
Influence of Single Layer Centrifugation with Canicoll on Semen Freezability in Dogs

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
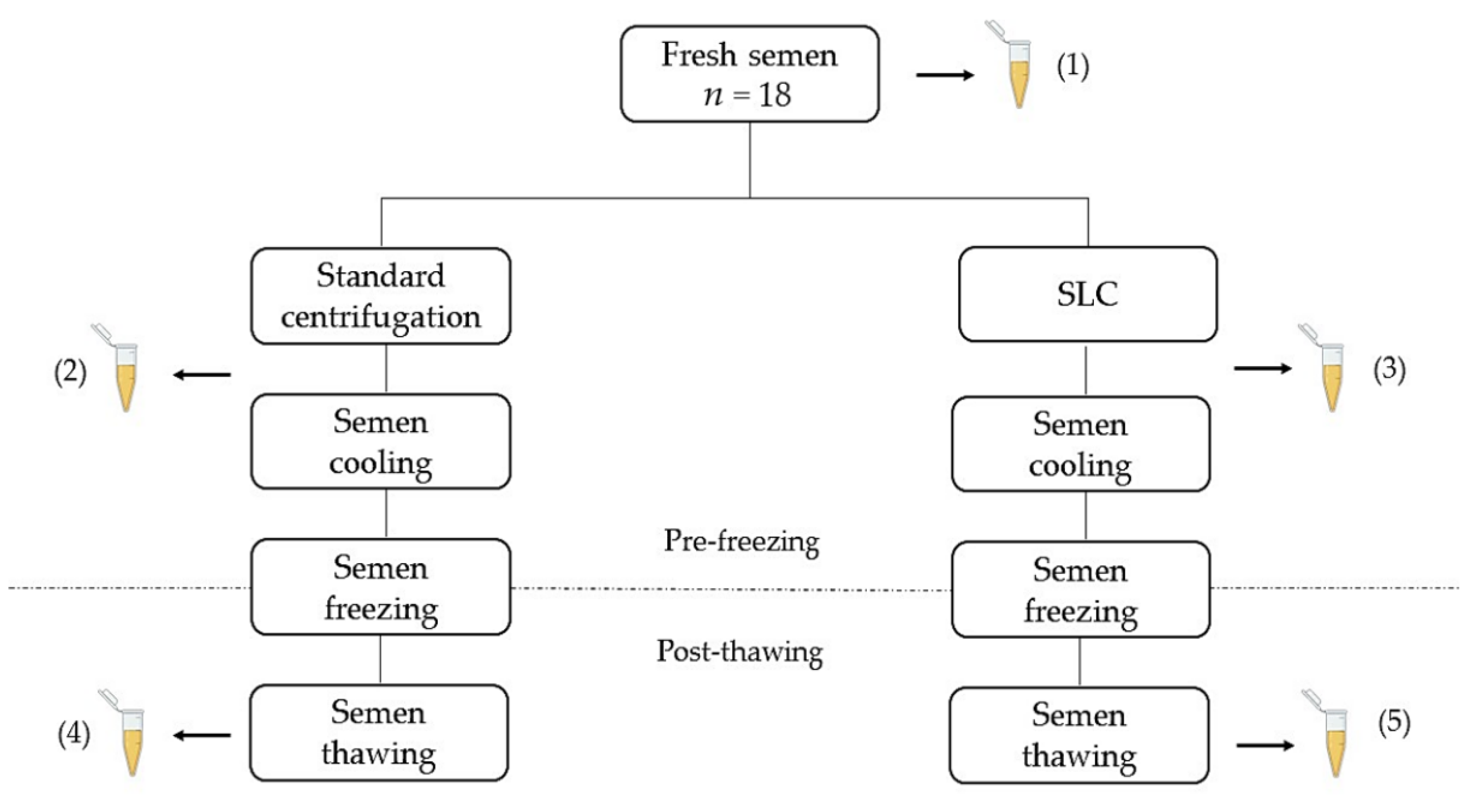
2.2. Semen Collection and Processing
2.3. Semen Quality Assessment
2.3.1. Concentration
2.3.2. Motility and Velocity
2.3.3. Morphology
2.3.4. Plasma Membrane Integrity, Acrosome Integrity and Mitochondrial Membrane Potential
2.3.5. Oxidative Stress
2.3.6. Sperm Chromatin Structure Assay (SCSA)
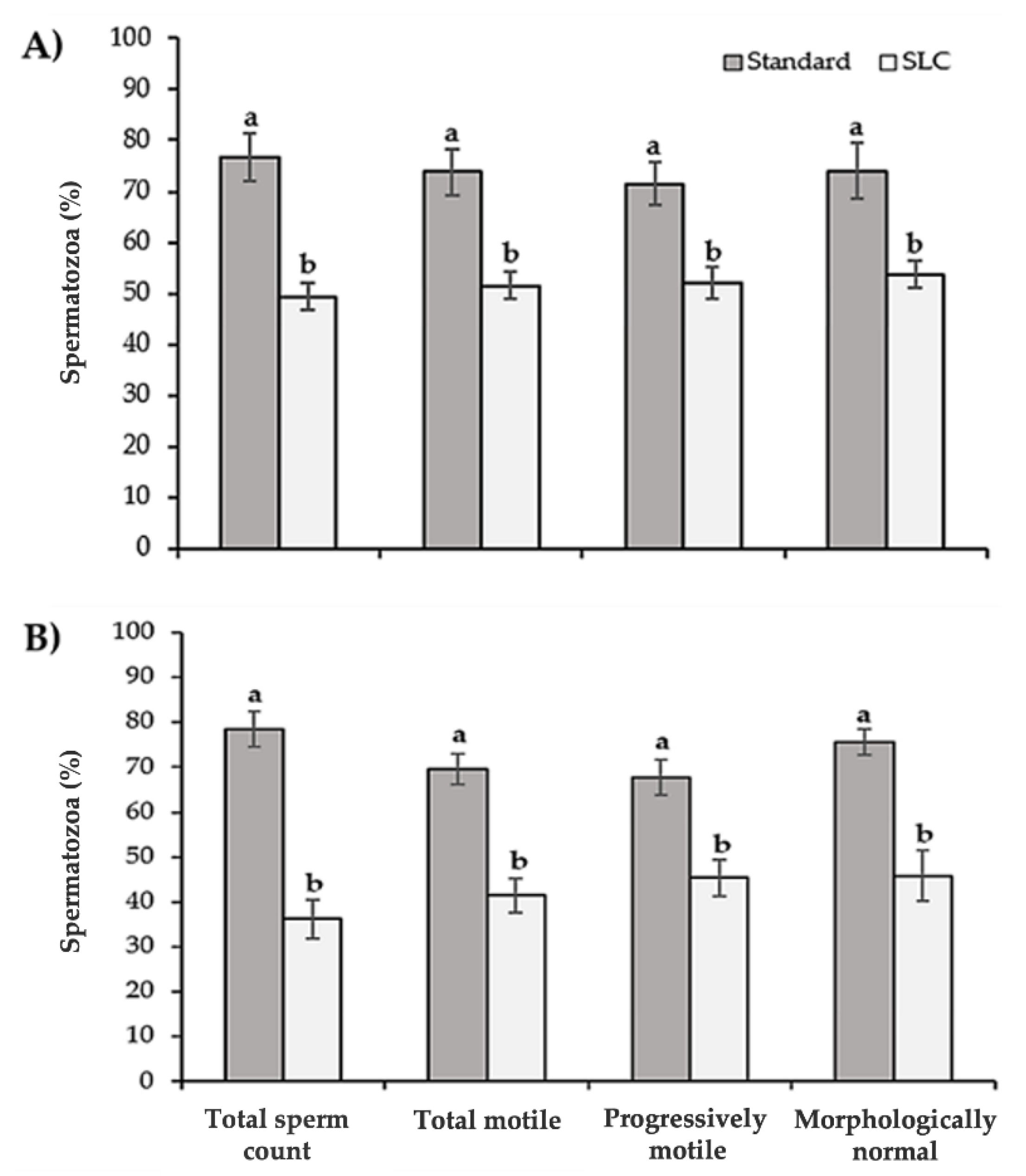
2.4. Recovery Rates
2.5. Statistical Analysis
3. Results
3.1. Pre-Freezing
Recovery Rates
3.2. Post-Thawing
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Linde-Forsberg, C. Achieving canine pregnancy by using frozen or chilled extended semen. Vet. Clin. N. Am. Small Anim. Pract. 1991, 21, 467–485. [Google Scholar] [CrossRef]
- Alamo, D.; Batista, M.; Gonzalez, F.; Rodriguez, N.; Cruz, G.; Cabrera, F.; Gracia, A. Cryopreservation of semen in the dog: Use of ultra-freezers of -152 degrees C as a viable alternative to liquid nitrogen. Theriogenology 2005, 63, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Domain, G.; Wydooghe, E.; Broeckx, B.J.G.; Hoogewijs, M.; Van Soom, A. Semen donation and establishment of an open canine semen bank: A novel tool to prevent inbreeding in pedigree dogs. Vlaams Diergen Tijds 2019, 88, 55–61. [Google Scholar] [CrossRef]
- Drobnis, E.Z.; Crowe, L.M.; Berger, T.; Anchordoguy, T.J.; Overstreet, J.W.; Crowe, J.H. Cold shock damage is due to lipid phase transitions in cell membranes: A demonstration using sperm as a model. J. Exp. Zool. 1993, 265, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Linde-Forsberg, C.; Strom Holst, B.; Govette, G. Comparison of fertility data from vaginal vs intrauterine insemination of frozen-thawed dog semen: A retrospective study. Theriogenology 1999, 52, 11–23. [Google Scholar] [CrossRef]
- Schafer-Somi, S.; Tichy, A. Canine post-thaw sperm quality can be predicted by using CASA, and classification and regression tree (CART)-analysis. Pol. J. Vet. Sci. 2019, 22, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Sieme, H.; Harrison, R.A.; Petrunkina, A.M. Cryobiological determinants of frozen semen quality, with special reference to stallion. Anim. Reprod. Sci. 2008, 107, 276–292. [Google Scholar] [CrossRef] [PubMed]
- Kuisma, P.; Andersson, M.; Koskinen, E.; Katila, T. Fertility of frozen-thawed stallion semen cannot be predicted by the currently used laboratory methods. Acta Vet. Scand. 2006, 48, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijnrocx, K.; Francois, L.; Stinckens, A.; Janssens, S.; Buys, N. Half of 23 Belgian dog breeds has a compromised genetic diversity, as revealed by genealogical and molecular data analysis. J. Anim. Breed. Genet. Z. Fur Tierz. Und Zucht. 2016, 133, 375–383. [Google Scholar] [CrossRef]
- Leroy, G. Genetic diversity, inbreeding and breeding practices in dogs: Results from pedigree analyses. Vet. J. 2011, 189, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Jansson, M.; Laikre, L. Pedigree data indicate rapid inbreeding and loss of genetic diversity within populations of native, traditional dog breeds of conservation concern. PLoS ONE 2018, 13, e0202849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bannasch, D.; Famula, T.; Donner, J.; Anderson, H.; Honkanen, L.; Batcher, K.; Safra, N.; Thomasy, S.; Rebhun, R. The effect of inbreeding, body size and morphology on health in dog breeds. Canine Med. Genet. 2021, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Morrell, J.M. Update on semen technologies for animal breeding. Reprod. Domest. Anim. Zuchthyg. 2006, 41, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Sikka, S.C. Relative impact of oxidative stress on male reproductive function. Curr. Med. Chem. 2001, 8, 851–862. [Google Scholar] [CrossRef] [PubMed]
- Kurkowska, W.; Bogacz, A.; Janiszewska, M.; Gabrys, E.; Tiszler, M.; Bellanti, F.; Kasperczyk, S.; Machon-Grecka, A.; Dobrakowski, M.; Kasperczyk, A. Oxidative Stress is Associated with Reduced Sperm Motility in Normal Semen. Am. J. Men's Health 2020, 14, 1557988320939731. [Google Scholar] [CrossRef] [PubMed]
- Alahmar, A.T. Role of Oxidative Stress in Male Infertility: An Updated Review. J. Hum. Reprod. Sci. 2019, 12, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Morrell, J.M.; Rodriguez-Martinez, H. Practical applications of sperm selection techniques as a tool for improving reproductive efficiency. Vet. Med. Int. 2010, 2011, 894767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrell, J.M.; Wallgren, M. Removal of bacteria from boar ejaculates by Single Layer Centrifugation can reduce the use of antibiotics in semen extenders. Anim. Reprod. Sci. 2011, 123, 64–69. [Google Scholar] [CrossRef]
- Morrell, J.M.; Klein, C.; Lundeheim, N.; Erol, E.; Troedsson, M.H. Removal of bacteria from stallion semen by colloid centrifugation. Anim. Reprod. Sci. 2014, 145, 47–53. [Google Scholar] [CrossRef]
- Guimaraes, T.; Lopes, G.; Pinto, M.; Silva, E.; Miranda, C.; Correia, M.J.; Damasio, L.; Thompson, G.; Rocha, A. Colloid centrifugation of fresh stallion semen before cryopreservation decreased microorganism load of frozen-thawed semen without affecting seminal kinetics. Theriogenology 2015, 83, 186–191. [Google Scholar] [CrossRef]
- Varela, E.; Rey, J.; Plaza, E.; Munoz de Propios, P.; Ortiz-Rodriguez, J.M.; Alvarez, M.; Anel-Lopez, L.; Anel, L.; De Paz, P.; Gil, M.C.; et al. How does the microbial load affect the quality of equine cool-stored semen? Theriogenology 2018, 114, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Blomqvist, G.; Persson, M.; Wallgren, M.; Wallgren, P.; Morrell, J.M. Removal of virus from boar semen spiked with porcine circovirus type 2. Anim. Reprod. Sci. 2011, 126, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Galvez, M.J.; Ortiz, I.; Hidalgo, M.; Morrell, J.M.; Dorado, J. Should single layer centrifugation of dog semen be done before or after the semen is cooled? Vet. Rec. 2015, 176, 359. [Google Scholar] [CrossRef] [PubMed]
- Dorado, J.; Galvez, M.J.; Morrell, J.M.; Alcaraz, L.; Hidalgo, M. Use of single-layer centrifugation with Androcoll-C to enhance sperm quality in frozen-thawed dog semen. Theriogenology 2013, 80, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Dorado, J.; Galvez, M.J.; Demyda-Peyras, S.; Ortiz, I.; Morrell, J.M.; Crespo, F.; Gosalvez, J.; Hidalgo, M. Differences in preservation of canine chilled semen using simple sperm washing, single-layer centrifugation and modified swim-up preparation techniques. Reprod. Fertil. Dev. 2015, 28, 1545–1552. [Google Scholar] [CrossRef] [PubMed]
- Urbano, M.; Dorado, J.; Ortiz, I.; Morrell, J.M.; Demyda-Peyras, S.; Galvez, M.J.; Alcaraz, L.; Ramirez, L.; Hidalgo, M. Effect of cryopreservation and single layer centrifugation on canine sperm DNA fragmentation assessed by the sperm chromatin dispersion test. Anim. Reprod. Sci. 2013, 143, 118–125. [Google Scholar] [CrossRef]
- Hoogewijs, M.; Morrell, J.; Van Soom, A.; Govaere, J.; Johannisson, A.; Piepers, S.; De Schauwer, C.; De Kruif, A.; De Vliegher, S. Sperm selection using single layer centrifugation prior to cryopreservation can increase thawed sperm quality in stallions. Equine Vet. J. Suppl. 2011, 43, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Alborcia, M.J.; Morrell, J.M.; Parrilla, I.; Barranco, I.; Vazquez, J.M.; Martinez, E.A.; Roca, J. Improvement of boar sperm cryosurvival by using single-layer colloid centrifugation prior freezing. Theriogenology 2012, 78, 1117–1125. [Google Scholar] [CrossRef]
- Martinez-Alborcia, M.J.; Morrell, J.M.; Gil, M.A.; Barranco, I.; Maside, C.; Alkmin, D.V.; Parrilla, I.; Martinez, E.A.; Roca, J. Suitability and effectiveness of single layer centrifugation using Androcoll-P in the cryopreservation protocol for boar spermatozoa. Anim. Reprod. Sci. 2013, 140, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Nongbua, T.; Johannisson, A.; Edman, A.; Morrell, J.M. Effects of single layer centrifugation (SLC) on bull spermatozoa prior to freezing on post-thaw semen characteristics. Reprod. Domest. Anim. Zuchthyg. 2017, 52, 596–602. [Google Scholar] [CrossRef]
- Farstad, W.K. Artificial insemination in dogs. In BSAVA Manual of Canine and Feline Reproduction and Neonatology, 2nd ed.; England, G., von Heimendahl, A., Eds.; British Small Animal Veterinary Association: Gloucester, UK, 2010; pp. 80–88. [Google Scholar]
- Johnston, S.D.; Root Kustritz, M.V.R.; Olson, P.N. Semen collection, evaluation and preservation. In Canine and Feline Theriogenology, 1st ed.; Kersey, R., Ed.; Saunders: Philadelphia, PA, USA, 2001; pp. 287–306. [Google Scholar]
- Oettle, E.E. Sperm Morphology and Fertility in the Dog. J. Reprod. Fertil. 1993, 47, 257–260. [Google Scholar]
- Rijsselaere, T.; Van Soom, A.; Maes, D.; de Kruif, A. Effect of centrifugation on in vitro survival of fresh diluted canine spermatozoa. Theriogenology 2002, 57, 1669–1681. [Google Scholar] [CrossRef]
- Morrell, J.M.; Nunes, M.M. Practical guide to single layer centrifugation of stallion semen. Equine Vet. Educ. 2018, 30, 392–398. [Google Scholar] [CrossRef]
- Pena, A.; Linde-Forsberg, C. Effects of Equex, one- or two-step dilution, and two freezing and thawing rates on post-thaw survival of dog spermatozoa. Theriogenology 2000, 54, 859–875. [Google Scholar] [CrossRef]
- Schafer-Somi, S.; Kluger, S.; Knapp, E.; Klein, D.; Aurich, C. Effects of semen extender and semen processing on motility and viability of frozen-thawed dog spermatozoa. Theriogenology 2006, 66, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Morrell, J.M.; Johannisson, A.; Juntilla, L.; Rytty, K.; Backgren, L.; Dalin, A.M.; Rodriguez-Martinez, H. Stallion Sperm Viability, as Measured by the Nucleocounter SP-100, Is Affected by Extender and Enhanced by Single Layer Centrifugation. Vet. Med. Int. 2010, 2010, 659862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rijsselaere, T.; Van Soom, A.; Maes, D.; de Kruif, A. Effect of technical settings on canine semen motility parameters measured by the Hamilton-Thorne analyzer. Theriogenology 2003, 60, 1553–1568. [Google Scholar] [CrossRef]
- Hoogewijs, M.K.; de Vliegher, S.P.; Govaere, J.L.; de Schauwer, C.; de Kruif, A.; van Soom, A. Influence of counting chamber type on CASA outcomes of equine semen analysis. Equine Vet. J. 2012, 44, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Angrimani, D.S.R.; Bicudo, L.C.; Luceno, N.L.; Leemans, B.; Nichi, M.; Vannucchi, C.I.; Van Soom, A. A triple stain method in conjunction with an in-depth screening of cryopreservation effects on post-thaw sperm in dogs. Cryobiology 2021, 105, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Nichi, M.; Goovaerts, I.G.; Cortada, C.N.; Barnabe, V.H.; De Clercq, J.B.; Bols, P.E. Roles of lipid peroxidation and cytoplasmic droplets on in vitro fertilization capacity of sperm collected from bovine epididymides stored at 4 and 34 degrees C. Theriogenology 2007, 67, 334–340. [Google Scholar] [CrossRef]
- Intasqui, P.; Antoniassi, M.P.; Camargo, M.; Nichi, M.; Carvalho, V.M.; Cardozo, K.H.; Zylbersztejn, D.S.; Bertolla, R.P. Differences in the seminal plasma proteome are associated with oxidative stress levels in men with normal semen parameters. Fertil. Steril. 2015, 104, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Evenson, D.P.; Darzynkiewicz, Z.; Melamed, M.R. Relation of mammalian sperm chromatin heterogeneity to fertility. Science 1980, 210, 1131–1133. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Machler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Miller, J.J. Inverse of Freeman-Tukey Double Arcsine Transformation. Am. Stat. 1978, 32, 138. [Google Scholar] [CrossRef]
- Huang, Y.L.; Tseng, W.C.; Cheng, S.Y.; Lin, T.H. Trace elements and lipid peroxidation in human seminal plasma. Biol. Trace Elem. Res. 2000, 76, 207–215. [Google Scholar] [CrossRef]
- Karoui, S.; Diaz, C.; Gonzalez-Marin, C.; Amenabar, M.E.; Serrano, M.; Ugarte, E.; Gosalvez, J.; Roy, R.; Lopez-Fernandez, C.; Carabano, M.J. Is sperm DNA fragmentation a good marker for field AI bull fertility? J. Anim. Sci. 2012, 90, 2437–2449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narud, B.; Klinkenberg, G.; Khezri, A.; Zeremichael, T.T.; Stenseth, E.B.; Nordborg, A.; Haukaas, T.H.; Morrell, J.M.; Heringstad, B.; Myromslien, F.D.; et al. Differences in sperm functionality and intracellular metabolites in Norwegian Red bulls of contrasting fertility. Theriogenology 2020, 157, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Fuente-Lara, A.; Hesser, A.; Christensen, B.; Gonzales, K.; Meyers, S. Effects from aging on semen quality of fresh and cryopreserved semen in Labrador Retrievers. Theriogenology 2019, 132, 164–171. [Google Scholar] [CrossRef]
- Lechner, D.; Aurich, J.; Schafer-Somi, S.; Aurich, C. Effects of age, size and season on cryotolerance of dog semen—A retrospective analysis. Anim. Reprod. Sci. 2022, 236, 106912. [Google Scholar] [CrossRef] [PubMed]
- Tesi, M.; Sabatini, C.; Vannozzi, I.; Di Petta, G.; Panzani, D.; Camillo, F.; Rota, A. Variables affecting semen quality and its relation to fertility in the dog: A retrospective study. Theriogenology 2018, 118, 34–39. [Google Scholar] [CrossRef]
- Mason, S.J. Current Review of Artificial Insemination in Dogs. Vet. Clin. N. Am. Small Anim. Pract. 2018, 48, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Tsutsui, T.; Hori, T.; Yamada, A.; Kirihara, N.; Kawakami, E. Intratubal insemination with fresh semen in dogs. J. Vet. Med. Sci. 2003, 65, 659–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagashima, J.B.; Sylvester, S.R.; Nelson, J.L.; Cheong, S.H.; Mukai, C.; Lambo, C.; Flanders, J.A.; Meyers-Wallen, V.N.; Songsasen, N.; Travis, A.J. Live Births from Domestic Dog (Canis familiaris) Embryos Produced by In Vitro Fertilization. PLoS ONE 2015, 10, e0143930. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Sperm Cell Pattern | PI | FITC-PNA | JC-1 |
|---|---|---|---|
| Intact plasma membrane, intact acrosome, high MMP (I) | − | − | Orange |
| Intact plasma membrane, intact acrosome, low MMP (II) | − | − | Green |
| Intact plasma membrane, damaged acrosome, high MMP (III) | − | + | Orange |
| Intact plasma membrane, damaged acrosome, low MMP (IV) | − | + | Green |
| Damaged plasma membrane, intact acrosome, high MMP (V) | + | − | Orange |
| Damaged plasma membrane, intact acrosome, low MMP (VI) | + | − | Green |
| Damaged plasma membrane, damaged acrosome, high MMP (VII) | + | + | Orange |
| Damaged plasma membrane, damaged acrosome, low MMP (VIII) | + | + | Green |
| Optimal Quality | Suboptimal Quality | |||||
|---|---|---|---|---|---|---|
| Parameter | Fresh | Standard | SLC | Fresh | Standard | SLC |
| Total motility (%) | 89.0 ± 1.1 b | 85.6 ± 1.3 b | 92.9 ± 0.7 a | 75.7 ± 3.8 y | 66.7 ± 4.0 z | 88.4 ± 1.9 x |
| Progressive motility (%) | 81.0 ± 1.4 a | 75.7 ± 1.6 b | 85.2 ± 0.6 a | 64.4 ± 3.9 y | 55.3 ± 4.1 z | 82.2 ± 2.0 x |
| Curvilinear velocity (µm/s) | 175.3 ± 7.1 a | 167.0 ± 3.9 b | 174.1 ± 3.9 a | 164.8 ± 7.8 x | 150.9 ± 4.9 y | 160.8 ± 5.7 x |
| Straight line velocity (µm/s) | 108.9 ± 1.2 b | 111.1 ± 3.8 b | 124.4 ± 2.9 a | 96.1 ± 6.0 y | 93.9 ± 4.5 y | 117.4 ± 3.4 x |
| Average path velocity (µm/s) | 131.7 ± 1.7 b | 133.7 ± 4.1 b | 147.8 ± 2.5 a | 118.3 ± 5.9 y | 114.3 ± 4.6 y | 135.9 ± 4.3 x |
| Linearity (%) | 62.4 ± 2.2 b | 66.2 ± 1.9 b | 71.1 ± 2.0 a | 57.8 ± 1.6 y | 61.8 ± 1.6 y | 72.7 ± 0.7 x |
| Straightness (%) | 82.4 ± 0.8 | 82.6 ± 0.7 | 83.6 ± 1.1 | 80.4 ± 1.1 y | 81.3 ± 0.7 y | 86.1 ± 0.3 x |
| Wobble (%) | 75.4 ± 2.1 c | 79.8 ± 1.8 b | 84.8 ± 1.4 a | 70.0 ± 1.9 z | 75.2 ± 1.5 y | 84.2 ± 0.5 x |
| Amplitude of lateral head displacement (µm) | 2.2 ± 0.3 | 2.1 ± 0.1 | 1.9 ± 0.1 | 2.3 ± 0.2 | 2.0 ± 0.0 | 1.8 ± 0.1 |
| Beat cross frequency (Hz) | 23.9 ± 0.4 a | 21.2 ± 0.7 b | 21.0 ± 0.6 b | 22.6 ± 0.8 x | 20.8 ± 0.4 y | 21.2 ± 0.4 y |
| Normal morphology (%) | 86.7 ± 1.8 b | 83.1 ± 2.6 b | 94.4 ± 0.9 a | 67.8 ± 2.0 y | 65.1 ± 2.8 y | 86.1 ± 2.9 x |
| Head defects (%) | 4.3 ± 0.9 a | 3.1 ± 0.7 a | 2.0 ± 0.2 b | 10.2 ± 2.4 x | 9.4 ± 2.6 x | 5.6 ± 1.6 y |
| Midpiece/tail defects (%) | 3.2 ± 1.0 b | 10.0 ± 3.0 a | 1.2 ± 0.3 c | 11.0 ± 1.9 y | 17.0 ± 2.4 x | 4.6 ± 2.1 z |
| Proximal protoplasmic droplet (%) | 4.0 ± 1.2 a | 2.5 ± 1.0 a | 2.3 ± 0.7 b | 6.2 ± 1.6 x | 6.2 ± 1.2 x | 2.7 ± 0.8 y |
| Distal protoplasmic droplet (%) | 1.7 ± 0.9 a | 1.3 ± 0.4 a | 0.1 ± 0.1 b | 4.7 ± 1.4 x | 2.2 ± 1.0 x | 0.6 ± 0.2 y |
| Intact plasma membrane (%) | 89.3 ± 1.3 a,b | 86.3 ± 1.1 b | 89.5 ± 1.2 a | 79.4 ± 3.7 x,y | 78.7 ± 4.1 y | 82.8 ± 3.2 x |
| Intact acrosome (%) | 92.6 ± 1.3 | 91.2 ± 1.0 | 89.7 ± 2.1 | 85.4 ± 3.1 | 87.2 ± 2.7 | 85.4 ± 4.0 |
| High mitochondrial membrane potential (%) | 81.6 ± 3.2 b | 81.6 ± 2.0 b | 90.4 ± 1.0 a | 74.6 ± 5.0 y | 69.0 ± 6.4 y | 80.0 ± 4.6 x |
| DNA fragmentation index (%) | 10.5 ± 3.1 a | 8.2 ± 0.8 a | 5.6 ± 0.8 b | 20.0 ± 4.4 x | 14.5 ± 2.9 x | 8.7 ± 1.2 y |
| TBA-RS (ng/106 sperm) | 8.3 ± 2.5 a | 3.2 ± 0.3 b | 3.2 ± 0.5 b | 8.9 ± 2.0 x | 4.2 ± 0.9 y | 4.2 ± 0.6 y |
| Optimal Quality | Suboptimal Quality | |||
|---|---|---|---|---|
| Parameter | Standard | SLC | Standard | SLC |
| Total motility (%) | 59.2 ± 3.8 | 53.4 ± 5.2 | 36.2 ± 5.1 | 38.6 ± 4.2 |
| Progressive motility (%) | 47.1 ± 3.7 | 43.4 ± 4.7 | 26.1 ± 4.7 | 28.8 ± 3.9 |
| Curvilinear velocity (µm/s) | 146.2 ± 6.9 a | 139.4 ± 6.4 b | 132.1 ± 6.9 x | 121.9 ± 6.3 y |
| Straight line velocity (µm/s) | 73.8 ± 4.0 | 74.6 ± 3.3 | 65.2 ± 3.4 | 64.2 ± 4.0 |
| Average path velocity (µm/s) | 97.5 ± 4.1 | 96.1 ± 3.8 | 85.3 ± 3.7 | 82.6 ± 3.8 |
| Linearity (%) | 50.3 ± 2.1 b | 53.5 ± 2.3 a | 49.3 ± 1.3 y | 52.2 ± 1.4 x |
| Straightness (%) | 75.2 ± 2.1 | 77.3 ± 2.2 | 76.1 ± 0.8 | 76.9 ± 1.4 |
| Wobble (%) | 66.6 ± 1.3 b | 68.9 ± 1.3 a | 64.4 ± 1.2 y | 67.7 ± 0.9 x |
| Amplitude of lateral head displacement (µm) | 2.2 ± 0.1 | 2.0 ± 0.0 | 2.1 ± 0.1 | 2.0 ± 0.0 |
| Beat cross frequency (Hz) | 21.9 ± 0.8 | 21.9 ± 0.9 | 21.3 ± 0.3 | 21.9 ± 0.6 |
| Normal morphology (%) | 68.9 ± 2.7 b | 80.2 ± 2.4 a | 59.2 ± 3.4 y | 76.1 ± 5.0 x |
| Head defects (%) | 12.0 ± 2.7 | 13.9 ± 2.8 | 13.2 ± 2.6 | 14.9 ± 3.8 |
| Midpiece/tail defects (%) | 12.8 ± 3.0 a | 3.9 ± 0.8 b | 20.0 ± 4.2 x | 4.8 ± 2.1 y |
| Proximal protoplasmic droplet (%) | 4.0 ± 1.4 a | 1.8 ± 1.0 b | 5.2 ± 1.6 x | 1.9 ± 0.4 y |
| Distal protoplasmic droplet (%) | 2.2 ± 1.0 a | 0.3 ± 0.2 b | 1.9 ± 0.5 x | 0.8 ± 0.5 y |
| Intact plasma membrane (%) | 59.6 ± 2.9 b | 62.3 ± 3.6 a | 42.3 ± 4.8 y | 48.6 ± 3.4 x |
| Intact acrosome (%) | 68.3 ± 3.2 | 67.7 ± 3.8 | 55.0 ± 5.0 | 59.1 ± 2.8 |
| High mitochondrial membrane potential (%) | 27.8 ± 4.2 b | 37.3 ± 4.5 a | 25.9 ± 4.0 | 24.9 ± 4.1 |
| DNA fragmentation index (%) | 6.8 ± 0.8 | 6.6 ± 0.9 | 13.6 ± 3.1 | 7.8 ± 0.8 |
| TBA-RS (ng/106 sperm) | 2.6 ± 0.4 | 2.8 ± 0.4 | 4.3 ± 0.8 | 4.2 ± 0.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domain, G.; Ali Hassan, H.; Wydooghe, E.; Bogado Pascottini, O.; Johannisson, A.; Morrell, J.M.; Niżański, W.; Van Soom, A. Influence of Single Layer Centrifugation with Canicoll on Semen Freezability in Dogs. Animals 2022, 12, 714. https://doi.org/10.3390/ani12060714
Domain G, Ali Hassan H, Wydooghe E, Bogado Pascottini O, Johannisson A, Morrell JM, Niżański W, Van Soom A. Influence of Single Layer Centrifugation with Canicoll on Semen Freezability in Dogs. Animals. 2022; 12(6):714. https://doi.org/10.3390/ani12060714
Chicago/Turabian StyleDomain, Guillaume, Hiba Ali Hassan, Eline Wydooghe, Osvaldo Bogado Pascottini, Anders Johannisson, Jane M. Morrell, Wojciech Niżański, and Ann Van Soom. 2022. "Influence of Single Layer Centrifugation with Canicoll on Semen Freezability in Dogs" Animals 12, no. 6: 714. https://doi.org/10.3390/ani12060714