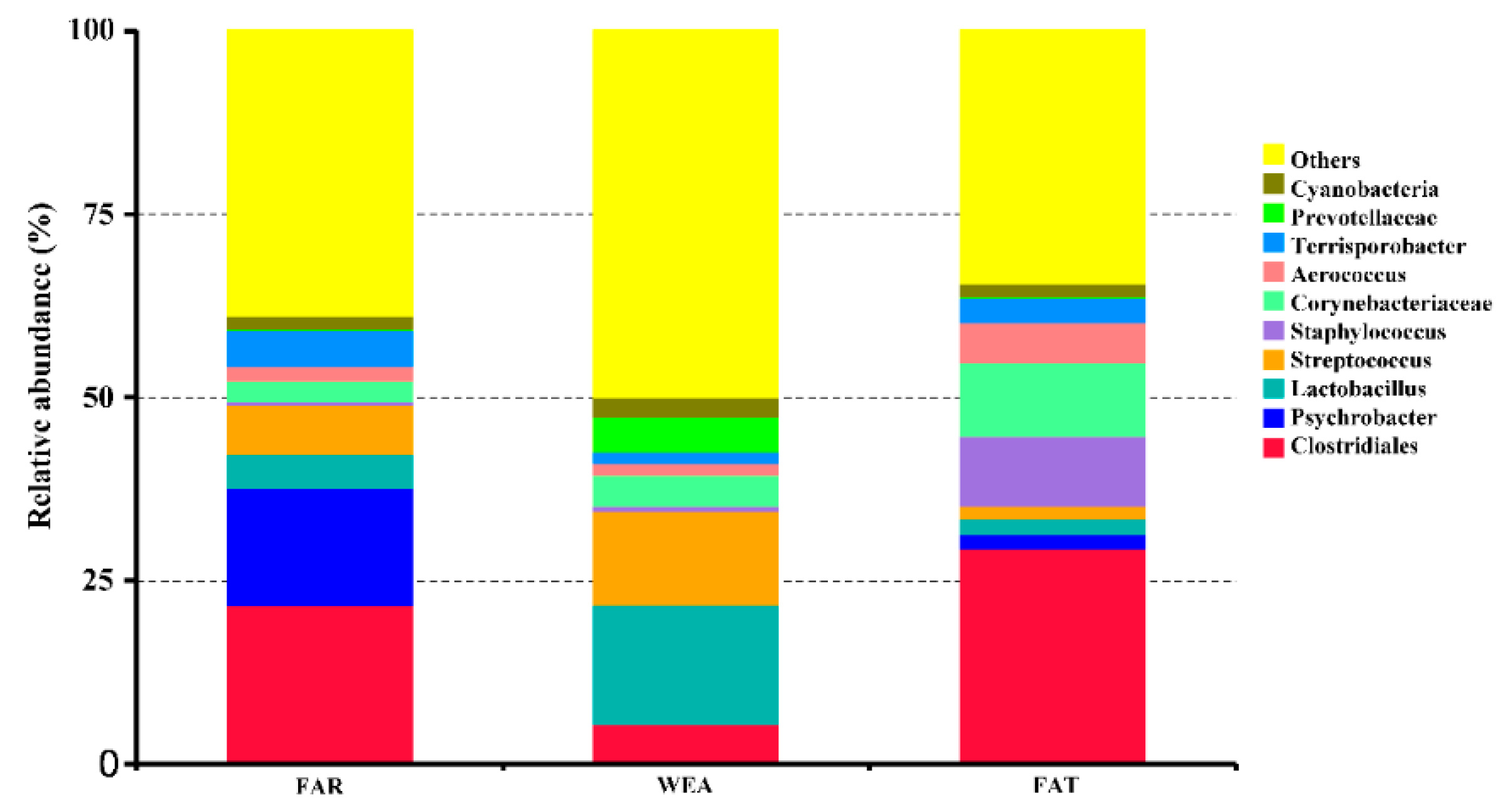
At the phylum level, Firmicutes and Proteobacteria were the most abundant in the bacterial community composition, accounting for 68.86% and 20.23% in the FAR stables, 66.94% and 3.54% in the WEA stables, and 74.95% and 5.4% in the FAT stables, respectively. Figure 5 and Table S1 show the 10 dominant genera in the samples. Our data show that the dominant bacterial genera in the FAR samples were Clostridiales (21.70%), Psychrobacter (16.07%), Streptococcus (6.78%), Terrisporobacter (4.96%), Lactobacillus (4.47%), Corynebacteriaceae (2.71%), Aerococcus (2.03%), Cyanobacteria (1.88%), Staphylococcus (0.58%), and Prevotellaceae (0.06%). The dominant bacterial genera in the WEA samples were Lactobacillus (16.24%), Streptococcus (12.78%), Clostridiales (5.48%), Prevotellaceae (4.73%), Corynebacteriaceae (4.14%), Cyanobacteria (2.54%), Aerococcus (1.70%), Terrisporobacter (1.60%), Staphylococcus (0.72%), and Psychrobacter (0.08%). The dominant bacterial genera in the FAT samples were Clostridiales (29.44%), Corynebacteriaceae (9.97%), Streptococcus (9.56%), Aerococcus (5.49%), Terrisporobacter (3.40%), Lactobacillus (2.12%), Psychrobacter (1.95%), Cyanobacteria (1.87%), Streptococcus (1.74%), and Prevotellaceae (0.12%). Heatmaps for the top 35 genera of all samples are shown in Figure 6. According to the Human Pathogenic Bacteria Directory provided by the Ministry of Health of China (MOHC), the potentially pathogenic bacteria in the FAR stables were Acinetobacter, Streptococcus, and Erysipelothrix; the potentially pathogenic bacteria in the WEA stables were Streptococcus.
4. Discussion
In this study, three types of pig houses (FAR, WEA, and FAT) of three typical closed pig farms in Hebei Province were investigated. The aerosol bacterial concentration and bacterial community, as well as the bacterial community diversity and composition of the three types of pig houses, were compared to provide a data basis and reference for healthy pig breeding and the threat of microbial aerosols to public health in pig houses.
Previous studies have shown that bacteria dominate microbial aerosols in pig houses [16], so this study focused on bacterial microbial aerosols. Our study shows that airborne bacterial concentrations in pig houses increase as pigs age. The concentrations of culturable airborne bacteria were the lowest in FAR and significantly higher in WEA and FAT (p < 0.001). This may be due to the increased activity and excretion of pigs as they grow and increased feeding density, resulting in increased concentrations of bacterial aerosols. The numbers of culturable bacteria measured in the three types of stables (FAR, WEA, and FAT) were 8.092 ± 1.089 × 103 CFU/m3, 15.669 ± 1.899 × 103 CFU/m3, and 35.027 ± 2.859 × 103 CFU/m3, respectively. According to studies, the culturable bacterial concentration in the WEA stables of Jiangsu, China, was 3.460× 103 CFU/m3 (1), and the culturable bacterial concentrations in the air of three types of pig houses (FAR, WEA, and FAT) in Korea were 2.322 ± 1.352 × 103 CFU/m3, 6.124 ± 1.527 × 103 CFU/m3, and 12.470 ± 4.869 × 103 CFU/m3 [35]. In our investigation, the concentration of bacterial microbial aerosols was greater than that in previous studies. The pigs, the construction of the barns, the methods of management, and the indoor climate may all have a role in the fluctuation in microbiological concentrations of bacterial aerosols [36].
Different particle sizes of bacterial aerosols enter the respiratory system in different locations. Particles of 4.7–7 µm in diameter can enter the nasal cavity and upper respiratory tract; particles of 2.1–4.7 µm can be deposited in the bronchi, causing asthma and other diseases; however, those less than 2 µm in diameter can penetrate the alveoli and enter the respiratory tract, causing alveolar inflammation and other diseases. The smallest fraction of the fine particles may have a disproportionately large role. Although fine particles are only a small proportion, they still pose a serious threat to the respiratory health of pigs. In the FAT stables, 71.9% of airborne bacteria had a particle size larger than 3.3 µm, and 28.1% had a particle size ranging from 0.6 to 3.3 µm. Aerosols with a particle size of less than 3.3 µm are defined as fine particles, and those larger than 3.3 µm are defined as coarse particles. Culturable bacteria in the air predominate in coarse particles, but not in fine particles, similar to our results [2]. Previous research showed that coarse particles accounted for 64% and fine particles accounted for 36% of particles in the FAR stables. In the WEA stables, coarse particles accounted for 60%, and fine particles accounted for 40% of the total particles. The proportions of coarse particles and fine particles were 63% and 37%, respectively [35]. The smallest fraction of the fine particles may have a disproportionately large role. Although fine particles are only a small proportion, they still pose a serious threat to the respiratory health of pigs. The results were similar to those in the FAR and WEA stables, but the proportion of coarse particles in the FAT stables was higher, which may have been caused by various factors, such as pig breed, stocking density, climate, and the structure of the barns.
We found that the bacterial community α-diversity index of microbial aerosols increased gradually with the age of pigs. The Chao1 index and Shannon index of the FAT stables were significantly higher than those of the FAR and WEA stables (p < 0.001). PCA showed that the composition of the bacterial community also changed significantly (p < 0.05). As animals grow, their activity and excretion increase and their metabolic levels change, which could cause remarkable changes in the microbial community structure in the air of feedlots [37]. Firmicutes and Proteobacteria were the dominant phyla in the three types of pig houses, which is consistent with previous studies [17,18,20]. Firmicutes are the most abundant phylum in the intestinal microbiome of pigs at all growth stages [38], suggesting that the dominant bacteria in the air of the pig house may originate from feces [39]. Proteobacteria is the largest phylum of bacteria, including many pathogenic bacteria, such as Escherichia coli, Salmonella, Vibrio cholera, Helicobacter pylori, and other well-known species [40,41,42,43], while some members of Proteobacteria play an important ecological role in various environments [44].
According to the human pathogenic bacteria directory provided by the Ministry of Health of China (MOHC), the potentially pathogenic bacteria in the FAR stables were Acinetobacter, Streptococcus, and Erysipelothrix; those in the WEA stables were Streptococcus. Acinetobacter widely exists in the environment, and studies have shown that the mortality rate of patients with underlying diseases infected with Acinetobacter is much higher than that of other patients [45,46,47]. Meanwhile, the number of Acinetobacter strains resistant to multiple antibiotics has increased significantly in recent years, suggesting potential biosafety risks [48]. Erysipelothrix is extensively distributed in the environment and may survive for a long period [49,50]. Erysipelothrix is a pathogen associated with many occupational diseases, and its pathogenesis may be related to hyaluronidase and neuraminidase produced by bacteria [19]. Streptococcus suis, an important zoonotic pathogen in the genus Streptococcus, lives naturally in the respiratory system of pigs, particularly the upper respiratory tract, and is found in nearly every country [22]. Previous studies have reported that Streptococcus has been found in nasal swabs of pig workers [51]. Thus, the prevention and control of Streptococcus deserve wider attention. Moraxella in WEA and Aerococcus in FAT, in addition to the list of human pathogenic bacteria, have been identified as potentially pathogenic concerns that should be monitored. Moraxella catarrhalis is a pathogenic strain of Moraxella that has been found in the human respiratory system [52]. In the respiratory system, Moraxella lacunata and Moraxella nonliquefaciens are also opportunistic [53]. Aerococcus has been recovered from clinical samples of pigs in a growing number of investigations, indicating that it is pathogenic [39,54,55]. Previous research has found that microbial aerosols in pig houses include many Aerococcus, and pig employees working in an environment with many Aerococcus are at risk of becoming sick [1]. The source of the pathogenic bacteria of this genus is unclear, but pathogenic bacteria are likely to spread through pig houses to the outside world, and pig employees are likely to carry these infections on their clothes or in their lungs, resulting in a wide range of biological safety hazards and concerns [2,46,56].
5. Conclusions
This study showed that with the increase in weekly pig age, the concentration of airborne bacteria, the species, and the diversity index of the bacterial community in pig houses has also increased gradually. The pathogenic bacteria Acinetobacter, Erysipelothrix, Streptococcus, Moraxella, and Aerococcus were detected in three types of pig houses. These bacteria with potential risk to pig and human health should be monitored. The number of disinfections should be increased to 2–3 times a week or every other day. A disinfectant should be replaced after 2–3 weeks of continuous use for another disinfectant to avoid pathogenic microorganisms producing drug resistance and reduce the disinfection effect. Spray disinfection should be used to kill airborne microorganisms in piggeries to make the piggeries cleaner. These measures may help optimize the piggery environment. From the point of view of personnel, farmers should wear masks, hats, and overalls to combat the effects of air pollution when working on farms. In this study, we emphasize the necessity of long-term detection of microbial aerosols in pig houses and the importance of more comprehensive research and analysis of microbial aerosols in livestock and poultry houses.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/ani12121540/s1, Table S1: Relative abundances of the airborne bacterial genera (%).
Author Contributions
Conceptualization, Z.G.; data curation, H.C., K.Z., L.C., Z.C. and Y.S.; funding acquisition, J.L. and Z.G.; investigation, H.C. and C.Z.; project administration, Z.G.; resources, S.D.; software, Z.C.; visualization, H.C.; writing—original draft, H.C. and C.Z. All authors contributed to the article and approved the submitted version. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Key Research and Development Program of China (2016YFD0500505) and the Key Research and Development Program of Hebei Province (18227517D and 20326622D). The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Institutional Review Board Statement
All the experiments were approved by the relevant regulatory agency of the Animal Ethical Committee of Changchun Veterinary Research Institute (document number of approval: SCXK201905067).
Informed Consent Statement
Not applicable.
Data Availability Statement
The study’s original contributions are included in the article; further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- White, J.K.; Nielsen, J.L.; Madsen, A.M. Microbial Species and Biodiversity in Settling Dust within and between Pig Farms. Environ. Res. 2019, 171, 558–567. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Huang, K.; Liu, J.; Jin, X.; Li, C. Distribution Characteristics of Bioaerosols inside Pig Houses and the Respiratory Tract of Pigs. Ecotoxicol. Environ. Saf. 2021, 212, 112006. [Google Scholar] [CrossRef] [PubMed]
- Weese, J.S.; Slifierz, M.; Jalali, M.; Friendship, R. Evaluation of the Nasal Microbiota in Slaughter-Age Pigs and the Impact on Nasal Methicillin-Resistant Staphylococcus Aureus (Mrsa) Carriage. BMC Vet. Res. 2014, 10, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlesinger, R.B.; Cassee, F. Atmospheric Secondary Inorganic Particulate Matter: The Toxicological Perspective as a Basis for Health Effects Risk Assessment. Inhal Toxicol. 2003, 15, 197–235. [Google Scholar] [CrossRef] [PubMed]
- Yin, P.; Guo, J.; Wang, L.; Fan, W.; Lu, F.; Guo, M.; Moreno, S.B.; Wang, Y.; Wang, H.; Zhou, M.; et al. Higher Risk of Cardiovascular Disease Associated with Smaller Size-Fractioned Particulate Matter. J. Environ. Sci. Technol. Lett. 2020, 7, 95–101. [Google Scholar] [CrossRef]
- Elliott, E.G.; Laden, F.; James, P.; Rimm, E.B.; Rexrode, K.M.; Hart, J.E. Interaction between Long-Term Exposure to Fine Particulate Matter and Physical Activity, and Risk of Cardiovascular Disease and Overall Mortality in U.S. Women. Environ. Health Perspect. 2020, 128, 127012. [Google Scholar] [CrossRef] [PubMed]
- Holme, S.A.N.; Sigsgaard, T.; Holme, J.A.; Holst, G.J. Effects of Particulate Matter on Atherosclerosis: A Link Via High-Density Lipoprotein (Hdl) Functionality? Part. Fibre Toxicol 2020, 17, 36. [Google Scholar] [CrossRef]
- Dijkhoff, I.M.; Drasler, B.; Karakocak, B.B.; Petri-Fink, A.; Valacchi, G.; Eeman, M.; Rothen-Rutishauser, B. Impact of Airborne Particulate Matter on Skin: A Systematic Review from Epidemiology to in Vitro Studies. Part. Fibre Toxicol. 2020, 17, 35. [Google Scholar] [CrossRef]
- Zhang, Z.; Chai, P.; Wang, J.; Ye, Z.; Shen, P.; Lu, H.; Jin, M.; Gu, M.; Li, D.; Lin, H.; et al. Association of Particulate Matter Air Pollution and Hospital Visits for Respiratory Diseases: A Time-Series Study from China. Environ. Sci. Pollut. Res. Int. 2019, 26, 12280–12287. [Google Scholar] [CrossRef]
- Losacco, C.; Perillo, A. Particulate Matter Air Pollution and Respiratory Impact on Humans and Animals. Environ. Sci. Pollut. Res. 2018, 25, 33901–33910. [Google Scholar] [CrossRef]
- Beltrán-Alcrudo, D.; Kukielka, E.A.; De Groot, N.; Dietze, K.; Sokhadze, M.; Martínez-López, B. Descriptive and Multivariate Analysis of the Pig Sector in Georgia and Its Implications for Disease Transmission. PLoS ONE 2018, 13, e0202800. [Google Scholar] [CrossRef] [PubMed]
- Hoppin, J.A.; Umbach, D.M.; Long, S.; Rinsky, J.L.; Henneberger, P.K.; Salo, P.M.; Zeldin, D.; London, S.; Alavanja, M.C.R.; Blair, A.; et al. Respiratory Disease in United States Farmers. Occup. Environ. Med. 2014, 71, 484–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Park, T.; Yu, Z. Metagenomic Investigation of Gastrointestinal Microbiome in Cattle. Asian-Australas, J. Anim. Sci. 2017, 30, 1515–1528. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Estellé, J.; Kiilerich, P.; Ramayo-Caldas, Y.; Xia, Z.; Feng, Q.; Liang, S.; Pedersen, A.O.; Kjeldsen, N.J.; Liu, C.; et al. A Reference Gene Catalogue of the Pig Gut Microbiome. Nat. Microbiol. 2016, 1, 16161. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Wang, X.-Q.; Wang, J.; Zhao, J. Life-Long Dynamics of the Swine Gut Microbiome and Their Implications in Probiotics Development and Food Safety. Gut Microbes 2020, 11, 1824–1832. [Google Scholar] [CrossRef]
- Tang, Q.; Huang, K.; Liu, J.; Shen, D.; Dai, P.; Li, Y.; Li, C. Seasonal Variations of Microbial Assemblage in Fine Particulate Matter from a Nursery Pig House. Sci. Total Environ. 2019, 708, 134921. [Google Scholar] [CrossRef]
- Hong, P.-Y.; Li, X.; Yang, X.; Shinkai, T.; Zhang, Y.; Wang, X.; Mackie, R.I. Monitoring Airborne Biotic Contaminants in the Indoor Environment of Pig and Poultry Confinement Buildings. Environ. Microbiol. 2012, 14, 1420–1431. [Google Scholar] [CrossRef]
- Kumari, P.; Choi, H.L. Seasonal Variability in Airborne Biotic Contaminants in Swine Confinement Buildings. PLoS ONE 2014, 9, e112897. [Google Scholar] [CrossRef]
- Reboli, A.C.; Farrar, W.E. Erysipelothrix Rhusiopathiae: An Occupational Pathogen. Clin. Microbiol. Rev. 1989, 2, 354–359. [Google Scholar] [CrossRef]
- Kraemer, J.G.; Aebi, S.; Oppliger, A.; Hilty, M. The Indoor-Air Microbiota of Pig Farms Drives the Composition of the Pig Farmers’ Nasal Microbiota in a Season-Dependent and Farm-Specific Manner. Appl. Environ. Microbiol. 2019, 85, e03038-18. [Google Scholar] [CrossRef] [Green Version]
- Foster, A.P. Staphylococcal Skin Disease in Livestock. Vet. Dermatol. 2012, 23, 342–351.e63. [Google Scholar] [CrossRef] [PubMed]
- Goyette-Desjardins, G.; Auger, J.P.; Xu, J.; Segura, M.; Gottschalk, M. Streptococcus Suis, an Important Pig Pathogen and Emerging Zoonotic Agent-an Update on the Worldwide Distribution Based on Serotyping and Sequence Typing. Emerg. Microbes Infect. 2014, 3, e45. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Wang, Z.; Qian, L.A.; Zhao, Z.; Zhang, C.; Fu, Y.; Li, J.; Zhang, C.; Lu, B.; Qian, J. Biological and Chemical Compositions of Atmospheric Particulate Matter During Hazardous Haze Days in Beijing. Environ. Sci. Pollut. Res. 2018, 25, 34540–34549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bønløkke, J.H.; Mériaux, A.; Duchaine, C.; Godbout, S.; Cormier, Y. Seasonal Variations in Work-Related Health Effects in Swine Farm Workers. Ann. Agric. Environ. Med. 2009, 16, 43–52. [Google Scholar] [PubMed]
- Kumari, P.; Woo, C.; Yamamoto, N.; Choi, H.-L. Variations in Abundance, Diversity and Community Composition of Airborne Fungi in Swine Houses across Seasons. Sci. Rep. 2016, 6, 37929. [Google Scholar] [CrossRef]
- Jiang, L.; Zhang, J.; Tang, J.; Li, M.; Zhao, X.; Zhu, H.; Yu, X.; Li, Y.; Feng, T.; Zhang, X. Analyses of Aerosol Concentrations and Bacterial Community Structures for Closed Cage Broiler Houses at Different Broiler Growth Stages in Winter. J. Food Prot. 2018, 81, 1557–1564. [Google Scholar] [CrossRef]
- Yan, H.; Li, Y.; Zhang, Y.; Zhang, H.; Guo, Z.; Liu, J. Deciphering of Microbial Diversity and Antibiotic Resistome of Bioaerosols in Swine Confinement Buildings. Sci. Total Environ. 2021, 781, 147095. [Google Scholar] [CrossRef]
- Cui, B.; Su, D.; Li, W.; She, X.; Zhang, M.; Wang, R.; Zhai, Q. Effects of Chronic Noise Exposure on the Microbiome-Gut-Brain Axis in Senescence-Accelerated Prone Mice: Implications for Alzheimer’s Disease. J. Neuroinflamm. 2018, 15, 190. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, J.; Wang, H.; Huang, Y.; Qi, M.; Liao, S.; Bin, P.; Yin, Y. Effects of GABA Supplementation on Intestinal SIgA Secretion and Gut Microbiota in the Healthy and ETEC-Infected Weanling Piglets. Mediat. Inflamm. 2020, 2020, 7368483. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian Classifier for Rapid Assignment of Rrna Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, K.S.; Kirkegaard, R.H.; Karst, S.M.; Albertsen, M. Ampvis2: An R Package to Analyse and Visualise 16s Rrna Amplicon Data. BioRxiv 2018, 299537. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Rubio, V. Ggplot2-Elegant Graphics for Data Analysis. J. Stat. Softw. 2017, 77, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.Y.; Ko, H.J. Indoor Distribution Characteristics of Airborne Bacteria in Pig Buildings as Influenced by Season and Housing Type. Asian-Australas. J. Anim. Sci. 2019, 32, 742–747. [Google Scholar] [CrossRef]
- Zhao, Y.; Aarnink, A.J.A.; de Jong, M.; Koerkamp, P.W.G.G. Airborne Microorganisms From Livestock Production Systems and Their Relation to Dust. Crit. Rev. Environ. Sci. Technol. 2014, 44, 1071–1128. [Google Scholar] [CrossRef]
- Liu, Y.W.; Zhang, X.; Bai, F.J.; Shi, Y.X.; Wang, B.; Liu, N.; Shi, R.Y.; Shen, J.; Huang, X.L. Dynamic Study on the Species and Concentration of Fungi in Aerosol of Henhouse. Chin. J. Vet. Sci. 2017, 37, 2095–2099. [Google Scholar]
- Noh, H.S.; Ingale, S.L.; Lee, S.H.; Kim, K.H.; Kwon, I.K.; Kim, Y.H.; Chae, B.J. Effects of citrus pulp, fish by-product and Bacillus subtilis fermentation biomass on growth performance, nutrient digestibility, and fecal microflora of weanling pigs. J. Anim. Sci. Technol. 2014, 56, 10. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.-W.; Park, J.; Jeong, H.; Kim, M. Evaluation of the microbiome composition in particulate matter inside and outside of pig houses. J. Anim. Sci. Technol. 2021, 63, 640–650. [Google Scholar] [CrossRef]
- Waite, D.W.; Vanwonterghem, I.; Rinke, C.; Parks, D.H.; Zhang, Y.; Takai, K.; Sievert, S.M.; Simon, J.; Campbell, B.J.; Hanson, T.E.; et al. Comparative Genomic Analysis of the Class Epsilonproteobacteria and Proposed Reclassification to Epsilonbacteraeota (Phyl. Nov.). Front. Microbiol. 2017, 8, 682. [Google Scholar] [CrossRef]
- Bublitz, D.C.; Wright, P.C.; Bodager, J.R.; Rasambainarivo, F.T.; Bliska, J.B.; Gillespie, T.R. Epidemiology of Pathogenic Enterobacteria in Humans, Livestock, and Peridomestic Rodents in Rural Madagascar. PLoS ONE 2014, 9, e101456. [Google Scholar] [CrossRef] [PubMed]
- Moura-Alves, M.; Carvalho, M.; Ribeiro, D.H.B.; Barbosa, J.; Silveira, L.; Pista, Â.; Pinto, H.P.; Saraiva, C.; Teixeira, P.; Esteves, A. Hygiene indicators and Salmonella sp. on swine carcass surfaces from two slaughterhouses in northern Portugal. J. Food Prot. 2022. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.L.; Zhou, J.Y.; Liang, S.J.; Wang, X.Q. Impaired Intestinal Stem Cell Activity in Etec Infection: Enterotoxins, Cyclic Nucleotides, and Wnt Signaling. Arch. Toxicol. 2022, 96, 1213–1225. [Google Scholar] [CrossRef]
- Nakagawa, S.; Takaki, Y. Nonpathogenic Epsilonproteobacteria. eLS 2009. [Google Scholar] [CrossRef]
- Tringe, S.; Zhang, T.; Liu, X.; Yu, Y.; Lee, W.H.; Yap, J.; Yao, F.; Suan, S.T.; Ing, S.K.; Haynes, M.; et al. The Airborne Metagenome in an Indoor Urban Environment. PLoS ONE 2008, 3, e1862. [Google Scholar] [CrossRef]
- Rintala, H.; Pitkäranta, M.; Toivola, M.; Paulin, L.; Nevalainen, A. Diversity and Seasonal Dynamics of Bacterial Community in Indoor Environment. BMC Microbiol. 2008, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Hahm, C.; Chung, H.-S.; Lee, M. Whole-genome sequencing for the characterization of resistance mechanisms and epidemiology of colistin-resistant Acinetobacter baumannii. PLoS ONE 2022, 17, e0264335. [Google Scholar] [CrossRef]
- Liu, C.; Yoon, E.-J.; Kim, D.; Shin, J.H.; Shin, J.H.; Shin, K.S.; Kim, Y.A.; Uh, Y.; Kim, H.S.; Kim, Y.R.; et al. Antimicrobial resistance in South Korea: A report from the Korean global antimicrobial resistance surveillance system (Kor-GLASS) for 2017. J. Infect. Chemother. 2019, 25, 845–859. [Google Scholar] [CrossRef]
- Janßen, T.; Voss, M.; Kühl, M.; Semmler, T.; Philipp, H.-C.; Ewers, C. A combinational approach of multilocus sequence typing and other molecular typing methods in unravelling the epidemiology of Erysipelothrix rhusiopathiae strains from poultry and mammals. Veter. Res. 2015, 46, 84. [Google Scholar] [CrossRef] [Green Version]
- Forde, T.L.; Ratheesh, N.K.; Harvey, W.T.; Thomson, J.R.; Williamson, S.; Biek, R.; Opriessnig, T. Genomic and Immunogenic Protein Diversity of Erysipelothrix rhusiopathiae Isolated From Pigs in Great Britain: Implications for Vaccine Protection. Front. Microbiol. 2020, 11, 418. [Google Scholar] [CrossRef] [Green Version]
- Bonifait, L.; Veillette, M.; Létourneau, V.; Grenier, D.; Duchaine, C. Detection of Streptococcus suis in Bioaerosols of Swine Confinement Buildings. Appl. Environ. Microbiol. 2014, 80, 3296–3304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, T.F.; Parameswaran, G.I. Moraxella catarrhalis, a Human Respiratory Tract Pathogen. Clin. Infect. Dis. 2009, 49, 124–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, D.R.; Band, J.D.; Thornsberry, C.; Hollis, D.G.; Weaver, R.E. Infections caused by Moraxella, Moraxella urethralis, Moraxella-like groups M-5 and M-6, and Kingella kingae in the United States, 1953–1980. Rev. Infect. Dis. 1990, 12, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Vela, A.I.; García, N.; Latre, M.V.; Casamayor, A.; Sánchez-Porro, C.; Briones, V.; Ventosa, A.; Domínguez, L.; Fernández-Garayzábal, J.F. Aerococcus Suis Sp. Nov., Isolated from Clinical Specimens from Swine. Int. J. Syst. Evol. Microbiol. 2007, 57, 1291–1294. [Google Scholar] [CrossRef]
- Moreno, L.Z.; Matajira, C.E.; Gomes, V.T.; Silva, A.P.S.; Mesquita, R.E.; Christ, A.P.G.; Sato, M.I.Z.; Moreno, A.M. Molecular and antibiotic susceptibility characterization of Aerococcus viridans isolated from porcine urinary infection. Veter. Microbiol. 2016, 184, 7–10. [Google Scholar] [CrossRef]
- Angen, Ø.; Feld, L.; Larsen, J.; Rostgaard, K.; Skov, R.; Madsen, A.M.; Larsen, A.R. Transmission of Methicillin-Resistant Staphylococcus aureus to Human Volunteers Visiting a Swine Farm. Appl. Environ. Microbiol. 2017, 83, e01489-17. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Concentrations of culturable airborne bacteria in different types of pig houses using an Andersen sampler (data presented as means ± SD. *** p < 0.001). FAR, farrowing houses; WEA, weaning houses; FAT, fattening houses.
Figure 1.
Concentrations of culturable airborne bacteria in different types of pig houses using an Andersen sampler (data presented as means ± SD. *** p < 0.001). FAR, farrowing houses; WEA, weaning houses; FAT, fattening houses.

Figure 2.
The distribution ratio of culturable airborne bacteria in different types of pig houses. FAR, farrowing houses; WEA, weaning houses; FAT, fattening houses.
Figure 2.
The distribution ratio of culturable airborne bacteria in different types of pig houses. FAR, farrowing houses; WEA, weaning houses; FAT, fattening houses.

Figure 3.
Alpha diversity of the bacterial community in different types of pig houses. (A) Observed species; (B) Chao1 index; (C) Shannon index (data presented as the means ± SDs. * p < 0.05, ** p < 0.01, *** p < 0.001). FAR, farrowing houses; WEA, weaning houses; FAT, fattening houses.
Figure 3.
Alpha diversity of the bacterial community in different types of pig houses. (A) Observed species; (B) Chao1 index; (C) Shannon index (data presented as the means ± SDs. * p < 0.05, ** p < 0.01, *** p < 0.001). FAR, farrowing houses; WEA, weaning houses; FAT, fattening houses.

Figure 4.
Principal component analysis (PCA) of samples from different types of pig houses. FAR, farrowing houses; WEA, weaning houses; FAT, fattening houses.
Figure 4.
Principal component analysis (PCA) of samples from different types of pig houses. FAR, farrowing houses; WEA, weaning houses; FAT, fattening houses.

Figure 5.
Relative abundances of the airborne bacterial genera (%).

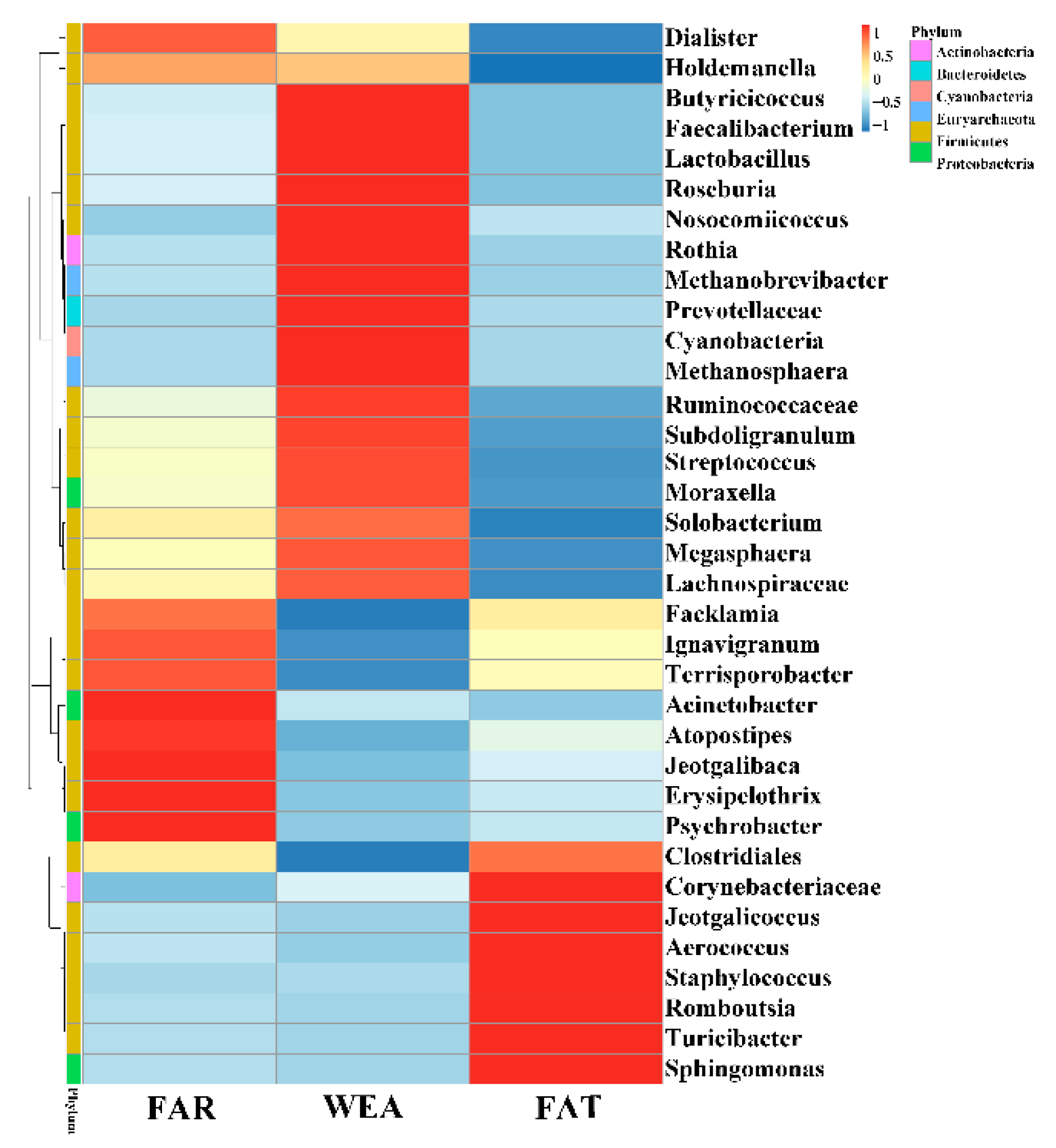
Figure 6.
Heatmap of airborne bacterial relative abundance at the genus level. FAR, farrowing houses; WEA, weaning houses; FAT, fattening houses.
Figure 6.
Heatmap of airborne bacterial relative abundance at the genus level. FAR, farrowing houses; WEA, weaning houses; FAT, fattening houses.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cui, H.; Zhang, C.; Liu, J.; Dong, S.; Zhao, K.; Chen, L.; Chen, Z.; Sun, Y.; Guo, Z. The Distribution Characteristics of Aerosol Bacteria in Different Types of Pig Houses. Animals 2022, 12, 1540. https://doi.org/10.3390/ani12121540
AMA Style
Cui H, Zhang C, Liu J, Dong S, Zhao K, Chen L, Chen Z, Sun Y, Guo Z. The Distribution Characteristics of Aerosol Bacteria in Different Types of Pig Houses. Animals. 2022; 12(12):1540. https://doi.org/10.3390/ani12121540
Chicago/Turabian StyleCui, Huan, Cheng Zhang, Juxiang Liu, Shishan Dong, Kui Zhao, Ligong Chen, Zhaoliang Chen, Yucheng Sun, and Zhendong Guo. 2022. "The Distribution Characteristics of Aerosol Bacteria in Different Types of Pig Houses" Animals 12, no. 12: 1540. https://doi.org/10.3390/ani12121540
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.