
Growth and Fatty Acid Composition of Black Soldier Fly Hermetia illucens (Diptera: Stratiomyidae) Larvae Are Influenced by Dietary Fat Sources and Levels


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Experimental Diets
2.2. Rearing Trial of Black Soldier Fly Larvae
2.3. Sampling
2.4. Proximate Nutrients Analysis
2.5. Fatty Acid Composition Analysis
2.6. Statistical Analysis
3. Results
3.1. Growth
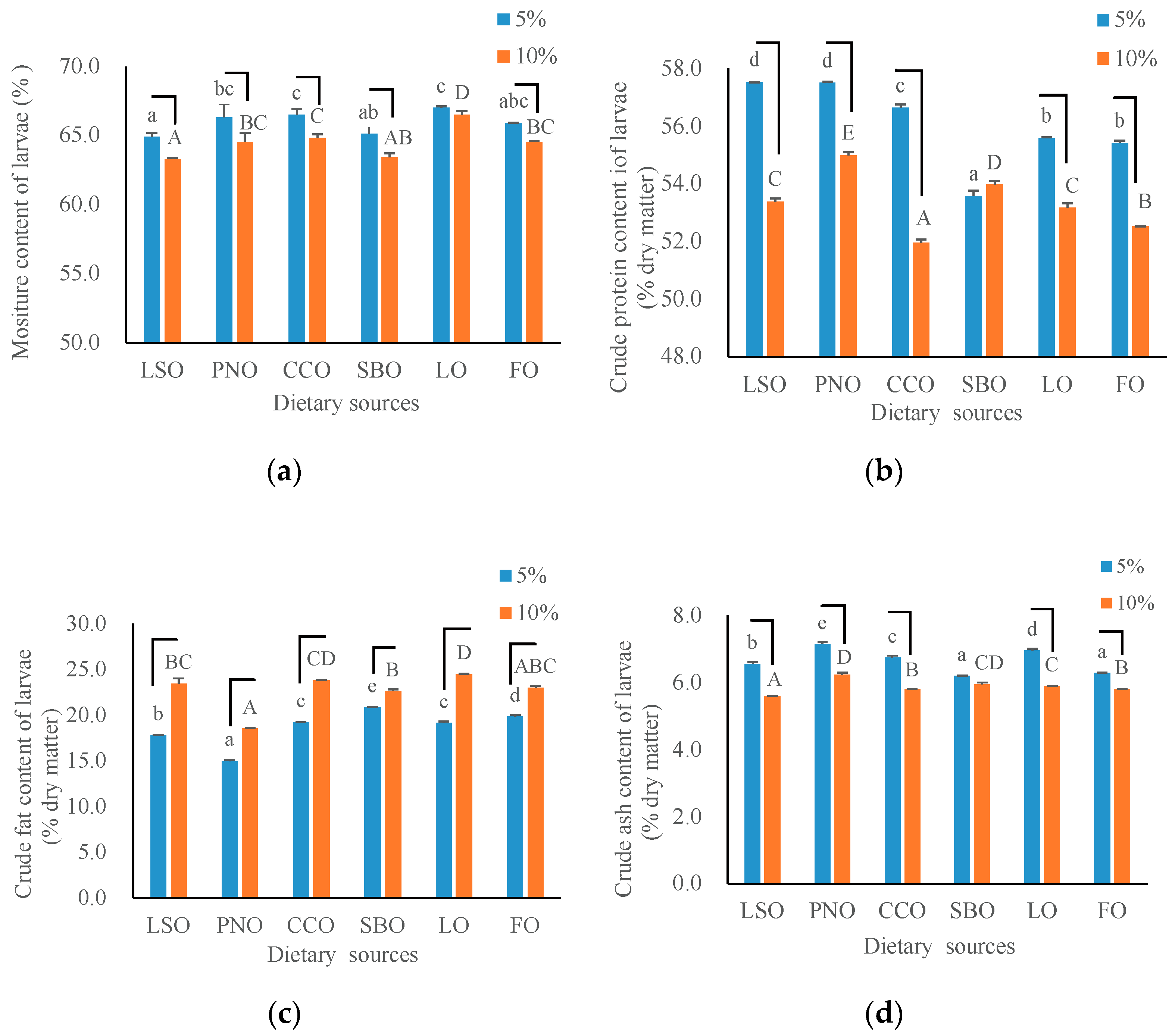
3.2. The Nutrient Composition of Larvae
3.3. Fatty Acid Composition
4. Discussion
4.1. Growth of Larvae
4.2. The Nutrient Composition of Larvae
4.3. Fatty Acid Composition
- (1)
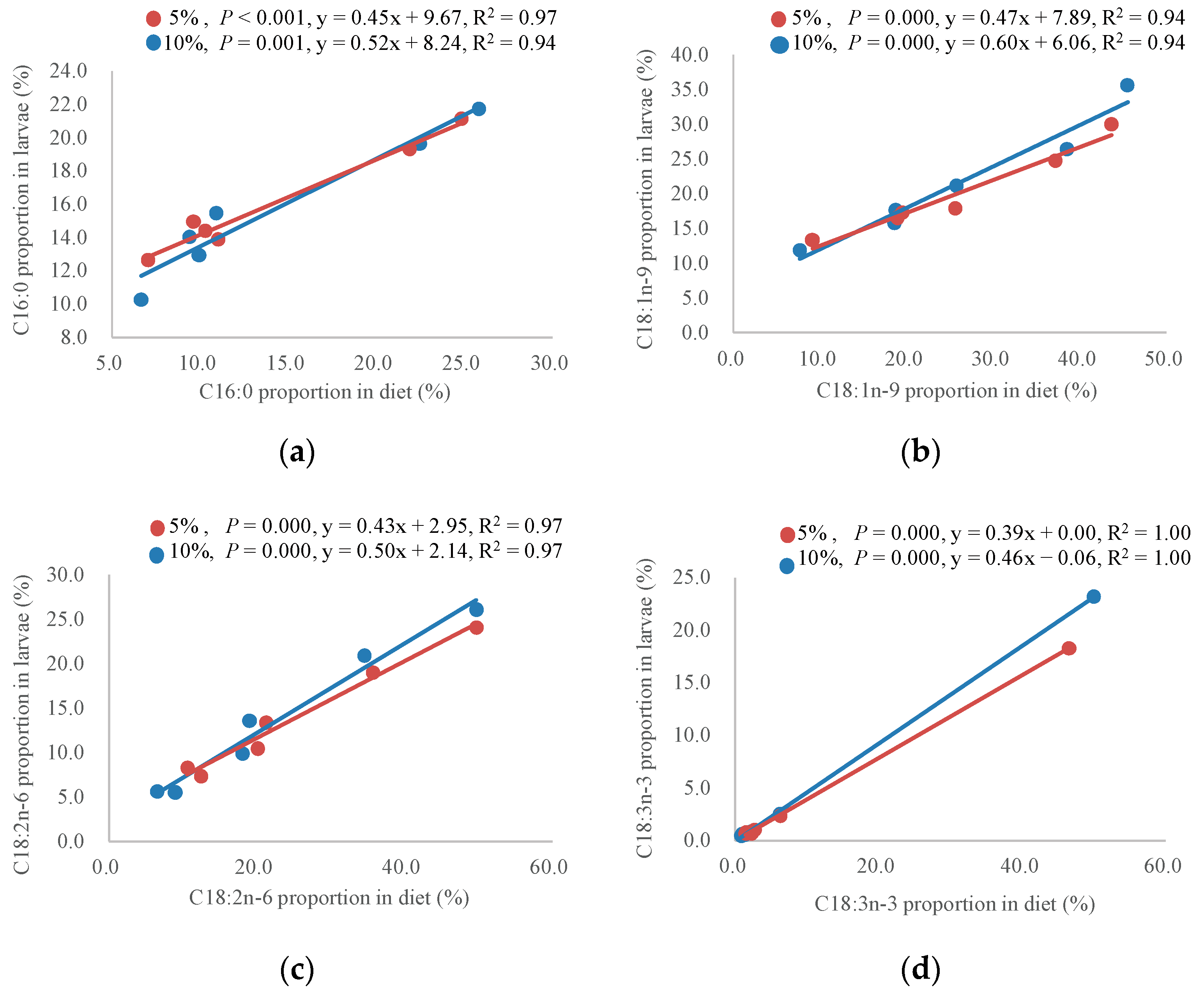
- C12:0, C14:0 and C16:1n-9. Their proportions in larvae were much higher in larvae than those in diets. For example, C12:0 was the most abundant (17.4–44.9%) fatty acid in larvae in all groups, but it can hardly be found in most of the oil sources except that of coconut oil. Similarly, a larger proportion of C14:0 (3.9–15.2%) and C16:1n-9 (1.3–3.7%) in larvae over most diets was determined, compared with that of them in diets. According to previous studies, the high proportion of C12:0 in larvae can distinguish the black soldier fly from other insects [17,53,54]. Several saturated fatty acids including C10:0, C12:0, and C14:0 were biosynthesized through the elongation process with carbohydrate as an essential source of acetyl-CoA. In addition, C16:1n-9 was produced through a desaturation process based on 16:0 [36]. Our data seemed to support this suggestion. However, C10:0 was not determined in larvae in this study. Moreover, it was apparent that dietary fatty acids and carbohydrates contributed to the bioaccumulation of C12:0 and C14:0. They were properly bioaccumulated by larvae up to almost 44.9% and 15.2% in the coconut group, respectively, indicating that the larvae may incorporate C12:0 and C14:0 when they occurr in diets. Interestingly, higher dietary fat content could enhance the bioaccumulation of C12:0 and C14:0. Hence, the enrichment of the nutritive value of C12:0 in black soldier fly larvae by manipulating fat content and fatty acid profiles may be technically feasible.
- (2)
- C16:0, C18:1n-9, C18:2n-6 and C18:3n-3. Their proportions in larvae were linearly related to those of them in diets. Compared with previous studies, the present study presented a quantitative assessment of the relationship of these fatty acids between larvae and diets, so that the proportions of the four fatty acids in larvae could be estimated from their proportions in diets. According to Hoc et al. (2020) [36], 16:0 and 18:1n-9 thought to be merely bioaccumulated from diets were also produced by the larvae itself, 18:2n-6 and 18:3n-3 could be partly retained in larval body, and the rest of them could be generated into acetyl-CoA through fatty acid β-oxidation. Our results were partly consistent with these suggestions. Specifically, C18:2n-6 and C18:3n-3 proportions in larvae (5.5–26.1% and 0.5–23.2%) were significantly reduced compared to those in diets (6.5–49.8% and 0.9–50.0%), reflecting that it was less economic for larvae to acquire the two fatty acids from diets. However, the larger proportions of C16:0 in larvae (10.3–15.5%) over diets (6.7–11.0%) were observed in linseed oil, peanut oil, coconut oil and soybean oil groups, while its proportions in larvae were around 20% in the lard oil and fish oil groups, despite the two diets containing high proportions of 16:0 (21.9–25.9%). Similar results were revealed for 18:1n-9. Therefore, we expected that the bioaccumulation of the two fatty acids in the body was limited, and that the extra part of them may be used for supplying oxidation energy, or biosynthesis as other fatty acids. Therefore, despite the high levels of C18:2n-6 and C18:3n-3 in plant oil, and EPA and DHA in fish oil, possibilities to tailor them into black soldier fly larvae through diets maybe limited and not cost-effective. Nevertheless, as a promising source of SFA, especially C12:0, black soldier fly larvae could work well in combination with oil rich in MUFA and PUFA to satisfy the fatty acid requirement of aquatic animals.
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Boland, M.J.; Rae, A.N.; Vereijken, J.M.; Meuwissen, M.P.M.; Fischer, A.R.H.; van Boekel, M.A.J.S.; Rutherfurd, S.M.; Gruppen, H.; Moughan, P.J.; Hendriks, W.H. The future supply of animal-derived protein for human consumption. Trends Food Sci. Technol. 2013, 29, 62–73. [Google Scholar] [CrossRef]
- Govorushko, S. Global status of insects as food and feed source: A review. Trends Food Sci. Technol. 2019, 91, 436–445. [Google Scholar] [CrossRef]
- Bessa, L.W.; Pieterse, E.; Marais, J.; Hoffman, L.C. Why for feed and not for human consumption? The black soldier fly larvae. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2747–2763. [Google Scholar] [CrossRef] [PubMed]
- Biasato, I.; Renna, M.; Gai, F.; Dabbou, S.; Meneguz, M.; Perona, G.; Martinez, S.; Lajusticia, A.C.B.; Bergagna, S.; Sardi, L.; et al. Partially defatted black soldier fly larva meal inclusion in piglet diets: Effects on the growth performance, nutrient digestibility, blood profile, gut morphology and histological features. J. Anim. Sci. Biotechnol. 2019, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Schiavone, A.; Cullere, M.; De Marco, M.; Meneguz, M.; Biasato, I.; Bergagna, S.; Dezzutto, D.; Gai, F.; Dabbou, S.; Gasco, L.; et al. Partial or total replacement of soybean oil by black soldier fly larvae (Hermetia illucens L.) fat in broiler diets: Effect on growth performances, feed-choice, blood traits, carcass characteristics and meat quality. Ital. J. Anim. Sci. 2017, 16, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.B.; Kim, D.-H.; Jeong, S.-B.; Lee, J.-W.; Kim, T.-H.; Lee, H.-G.; Lee, K.-W. Black soldier fly larvae oil as an alternative fat source in broiler nutrition. Poult. Sci. 2020, 99, 3133–3143. [Google Scholar] [CrossRef]
- Heuel, M.; Sandrock, C.; Leiber, F.; Mathys, A.; Gold, M.; Zurbrügg, C.; Gangnat, I.; Kreuzer, M.; Terranova, M. Black soldier fly larvae meal and fat can completely replace soybean cake and oil in diets for laying hens. Poult. Sci. 2021, 100, 101034. [Google Scholar] [CrossRef]
- Xu, X.; Ji, H.; Belghit, I.; Liland, N.S.; Wu, W.; Li, X. Effects of black soldier fly oil rich in n-3 HUFA on growth performance, metabolism and health response of juvenile mirror carp (Cyprinus carpio var. specularis). Aquaculture 2021, 533, 736144. [Google Scholar] [CrossRef]
- Kumar, V.; Fawole, F.J.; Romano, N.; Hossain, S.; Labh, S.N.; Overturf, K.; Small, B.C. Insect (black soldier fly, Hermetia illucens) meal supplementation prevents the soybean meal-induced intestinal enteritis in rainbow trout and health benefits of using insect oil. Fish Shellfish Immunol. 2020, 109, 116–124. [Google Scholar] [CrossRef]
- Rawski, M.; Mazurkiewicz, J.; Kierończyk, B.; Józefiak, D. Black Soldier Fly Full-Fat Larvae Meal as an Alternative to Fish Meal and Fish Oil in Siberian Sturgeon Nutrition: The Effects on Physical Properties of the Feed, Animal Growth Performance, and Feed Acceptance and Utilization. Animals 2020, 10, 2119. [Google Scholar] [CrossRef]
- Tomberlin, J.K.; Cammack, J.A. Black Soldier Fly: Biology and Mas Production, 1st ed.; Tomberlin, J.K., Ed.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2017. [Google Scholar]
- Henry, M.; Gasco, L.; Piccolo, G.; Fountoulaki, E. Review on the use of insects in the diet of farmed fish: Past and future. Anim. Feed Sci. Technol. 2015, 203, 1–22. [Google Scholar] [CrossRef]
- Müller, A.; Wolf, D.; Gutzeit, H.O. The black soldier fly, Hermetia illucens—A promising source for sustainable production of proteins, lipids and bioactive substances. Z. Naturforschung C 2017, 72, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Caligiani, A.; Marseglia, A.; Sorci, A.; Bonzanini, F.; Lolli, V.; Maistrello, L.; Sforza, S. Influence of the killing method of the black soldier fly on its lipid composition. Food Res. Int. 2019, 116, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Rabani, V.; Cheatsazan, H.; Davani, S. Proteomics and Lipidomics of Black Soldier Fly (Diptera: Stratiomyidae) and Blow Fly (Diptera: Calliphoridae) Larvae. J. Insect Sci. 2019, 19, 29. [Google Scholar] [CrossRef] [PubMed]
- Barroso, F.G.; Sánchez-Muros, M.-J.; Segura, M.; Morote, E.; Torres, A.; Ramos, R.; Guil, J.-L. Insects as food: Enrichment of larvae of Hermetia illucens with omega 3 fatty acids by means of dietary modifications. J. Food Compos. Anal. 2017, 62, 8–13. [Google Scholar] [CrossRef]
- Ewald, N.; Vidakovic, A.; Langeland, M.; Kiessling, A.; Sampels, S.; Lalander, C. Fatty acid composition of black soldier fly larvae (Hermetia illucens)—Possibilities and limitations for modification through diet. Waste Manag. 2020, 102, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.-W.; Mohd-Noor, S.-N.; Wong, C.-Y.; Lam, M.K.; Goh, P.-S.; Beniers, J.; Oh, W.-D.; Jumbri, K.; Ghani, N.A. Palatability of black soldier fly larvae in valorizing mixed waste coconut endosperm and soybean curd residue into larval lipid and protein sources. J. Environ. Manag. 2019, 231, 129–136. [Google Scholar] [CrossRef]
- Cullere, M.; Woods, M.J.; Van Emmenes, L.; Pieterse, E.; Hoffman, L.C.; Dalle Zotte, A. Hermetia illucens Larvae Reared on Different Substrates in Broiler Quail Diets: Effect on Physicochemical and Sensory Quality of the Quail Meat. Animals 2019, 9, 525. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Awasthi, M.K.; Chen, H.; Duan, Y.; Awasthi, S.K.; Zhang, Z. Performance of black soldier fly larvae (Diptera: Stratiomyidae) for manure composting and production of cleaner compost. J. Environ. Manag. 2019, 251, 109593. [Google Scholar] [CrossRef] [PubMed]
- Liland, N.S.; Biancarosa, I.; Araujo, P.; Biemans, D.; Bruckner, C.G.; Waagbø, R.; Torstensen, B.E.; Lock, E.-J. Modulation of nutrient composition of black soldier fly (Hermetia illucens) larvae by feeding seaweed-enriched media. PLoS ONE 2017, 12, e0183188. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; Van Broekhoven, S.; Van Huis, A.; Van Loon, J.J.A. Feed Conversion, Survival and Development, and Composition of Four Insect Species on Diets Composed of Food By-Products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef] [Green Version]
- Oonincx, D.G.A.B.; Laurent, S.; Veenenbos, M.E.; Van Loon, J.J. Dietary enrichment of edible insects with omega 3 fatty acids. Insect Sci. 2020, 27, 500–509. [Google Scholar] [CrossRef] [Green Version]
- Erbland, P.; Alyokhin, A.; Perkins, L.B.; Peterson, M. Dose-Dependent Retention of Omega-3 Fatty Acids by Black Soldier Fly Larvae (Diptera: Stratiomyidae). J. Econ. Èntomol. 2020, 113, 1221–1226. [Google Scholar] [CrossRef]
- Truzzi, C.; Giorgini, E.; Annibaldi, A.; Antonucci, M.; Illuminati, S.; Scarponi, G.; Riolo, P.; Isidoro, N.; Conti, C.; Zarantoniello, M.; et al. Fatty acids profile of black soldier fly (Hermetia illucens): Influence of feeding substrate based on coffee-waste silverskin enriched with microalgae. Anim. Feed Sci. Technol. 2020, 259, 114309. [Google Scholar] [CrossRef]
- Ramos-Bueno, R.P.; González-Fernández, M.J.; Sánchez-Muros-Lozano, M.J.; García-Barroso, F.; Guil-Guerrero, J.L. Fatty acid profiles and cholesterol content of seven insect species assessed by several extraction systems. Eur. Food Res. Technol. 2016, 242, 1471–1477. [Google Scholar] [CrossRef]
- Manurung, R.; Supriatna, A.; Esyanthi, R.R.; Putra, R.E. Bioconversion of Rice straw waste by black soldier fly larvae (Hermetia illucens L.): Optimal feed rate for biomass production. J. Entomol. Zool. Stud. 2016, 4, 1036–1041. [Google Scholar]
- Palma, L.; Fernandez-Bayo, J.; Niemeier, D.; Pitesky, M.; VanderGheynst, J.S. Managing high fiber food waste for the cultivation of black soldier fly larvae. npj Sci. Food 2019, 3, 15. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.C.; Li, C.; Chen, B.; Zhang, Y.; Hu, B.; Li, X.; Hu, W. Effects of Dietary Lipid and Salt Contents on Growth Performance of Black Soldier Fly Larvae. Feed Ind. 2021, 42, 58–64. [Google Scholar] [CrossRef]
- Haunerland, N.H.; Shirk, P.D. Regional and Functional Differentiation in the Insect Fact Body. Annu. Rev. Èntomol. 1995, 40, 121–145. [Google Scholar] [CrossRef]
- Dallerac, R.; Labeur, C.; Jallon, J.-M.; Knipple, D.C.; Roelofs, W.L.; Wicker-Thomas, C. A Delta 9 desaturase gene with a different substrate specificity is responsible for the cuticular diene hydrocarbon polymorphism in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2000, 97, 9449–9454. [Google Scholar] [CrossRef] [Green Version]
- Lam, S.M.; Zhou, T.; Li, J.; Zhang, S.; Chua, G.H.; Li, B.; Shui, G. A robust, integrated platform for comprehensive analyses of acyl-coenzyme As and acyl-carnitines revealed chain length-dependent disparity in fatty acyl metabolic fates across Drosophila development. Sci. Bull. 2020, 65, 1840–1848. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, J.; Wu, M.; Tong, H. Effects of palmitic acid on growth development and metabolism of glucose and lipid in Drosophila melanogaster. Modern Food Sci. Technol. 2021, 37, 114–119, 213. [Google Scholar] [CrossRef]
- Shi, D.; Han, T.; Chu, X.; Lu, H.; Yang, X.; Zi, T.; Zhao, Y.; Wang, X.; Liu, Z.; Ruan, J.; et al. An isocaloric moderately high-fat diet extends lifespan in male rats and Drosophila. Cell Metab. 2021, 33, 581–597.e9. [Google Scholar] [CrossRef] [PubMed]
- Zhan, S.; Fang, G.; Cai, M.; Kou, Z.; Xu, J.; Cao, Y.; Bai, L.; Zhang, Y.; Jiang, Y.; Luo, X.; et al. Genomic landscape and genetic manipulation of the black soldier fly Hermetia illucens, a natural waste recycler. Cell Res. 2020, 30, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Hoc, B.; Genva, M.; Fauconnier, M.L.; Lognay, G.; Francis, F.; Megido, R.C. About lipid metabolism in hermetia illucens (l. 1758): On the origin of fatty acids in prepupae. Sci. Rep. 2020, 10, 11916. [Google Scholar] [CrossRef]
- Champigny, C.M.; Cormier, R.P.J.; Simard, C.J.; St-Coeur, P.-D.; Fortin, S.; Pichaud, N. Omega-3 Monoacylglyceride Effects on Longevity, Mitochondrial Metabolism and Oxidative Stress: Insights from Drosophila melanogaster. Mar. Drugs 2018, 16, 453. [Google Scholar] [CrossRef] [Green Version]
- Groen, C.M.; Spracklen, A.J.; Fagan, T.N.; Tootle, T.L. Drosophila Fascin is a novel downstream target of prostaglandin signaling during actin remodeling. Mol. Biol. Cell 2012, 23, 4567–4578. [Google Scholar] [CrossRef]
- Li, X.; Song, L.; Chen, C.; Li, Y.; Wu, S. Influence of different fatty acids in artificial diets on growth, development and fecundity of Arma chinensis. Sci. Silva. Sin. 2018, 54, 85–93. [Google Scholar] [CrossRef]
- Chang, C.L.; Vargas, R.I. Wheat germ oil and Its effects on a liquid larval rearing diet for oriental fruit flies (Diptera: Tephritidae). J. Econ. Entomol. 2007, 100, 322–326. [Google Scholar] [CrossRef]
- Senyilmaz, D.; Virtue, S.; Xu, X.; Tan, C.Y.; Griffin, J.L.; Miller, A.; Vidal-Puig, A.; Teleman, A.A. Regulation of mitochondrial morphology and function by stearoylation of TFR1. Nature 2015, 525, 124–128. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.C. Insect Physiology; China Agriculture Press: Beijing, China, 2013. [Google Scholar]
- Wang, S.W.; Xu, B.H.; Wang, H. Appropriate Linoleic acid supplemental level in Apis mellifera ligustica worker larvae diet Chinese. J. Anim. Anim. Nutr. 2015, 27, 1440–1449. [Google Scholar]
- Vanderzant, E.S.; Kerur, D.; Retser, R. The Role of Dietary Fatty Acids in the Development of the Pink Bollworm. J. Econ. Èntomol. 1957, 50, 606–608. [Google Scholar] [CrossRef]
- Meneguz, M.; Schiavone, A.; Gai, F.; Dama, A.; Lussiana, C.; Renna, M.; Gasco, L. Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia illucens) larvae. J. Sci. Food Agric. 2018, 98, 5776–5784. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Tomberlin, J.K.; Brady, J.A.; Sanford, M.R.; Yu, Z. Black Soldier Fly (Diptera: Stratiomyidae) Larvae Reduce Escherichia coli in Dairy Manure. Environ. Èntomol. 2008, 37, 1525–1530. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Rehman, K.U.; Yu, Y.; Liu, X.; Wang, H.; Tomberlin, J.K.; Sze, S.-H.; Cai, M.; Zhang, J.; Yu, Z.; et al. De novo transcriptome sequencing and analysis revealed the molecular basis of rapid fat accumulation by black soldier fly (Hermetia illucens L.) for development of insectival biodiesel. Biotechnol. Biofuels 2019, 12, 194. [Google Scholar] [CrossRef]
- Wang, Z.L. Development of functional components of insect oil. Chn. Oils. Fats. 2003, 28, 33–34. [Google Scholar]
- Barragan-Fonseca, K.B.; Gort, G.; Dicke, M.; Van Loon, J.J.A. Effects of dietary protein and carbohydrate on life-history traits and body protein and fat contents of the black soldier fly Hermetia illucens. Physiol. Èntomol. 2019, 44, 148–159. [Google Scholar] [CrossRef] [Green Version]
- Matthäus, B.; Piofczyk, T.; Katz, H.; Pudel, F. Renewable Resources from Insects: Exploitation, Properties, and Refining of Fat Obtained by Cold-Pressing from Hermetia illucens (Black Soldier Fly) Larvae. Eur. J. Lipid Sci. Technol. 2019, 121, 121. [Google Scholar] [CrossRef]
- Spranghers, T.; Ottoboni, M.; Klootwijk, C.; Ovyn, A.; Deboosere, S.; De Meulenaer, B.; Michiels, J.; Eeckhout, M.; De Clercq, P.; De Smet, S. Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. J. Sci. Food Agric. 2017, 97, 2594–2600. [Google Scholar] [CrossRef]
- Dias, R.O.; Cardoso, C.; Pimentel, A.C.; Damasceno, T.F.; Ferreira, C.; Terra, W.R. The roles of mucus-forming mucins, peritrophins and peritrophins with mucin domains in the insect midgut. Insect Mol. Biol. 2017, 27, 46–60. [Google Scholar] [CrossRef]
- Giannetto, A.; Oliva, S.; Lanes, C.F.C.; de Araújo Pedron, F.; Savastano, D.; Baviera, C.; Parrino, V.; Lo Paro, G.; Spanò, N.C.; Cappello, T.; et al. Hermetia illucens (Diptera: Stratiomydae) larvae and prepupae: Biomass production, fatty acid profile and expression of key genes involved in lipid metabolism. J. Biotechnol. 2020, 307, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.; Frederich, M.; Megido, R.C.; Alabi, T.; Malik, P.; Uyttenbroeck, R.; Francis, F.; Blecker, C.; Haubruge, E.; Lognay, G.; et al. Insect fatty acids: A comparison of lipids from three Orthopterans and Tenebrio molitor L. larvae. J. Asia Pac. Èntomol. 2017, 20, 337–340. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Ingredients | Content (%) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Soybean meal | 95 | 90 | 95 | 90 | 95 | 90 | 95 | 90 | 95 | 90 | 95 | 90 |
| Linseed oil | 5 | 10 | ||||||||||
| Peanut oil | 5 | 10 | ||||||||||
| Coconut oil | 5 | 10 | ||||||||||
| Soybean oil | 5 | 10 | ||||||||||
| Lard oil | 5 | 10 | ||||||||||
| Fish oil | 5 | 10 | ||||||||||
| Nutrient composition of diets (dry weight) | ||||||||||||
| Crude protein (%) | 44.1 | 41.1 | 43.7 | 41.1 | 43.5 | 42.1 | 44 | 41.5 | 43.5 | 41.5 | 44.3 | 41.4 |
| Crude lipid (%) | 5.8 | 9.8 | 5.6 | 9.9 | 6.3 | 10.3 | 6.1 | 10.3 | 5.6 | 9.7 | 6.1 | 10.0 |
| Crude ash (%) | 5.9 | 5.5 | 5.8 | 5.6 | 5.9 | 5.6 | 5.9 | 5.6 | 5.9 | 5.7 | 5.8 | 5.6 |
| Gross energy (KJ/kg) | 18.7 | 19.6 | 18.1 | 19.6 | 18.8 | 19.9 | 18.8 | 19.6 | 18.1 | 19.0 | 18.8 | 19.6 |
| Lysine (%) | 2.50 | 2.35 | 2.52 | 2.32 | 2.52 | 2.38 | 2.52 | 2.40 | 2.48 | 2.32 | 2.48 | 2.39 |
| Methionine (%) | 0.55 | 0.52 | 0.54 | 0.53 | 0.55 | 0.51 | 0.58 | 0.54 | 0.52 | 0.50 | 0.56 | 0.49 |
| Arginine (%) | 2.89 | 2.82 | 2.88 | 2.85 | 2.85 | 2.85 | 2.91 | 2.85 | 2.88 | 2.82 | 2.90 | 2.82 |
| Ca (%) | 0.32 | 0.30 | 0.31 | 0.29 | 0.33 | 0.30 | 0.30 | 0.28 | 0.32 | 0.29 | 0.31 | 0.28 |
| P (%) | 0.60 | 0.56 | 0.58 | 0.56 | 0.60 | 0.57 | 0.59 | 0.56 | 0.59 | 0.56 | 0.58 | 0.56 |
| Cl (%) | 0.08 | 0.07 | 0.07 | 0.08 | 0.07 | 0.08 | 0.07 | 0.05 | 0.13 | 0.08 | 0.08 | 0.07 |
| Na (mg/kg) | 140 | 130 | 110 | 89 | 140 | 120 | 110 | 97 | 97 | 100 | 110 | 98 |
| Dietary Sources and Levels | C8:0 | C10:0 | C12:0 | C14:0 | C16:0 | C16:1n-7 | C18:0 | C18:1n-9 | C18:2n-6 | C18:3n-3 | C20:1n-9 | C20:5n-3 | C22:6n-3 | Others | SFA | MUFA | PUFA | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Linseed oil | 5% | - | - | - | 0.1 | 7.0 | 0.1 | 4.5 | 19.5 | 21.3 | 46.5 | 0.1 | - | - | 0.9 | 12.1 | 19.8 | 68.1 |
| 10% | - | - | - | 0.1 | 6.7 | 0.1 | 4.6 | 18.7 | 19.0 | 50.0 | 0.1 | - | - | 0.8 | 11.7 | 19.0 | 69.3 | |
| Peanut oil | 5% | - | - | - | 0.1 | 9.6 | 0.1 | 3.7 | 43.6 | 35.8 | 1.5 | 1.0 | - | - | 4.7 | 17.8 | 45.0 | 37.2 |
| 10% | - | - | - | 0.1 | 9.4 | 0.1 | 3.6 | 45.5 | 34.6 | 0.9 | 1.1 | - | - | 4.8 | 17.7 | 46.9 | 35.4 | |
| Coconut oil | 5% | 4.8 | 4.5 | 38.6 | 16.0 | 10.3 | - | 3.9 | 9.1 | 10.6 | 1.6 | 0.1 | - | - | 0.6 | 78.7 | 9.2 | 12.2 |
| 10% | 5.4 | 5.0 | 42.7 | 17.5 | 9.9 | - | 3.8 | 7.7 | 6.5 | 0.9 | 0.1 | - | - | 0.5 | 84.8 | 7.8 | 7.4 | |
| Soybean oil | 5% | - | - | - | 0.1 | 11.0 | 0.1 | 5.2 | 25.6 | 49.8 | 6.3 | 0.2 | - | - | 1.6 | 17.9 | 26.0 | 56.1 |
| 10% | 0.1 | 0.1 | - | 0.1 | 10.9 | 0.1 | 5.3 | 25.7 | 49.8 | 6.2 | 0.2 | - | - | 1.5 | 17.9 | 26.1 | 56.0 | |
| Lard oil | 5% | - | - | 0.1 | 1.0 | 24.9 | 1.7 | 11.0 | 37.2 | 20.2 | 2.2 | 0.5 | - | - | 1.4 | 37.5 | 39.5 | 23.0 |
| 10% | - | - | 0.1 | 1.0 | 25.9 | 1.8 | 11.4 | 38.5 | 18.1 | 1.3 | 0.5 | - | - | 1.3 | 39.0 | 41.0 | 20.1 | |
| Fish oil | 5% | - | - | 0.1 | 5.5 | 21.9 | 5.9 | 5.3 | 18.9 | 12.5 | 2.7 | 2.4 | 7.4 | 10.7 | 6.6 | 35.9 | 28.9 | 35.2 |
| 10% | - | - | 0.1 | 6.0 | 22.5 | 6.4 | 5.2 | 18.6 | 9.0 | 2.3 | 2.6 | 8.3 | 12.1 | 7.1 | 37.1 | 29.4 | 33.5 | |
| Growth Parameter | Fat Levels | Fat Sources | |||||
|---|---|---|---|---|---|---|---|
| Linseed Oil | Peanut Oil | Coconut Oil | Soybean Oil | Lard Oil | Fish Oil | ||
| Pre-pupa rate 1 (%) | 5% | 0.0 ± 0.0 aA | 0.0 ± 0.0 a | 0.0 ± 0.0 aA | 1.1 ± 0.7 a | 7.7 ± 0.3 bA | 3.2 ± 1.2 ab |
| 10% | 11.8 ± 1.2 dB | 0.0 ± 0.0 a | 5.7 ± 0.8 cB | 1.2 ± 1.2 b | 19.8 ± 1.6 eB | 6.2 ± 1.5 c | |
| Final body weight (mg/individual) | 5% | 59.5 ± 10.7 aA | 55.9 ± 7.8 aA | 103.7 ± 11.4 bA | 104.3 ± 7.3 b | 118.7 ± 13.8 c | 113.7 ± 17.5 cA |
| 10% | 71.4 ± 6.2 aB | 94.2 ± 9.5 bB | 115.5 ± 8.7 cB | 100.0 ± 9.7 b | 115.6 ± 14.4 c | 147.2 ± 13.9 dB | |
| Final body size (mm/individual) | 5% | 13.3 ± 1.1 b | 12.2 ± 0.7 a | 15.3 ± 0.7 bc | 15.3 ± 0.5 bc | 15.8 ± 0.7 cd | 16.0 ± 0.8 d |
| 10% | 13.0 ± 0.8 a | 15.5 ± 0.7 bc | 15.8 ± 0.8 c | 15.1 ± 0.6 b | 15.9 ± 0.8 c | 17.1 ± 0.7 d | |
| Models | R2 | p-Value |
|---|---|---|
| Pre-pupa rate = 0.879 × C18:0 content | 0.749 | <0.001 |
| Pre-pupa rate = 0.908 × C18:0 content + 0.298 × C18:3n-3 content | 0.829 | <0.001 |
| Pre-pupa rate = 0.927 × C18:0 content + 0.301 × C18:3n-3 content − 0.258 × C18:2n-6 content | 0.899 | <0.001 |
| Final body weight = 0.758 × C16:0 content | 0.531 | 0.004 |
| Fatty Acids | C12:0 | C14:0 | C16:0 | C16:1n-9 | C18:0 | C18:1n-9 | C18:2n-6 | C18:3n-3 | C20:5n-3 | Others | SFA | MUFA | PUFA | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Initial larvae | 0.7 a | 0.5 a | 12.9 bc | 2.2 c | 13.5 h | 29.5 h | 33.8 k | 3.3 d | 0.1 a | 3.4 f | 30.0 a | 32.5 h | 37.5 j | |
| Linseed oil | 5% | 23.9 e | 5.6 e | 12.6 b | 2.6 c | 4.5 ef | 17.3 d | 13.4 f | 18.2 e | - | 1.9 b | 47.6 e | 20.5 c | 31.9 i |
| 10% | 23.8 e | 4.9 d | 10.3 a | 1.3 a | 3.6 cd | 17.7 d | 13.6 f | 23.2 f | 0.1 a | 1.6 a | 43.6 c | 19.3 b | 37.1 j | |
| Peanut oil | 5% | 21.0 c | 4.8 c | 15.0 g | 2.5 c | 4.4 e | 30.0 h | 19.0 g | 0.7 ab | - | 2.7 e | 46.9 de | 33.4 i | 19.7 e |
| 10% | 17.4 b | 3.9 b | 14.0 ef | 1.5 a | 3.7 cd | 35.7 i | 20.9 h | 0.5 a | - | 2.5 de | 40.7 b | 37.9 j | 21.3 f | |
| Coconut oil | 5% | 38.7 i | 14.0 i | 14.4 ef | 4.3 e | 3.5 c | 13.4 b | 8.3 c | 0.8 ab | - | 2.6 de | 72.2 j | 18.7 b | 9.1 b |
| 10% | 44.9 j | 15.2 j | 13.2 cd | 3.7 d | 2.7 a | 11.9 a | 5.6 a | 0.6 a | - | 2.5 d | 77.3 k | 16.5 a | 6.2 a | |
| Soybean oil | 5% | 28.5 h | 5.8 f | 13.9 de | 1.9 b | 3.7 cd | 17.9 d | 24.1 i | 2.4 c | 0.1 a | 1.8 b | 53.1 f | 20.4 c | 26.5 g |
| 10% | 22.2 d | 4.9 d | 15.5 h | 1.9 b | 3.9 d | 21.2 e | 26.1 j | 2.5 c | 0.4 b | 1.5 a | 47.5 e | 23.6 d | 28.9 h | |
| Lard oil | 5% | 25.8 f | 5.9 f | 21.1 j | 3.7 d | 5.4 g | 24.8 f | 10.5 e | 0.7 ab | - | 2.3 c | 59.3 h | 29.2 f | 11.4 c |
| 10% | 25.5 f | 5.6 e | 21.7 j | 3.6 d | 4.6 f | 26.5 g | 9.9 d | 0.6 a | 0.1 a | 2.1 c | 58.4 g | 30.7 g | 10.9 c | |
| Fish oil | 5% | 27.4 g | 7.7 g | 19.3 i | 6.0 f | 3.7 cd | 16.6 c | 7.3 b | 1.0 b | 4.8 c | 6.2 g | 60.5 hi | 24.2 e | 15.3 d |
| 10% | 27.6 gh | 8.0 h | 19.6 i | 6.7 g | 3.2 b | 15.8 c | 5.5 a | 0.9 ab | 5.6 d | 7.1 h | 61.1 i | 24.4 e | 14.5 d | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Dong, Y.; Sun, Q.; Tan, X.; You, C.; Huang, Y.; Zhou, M. Growth and Fatty Acid Composition of Black Soldier Fly Hermetia illucens (Diptera: Stratiomyidae) Larvae Are Influenced by Dietary Fat Sources and Levels. Animals 2022, 12, 486. https://doi.org/10.3390/ani12040486
Li X, Dong Y, Sun Q, Tan X, You C, Huang Y, Zhou M. Growth and Fatty Acid Composition of Black Soldier Fly Hermetia illucens (Diptera: Stratiomyidae) Larvae Are Influenced by Dietary Fat Sources and Levels. Animals. 2022; 12(4):486. https://doi.org/10.3390/ani12040486
Chicago/Turabian StyleLi, Xiangce, Yewei Dong, Qiuxuan Sun, Xiaohong Tan, Cuihong You, Yanhua Huang, and Meng Zhou. 2022. "Growth and Fatty Acid Composition of Black Soldier Fly Hermetia illucens (Diptera: Stratiomyidae) Larvae Are Influenced by Dietary Fat Sources and Levels" Animals 12, no. 4: 486. https://doi.org/10.3390/ani12040486