
The Reproductive Success of Triturus ivanbureschi × T. macedonicus F1 Hybrid Females (Amphibia: Salamandridae)
by
 ,
,
Tijana Vučić
1,2,* ,
,
Ana Ivanović
1,
Maja Ajduković
2,
Nikola Bajler
1,2 and
Milena Cvijanović
2,* 1
Faculty of Biology, Institute of Zoology, University of Belgrade, Studentski trg 16, 11000 Belgrade, Serbia
2
Department of Evolutionary Biology, Institute for Biological Research “Siniša Stanković”, National Institute of the Republic of Serbia, University of Belgrade, Bulevar Despota Stefana 142, 11000 Belgrade, Serbia
*
Authors to whom correspondence should be addressed.
Animals 2022, 12(4), 443; https://doi.org/10.3390/ani12040443
Submission received: 24 December 2021
/
Revised: 10 February 2022
/
Accepted: 10 February 2022
/
Published: 12 February 2022
(This article belongs to the Special Issue Salamanders: Distribution, Diversity, and Conservation)
Abstract
:Simple Summary
Two moderately related large-bodied newt species endemic to the Balkan Peninsula, the Balkan crested newt (Triturus ivanbureschi) and the Macedonian crested newt (T. macedonicus), coexist and hybridize in central Serbia. Many generations of mutual hybrid crossings and backcrossings with parental species shaped the genetic composition of hybrid populations. Natural populations have admixed nuclear DNA (nuDNA) of parental species and T. ivanbureschi mitochondrial DNA (mtDNA), which is usually maternally inherited. The mechanisms that direct gene flow and shape the first generations of hybrids could explain the formation of hybrid zones and their maintenance in nature. We followed and compared life history traits related to reproduction of the first generation of reciprocal hybrids obtained by experimental crossing. Our results suggested that possible incompatibilities between mitochondrial and nuclear genomes, which could lead to the exclusion of T. macedonicus mtDNA in natural populations, most likely act at later stages of development or subsequent hybrid generations. Results from this study add to the growing knowledge of Triturus hybrid biology and ecology, which is the baseline for conservation programs necessary to protect these highly endangered amphibians.
Abstract
Two large-bodied newt species, Triturus ivanbureschi and T. macedonicus, hybridize in nature across the Balkan Peninsula. Consequences of hybridization upon secondary contact of two species include species displacement and asymmetrical introgression of T. ivanbureschi mtDNA. We set an experimental reciprocal cross of parental species and obtained two genotypes of F1 hybrids (with T. ivanbureschi or T. macedonicus mtDNA). When hybrids attained sexual maturity, they were engaged in mutual crossings and backcrossing with parental species. We followed reproductive traits over two successive years. Our main aim was to explore the reproductive success of F1 females carrying different parental mtDNA. Additionally, we tested for differences in reproductive success within female genotypes depending on the crossing with various male genotypes (hybrids or parental species). Both female genotypes had similar oviposition periods, number of laid eggs and hatched larvae but different body and egg sizes. Overall reproductive success (percentage of egg-laying females and viability of embryos) was similar for both genotypes. The type of crossing led to some differences in reproductive success within female genotypes. The obtained results suggest that processes that led to exclusion of T. macedonicus mtDNA in natural populations may be related to the survival at postembryonic stages of F2 generation or reproductive barriers that emerged in subsequent hybrid generations.
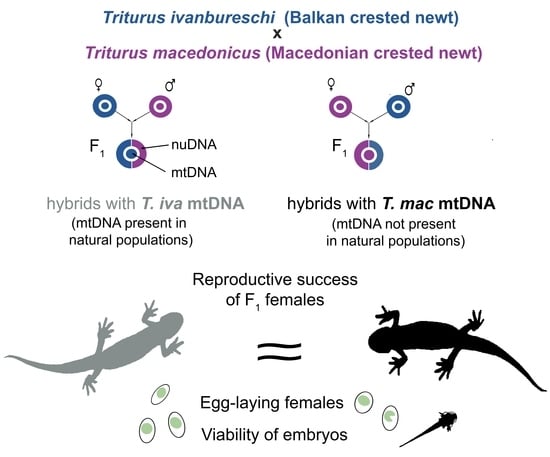
1. Introduction
The reproduction of genetically divergent taxa is a frequent phenomenon in natural populations (e.g., [1,2]). Hybrids could be sterile or produce further generations by mutual crossings and/or backcrossing with parental genotypes. The novel combination of genotypes in hybrids could be adaptive and beneficial for fitness, leading to hybrid speciation or these combinations could cause fitness loss [2,3,4,5,6,7,8]. The aforementioned outcomes of hybridization are largely dependent on the phylogenetic relatedness of parental species and/or the time of divergence from the most common ancestor. More closely related taxa, i.e., phylogenetically recent lineages, are expected to have viable hybrid offspring that could have adaptive advantages. The hybrid breakdown could be expected in the second or even in subsequent generations as a result of an accumulation of incompatibilities that produce reproductive barriers. On the other hand, hybridization between phylogenetically more divergent taxa could result in largely decreased viability or sterility of one or both sexes of F1 hybrids [6,9,10,11,12,13,14,15,16,17]. One of the hybridization consequences is asymmetrical gene flow resulting in introgression of mtDNA from one species to another, which could disturb mito-nuclear compatibility and potentially lead to substantial loss of hybrid fitness (e.g., [18,19,20]). There are several models that describe and explain the accumulation of genetic incompatibilities over time. For mito-nuclear incompatibilities, the Dobzhansky–Muller model was proposed as the most probable model of the evolution of incompatibilities (see [18] and references therein).
Phylogenetic relations within large-bodied newts (Triturus, Salamandridae) and a lack of complete reproductive barriers among species, with a wide range of hybridization outcomes, provide a substantial base for evolutionary studies of hybridization consequences. This monophyletic genus consists of two major groups, marbled and crested newts, which diverged from each other around 24 mya [21]. The marbled newts are T. marmoratus and T. pygmeus, while within crested newts, four groups are recognized based on genetic data and ecomorphological characteristics: (1) T. ivanbureschi, T. anatolicus, T. karelinii; (2) T. carnifex, T. macedonicus; (3) T. cristatus; and (4) T. dobrogicus [22]. The species have mostly parapatric distribution throughout Europe and parts of adjacent Asia. In the zones of species contact, interspecific hybridization occurs [23], resulting in dynamic hybrid zones in space and time. The movement of a hybrid zone implies advantages of one species at the expense of the other, which eventually leads to species spatial displacement. Most often, this scenario includes asymmetrical introgression of mtDNA from outcompeted to colonizing species [24,25,26,27,28,29,30,31]. Hybridization between Triturus species was also confirmed between autochthonous species and anthropogenically introduced species in areas well outside their natural range [32,33,34].
Natural hybrid populations could consist mostly of F1 hybrid generation, as in the case of two genetically well-separated species with different ecology and morphology, T. cristatus and T. marmoratus, which hybridize in western France (see [35] and references therein). These hybrids express typical heterosis; they are larger than the parental species and mostly sterile [36]. The opposite example is that of T. ivanbureschi × T. macedonicus hybrid populations on the Balkan Peninsula [27,37], which consist of an unknown generation of hybrid individuals derived from a long history of mutual hybrid crossings and backcrossing with both parental species [27,38,39]. Triturus ivanbureschi × T. macedonicus hybrids have intermediate body lengths related to parental species, and they are morphologically largely similar to both parental species [39]. In central Serbia, all populations of T. macedonicus, as well as T. ivanbureschi × T. macedonicus hybrids, have T. ivanbureschi mtDNA [25,26,27]. Asymmetrical mtDNA introgression is hypothesized to be a consequence of T. macedonicus range expansion over the range of T. ivanbureschi [25]. Triturus ivanbureschi and T. macedonicus are two moderately related, morphologically divergent species [22]. They diverge in body shape during postembryonic, larval development [40,41] as well as in the postmetamorphic period [42,43]. These species reproduce in similar aquatic habitats [44] but differ in reproductive traits, where T. macedonicus lay more eggs and have greater embryonic survival [43,45].
The mito-nuclear mismatch in Triturus populations in the central Balkans caused confusion in interpretations of species distribution and taxonomy (see [37] and references therein). In Serbia, many populations were considered to be T. ivanbureschi (syn. T. karelinii) according to mtDNA analysis, or T. macedonicus (syn. T. carnifex), according to morphology, were later confirmed as hybrid populations. Therefore, in previous studies, the reproductive data of some species are actually data for hybrid populations (Pirot [46,47]; Đurđevac [47]). These hybrid populations consist of T. ivanbureschi × T. macedonicus individuals of unknown generation (Fn) with T. ivanbureschi mtDNA [27]. Body and egg sizes of hybrid females [46,47] were similar to those reported for parental species [43,46,48].
The first generation of T. ivanbureschi × T. macedonicus hybrids has not yet been confirmed in nature [27,38,39]. Since processes occurring in the F1 generation are of special interest for an explanation of forming and maintenance of hybrid zones [49], we set up experimental reciprocal crossings to obtain two types of hybrid genotypes: T. ivanbureschi-mothered hybrids (with T. ivanbureschi mtDNA, confirmed in natural populations) and T. macedonicus-mothered hybrids (with T. macedonicus mtDNA, not confirmed in natural populations). Our main goal was to explore the eventual differences in reproductive success of reciprocal hybrids carrying different parental mtDNA. Since growth and reproduction in newts are negatively interconnected due to resource allocation [50,51] and the period between the first and second reproductions is characterized by considerable growth [52], we followed life history traits in the first two consecutive years of reproduction. We recorded the life history traits directly related to reproductive output: body size (length and mass), the number of egg-laying females, duration of oviposition, the number and size of eggs and the number of hatchlings. As an estimation of reproductive success, we calculated the percentage of egg-laying females and the viability of their embryos. We also tested for eventual differences in reproductive success when hybrid females were bred with the hybrid males (with the same or reciprocal mtDNA) and with the males of parental species T. ivanbureschi and T. macedonicus (backcross combinations). Overall, we expected that hybrids with T. macedonicus mtDNA would have at least lower reproductive success, considering that T. macedonicus mtDNA is not present in natural populations. Failure in their reproductive success would indicate early exclusion of T. macedonicus mtDNA in the first generations of crossings upon T. ivanbureschi and T. macedonicus secondary contact.
2. Materials and Methods
Individuals of T. ivanbureschi and T. macedonicus were collected from natural populations away from their contact zones: T. ivanbureschi from Zli Dol, Serbia (42°25 N; 22°27 E) with permission obtained from the Serbian Ministry of Energy, Development and Environmental Protection (permit No. 353-01-75/2014-08) and T. macedonicus in Ceklin, Montenegro (42°21 N; 18°59 E) with permission obtained from the Agency for Environmental Protection, Montenegro (permit No. UPI-328/4). Genetic data confirm that these were T. ivanbureschi and T. macedonicus populations [27]. A series of crossbreeding experiments were carried out at the Institute for Biological Research “Siniša Stanković” and approved by the Ethics Committee of the Institute (decision Nos. 03-03/16 and 01-1949). By crossing parental species, two genotypes of hybrids (HI—with T. ivanbureschi mtDNA and HM—T. macedonicus mtDNA) were obtained in two different years:
- HI in 2016: T. ivanbureschi ♀ (n = 8) × T. macedonicus ♂ (n = 5);
- HM in 2017: T. macedonicus ♀ (n = 4) × T. ivanbureschi ♂ (n = 4).
Since growth and survival, as well as reproductive traits, can be affected by external factors (e.g., temperature, density, availability of food) and their mutual interactions (e.g., [53]), we kept the experimental settings and conditions the same throughout different years of the experiment. Crossing of parental species was done outdoors in 500 L plastic vats (separate vat for each breeding crossing). Plastic strips were provided for egg deposition. When females started laying eggs, they were transferred to the laboratory in separate 10 L aquariums half-filled with dechlorinated tap water. Eggs, embryos and larvae were raised in the same controlled experimental conditions in both years. The temperature was kept constant (18–19 °C) with a natural day/night regime. Eggs and embryos were raised in Petri dishes (up to 10 eggs/embryos per dish). Dechlorinated tap water was changed every other day. All Petri dishes were checked daily to remove non-developing eggs and arrested embryos. Larvae were raised in 2 L plastic containers (single larva per container). Containers were half-filled with dechlorinated tap water, which was changed every other day. Larvae were fed ad libitum with Artemia sp. and Tubifex sp. After metamorphosis, animals hibernated during winter in a cold chamber at a constant temperature (4 °C). During the first postmetamorphic year, between first and second hibernation, juveniles were kept in 200 L plastic vats placed outdoors with a similar number of animals per vat. Plastic vats were enriched with perforated bricks, providing shelter and plastic lids, which were used as floating platforms. Newts were fed Tubifex sp. and Lumbricus sp. twice a week.
When hybrids had developed secondary sexual characters during the second postmetamorphic year, they were engaged in breeding series of mutual crossings and backcrossings with parental species (Figure 1). Animals hibernated before each breeding. HI females mated in 2018 (first reproduction) and 2019 (second reproduction). HM females mated in 2019 (first reproduction) and 2020 (second reproduction). The numbers of females involved in the breeding series were 23 HI and 20 HM in the first reproductive year and 25 HI and 20 HM in the second year of reproduction. To exclude the possibility of spermatozoid retention [54], the same females were crossed with the same males in two consecutive years. The only exceptions are two HI females, which were included in the crossing with HM males. Each crossing (see Figure 1) was conducted in a separate 200 L plastic container. Containers were set in the backyard of the Institute in semi-natural conditions. After females started laying eggs, they were transferred to separate 10 L aquariums, half-filled with dechlorinated water. Eggs and embryos were raised in the same controlled experimental conditions (see above). For more details of animals’ housing and experimental conditions, see [40,41,52].
Data for this study were collected during two consecutive years, in 2018 and 2019 for HI and in 2019 and 2020 for HM. To obtain measures of body size, females were photographed and weighed each year right after hibernation. Imaging was performed using a camera (Nikon D7100 or Sony DSC-F828) on a fixed stand with millimeter paper as a background. Body length (snout to vent length—SVL) was measured as the distance between the tip of the snout and the level of the posterior edge of the hind legs from the dorsal view using ImageJ software v. 1.50i [55]. Body mass (BM) was weighted to the nearest 0.01g using an electronic scale (MP300, Chyo Balance Corporation, Kyoto, Japan).
The eggs of newts consist of vitellus and a protective jelly layer. Females deposit individual eggs and wrap them in submerged vegetation [56], thus providing transparent plastic strips for egg deposition enabled easy observation of newly laid eggs. To estimate the number of laid eggs and egg size, we collected eggs on a daily basis and photographed them with a Nikon Digital Sight Fi2 camera attached to a Nikon SMZ800 stereo zoom microscope immediately after removal from the plastic strips for measurements. The number of eggs within each genotype was calculated from the first laid egg in common containers to the last laid egg in separate aquaria. Egg dimensions (maximum width of vitellus; maximum length and width of jelly) were taken using ImageJ software. Based on these measures, we calculated the volume of the vitellus and the volume of the entire egg. We calculated vitellus volume as the volume of a sphere: , where r is the vitellus width/2. For the volume of egg, we measured ellipsoid volume: where r1 and r2 are the radii length/2 and width of egg/2, respectively. The volume of the jelly was calculated as a difference between the volume of egg and the volume of vitellus: For statistical analysis of egg size, we used 100 eggs per genotype per year, which were taken randomly (RAND function in Microsoft Excel).
We also observed changes in other reproductive traits between the two reproductions, such as the number of egg-laying females, duration of oviposition, the total number of laid eggs and the total number of hatched larvae. Duration of oviposition was estimated as the number of days between the first and last laid egg within each genotype. To estimate reproductive success, we calculated the percentage of egg-laying females and the viability of their embryos. The percentage of egg-laying females was calculated as the ratio between the number of egg-laying females and the total number of females involved in breeding per hybrid genotype multiplied by 100. The viability of embryos was calculated as the ratio between the numbers of hatched larvae and the deposited eggs per hybrid genotype multiplied by 100.
Hybrid females of both genotypes were subdivided into four breeding groups exposed to different males (see Figure 1). As the exposure of females to different males can affect reproductive success in amphibians (e.g., [57]), we compared the percentages of egg-laying females and the viability of embryos separately within HI and HM females.
We used separated repeated measures ANOVAs to test for differences in the average body size (SVL and BM) and egg size (vitellus and jelly volumes), with female genotype as the categorical factor and year of reproduction as the within-subject (repeated measure) factor on the measured variables. Tukey’s HSD post hoc test was used to determine the statistical significance of between-group differences. Other reproductive traits (duration of oviposition, total number of eggs and total number of hatched larvae) were compared by the Kruskal–Wallis test since their values did not meet the criteria for ANOVA (assumptions of normality and sphericity). Differences in reproductive success were tested using differences between two proportions. Statistical analyses were done using Statistica10 software (StatSoft Inc., Tulsa, OK, USA, 2011).
3. Results
The repeated measures ANOVA showed that reproductive year (SVL: F1,38 = 317.53, p < 0.0001; BM: F1,38 = 109.94, p < 0.0001), genotype (SVL: F1,38 = 15.34, p = 0.004; BM: F1,38 = 10.95, p = 0.002) and their interaction (SVL: F1,38 = 36.99, p < 0.0001; BM: F1,38 = 14.57, p = 0.0005) had a significant effect on the increase of body size between two reproductive years. Post-hoc tests showed a significant ontogenetic increase in body size (SVL and BM) within both genotypes. Differences in body size between genotypes were evident only in the second year of reproduction (Figure 2, Table 1).
For the volume of vitellus, repeated measures ANOVA (Figure 2) showed a significant effect of reproductive year (F1,198 = 62.10, p < 0.0001) and year × genotype interaction (F1,198 = 27.17, p < 0.0001), but non-significant effect of genotype alone (F1,198 = 0.08, p = 0.7714). Volume of jelly was significantly affected by both factors, reproductive year (F1,198 = 34.92, p < 0.0001) and genotype (F1,198 = 21.41, p < 0.0001), as well as their interaction (F1,198 = 7.28, p = 0.0076). Post-hoc tests revealed that there was no ontogenetic difference in the volume of vitellus and jelly within HI, while HM eggs were significantly larger in the second reproductive year. HI had larger eggs (volume of vitellus and jelly) than HM in the first reproductive year. In the second reproductive year, HM had a larger volume of vitellus but a similar jelly volume to HI (Figure 2, Table 1). Kruskal–Wallis analyses showed that there were no differences in other reproductive traits (duration of oviposition, total number of laid eggs and total number of hatched larvae) between and within hybrid genotypes (p > 0.05 in all comparisons, Figure 2).
As the percentage of egg-laying females and viability of embryos also did not differ between HI and HM females in both reproductive years (p > 0.05 in all comparisons, Figure 3), we tested for differences in reproductive success between types of crossings (male effect) within each hybrid genotype. The percentage of egg-laying females did not differ within HI in both reproductive years regardless of different types of crossings, i.e., exposure to males of various genotypes. Within the HM females, the crossing of HM ♀ × ♂ had a relatively low percentage of egg-laying females, which was significantly lower from backcrossing of HM females with males of both parental species (T. ivanbureschi and T. macedonicus) in the first year, but this difference was lost in the second reproductive year (Table 2 and Table 3).
Offspring were obtained from all available breeding crossings in both reproductive years (see Figure 1). The viability of embryos obtained by different hybrid crossings and backcrossing was similar in the first reproductive year for both HI and HM females. For HI females, embryos obtained from the backcrossing of HI ♀ × T. macedonicus ♂ had greater viability than from the backcrossing of HI ♀ × T. ivanbureschi ♂. Embryos obtained from the reciprocal hybrid crossing (HI ♀ × HM ♂) had greater viability than those obtained from all other crossings of HI females and different male genotypes (HI, T. ivanbureschi and T. macedonicus) (Table 2 and Table 3). For HM females, embryos obtained from the crossing of HM ♀ × ♂ in the second reproductive year had the largest viability, significantly different from the crossing of reciprocal hybrids (HM ♀ × HI ♂), as well as from the backcrossing of HM females with T. ivanbureschi males (Table 2 and Table 3).
4. Discussion
In natural populations, overall reproductive output and success of F1 hybrid generation, particularly divergence of reciprocal hybrids, have an important impact on the long-term consequence of hybridization, as they can alter the direction of gene flow [58,59]. Therefore, the mito-nuclear incompatibility could shape patterns of genetic variation in hybrid zones and impact their dynamics. Triturus newts are a suitable model system for studies of mechanisms that lead to asymmetrical direction of mtDNA and introgression. Hybridization of the phylogenetically distant T. cristatus and T. marmoratus resulted in mtDNA introgression asymmetry due to divergence in the reproductive success of reciprocal crossings. More than 90% of individuals in natural populations have T. cristatus mtDNA [60], while F2 and further hybrid generations are rare [35,60]. Hybridization of T. ivanbureschi and T. macedonicus, two moderately related species, also resulted in an asymmetrical mtDNA introgression. Triturus ivanbureschi mtDNA is found across populations of T. ivanbureschi, T. macedonicus and T. ivanbureschi × T. macedonicus, in the Balkan Peninsula with the loss of T. macedonicus mtDNA in central Serbia [25,26,27].
Studies of T. ivanbureschi × T. macedonicus hybridization showed that hybrids have intermediate morphology compared to parental genotypes [39,40,41]. Hybridization affected physiological response resulting in raised oxidative stress parameters in hybrid larvae, which might have a negative impact on their survival [61,62,63]. However, fitness consequences of hybridization comprising slower growth rate, sex ratio disturbance and lower survival were not evident in F1 hybrids compared to maternal species during the early postmetamorphic period [52]. The morphological divergences between reciprocal hybrids were not recorded in previous morphological studies of larvae and recently metamorphosed juveniles. They have similar developmental patterns and growth rates [40,41].
We found that, at the beginning of the second postmetamorphic year, both F1 hybrids showed secondary sexual characters but did not diverge in body size. The divergence in size between reciprocal hybrids was recorded after the first reproduction, which coincides with the timing of sexual size dimorphism and divergence in size between hybrids and maternal species [52].
The other significant difference between reciprocal hybrids is related to egg size, which can have a profound impact on embryonic development and survival, as well as on larval growth and developmental rates [64]. Hybrids with T. ivanbureschi mtDNA laid larger eggs in the first reproduction (no differences between reciprocal hybrids in female body size), while eggs of hybrids with T. macedonicus mtDNA had substantially larger vitellus in the second reproduction, despite significantly smaller body size compared to hybrid females with T. ivanbureschi mtDNA. Additionally, reciprocal hybrids differed in the pattern of change in egg size between the first and second reproductions, where hybrids with T. macedonicus mtDNA showed a greater increase in vitellus size (see Figure 2). It is considered that the size of an amphibian egg is positively correlated with female body size. Thus, it is expected that older and larger females produce larger eggs, i.e., that ontogenetic changes of female body size should affect egg size [46,65,66]. Our results do not support previous notations. We suggest three mutually non-exclusive hypotheses that can be applied to explain the observed pattern of ontogenetic changes and divergences in vitellus size between reciprocal hybrids. First, the observed divergences may indicate that hybrids with T. ivanbureschi mtDNA invest more in somatic growth than in reproduction, while the opposite trend could be true for hybrids with T. macedonicus mtDNA, which could invest more in reproduction by enlarging vitellus volume. Second, our results could indicate that egg size is a heritable trait that is inherited by maternal investments through egg cytoplasm. In newts, egg size is a species-specific trait [47]. In a comparison of parental species, T. macedonicus has larger eggs than T. ivanbureschi [43]. The cytoplasm of the oocyte is full of maternal cytoplasmic components subsequently present in each cell of the new embryo [67], having an important role in the determination of egg size and possibly leading to significant divergences in egg size between two reciprocal hybrids. The third possible explanation for the divergence in egg size and female growth could be different sensitivity to external conditions of reciprocal hybrids. Previously, it was proposed that T. macedonicus is a thermophilic species [43,68]. Vitellogenesis occurs way before the breeding season, usually before hibernation [64], and it is dependent on different external and internal factors (e.g., [56]). It could be possible that external conditions were less suitable for F1 hybrids with T. macedonicus mtDNA at the time of their preparation for the first reproduction and vitellogenesis, i.e., hybrids with T. macedonicus mtDNA could be more susceptible to environmental variation during these periods. Studies on the plasticity of egg size and early life-history traits under different environmental conditions could give insight into the sensitivity of different genotypes to environmental conditions, with emphasis on different mtDNA groups. Hybrid females with both T. ivanbureschi and T. macedonicus mtDNA deposited a similar number of eggs in two reproductive years during a similar oviposition period. The number of eggs that hybrid females laid was in the range reported for both parental species in the same experimental settings [43], as well as for females from T. ivanburechi × T. macedonicus natural population [46], suggesting similar reproductive potential to that of the parental species. The observed pattern is opposite to T. cristatus and T. marmoratus hybridization, where hybrids have higher reproductive potential compared to parental species in experimental conditions, but low viability of eggs [69].
In our study, hybrid females were bred with four male genotypes (see Figure 1). In some amphibian taxa, females tend to choose males with whom they will engage in mating [56,57,70,71], which could affect overall reproductive success by decreasing or increasing the percentage of females that are involved in active breeding. Moreover, in hybridizing species, females’ choice could be a prezygotic barrier causing unidirectional mtDNA exchange if females of one species mate with males of the other and not vice versa [71]. Considering hybrid mutual mating and mating with parental species, it has been proposed that intermediate phenotypes can be disadvantageous for hybrid males, as they could be less attractive to females [7]. This could be true for newts, as it was shown that females do prefer larger males with a prominent crest [72]. In our experimental settings, the absence of differences between reciprocal hybrids in the percentage of egg-laying females, i.e., females engaged in active reproduction, suggests that females did not refuse to breed with either male genotype and their pickiness did not represent a barrier for either mtDNA to be passed on to the next generation. In comparisons within each female genotype, slight differences in percentages of egg-laying females in the first reproduction were not recorded in the second year of reproduction. Possibly, in their first reproduction, females preferred older (i.e., larger) males with prominent crests [72], as their conspecific males were of the same age or hybrid males were less successful in their first year of reproduction.
One characteristic of Triturus newts is that half of embryos die during development at the tail bud stage due to balanced lethal syndrome [73]. Viable eggs that had developed to hatchlings were obtained from all types of crossings (Figure 1) with no difference in embryo mortality between females with T. ivanbureschi and T. macedonicus mtDNA. However, viability differed in the second reproduction within reciprocal hybrid females when a different type of crossing (i.e., different male genotype) was included. Differences in the viability of embryos from different crossings and backcrossings indicate a possible decrease in survival of the F2 generation. For example, the backcrossing of hybrid females with T. ivanbureschi mtDNA and T. macedonicus was more successful than the one including T. ivanbureschi males. The obtained result could back up the hypothesis of postglacial species displacement and T. macedonicus nuclear DNA spreading [25]. This hypothesis suggests that, throughout generations, subsequent backcrossing of hybrid females with T. ivanbuerschi mtDNA with males of T. macedonicus diminished T. ivanbureschi nuclear DNA, rebuilt T. macedonicus nuclear DNA and retained T. ivanbureschi mtDNA [25]. The difference in the viability of F2 generations should be confirmed and further investigated in a larger study that would include postembryonic developmental stages, as it was shown that asymmetry in the frequency of reciprocal hybrids emerged after embryonic development in T. cristatus × T. marmoratus hybridization [59].
5. Conclusions
We found that the reproductive traits and success of reciprocal T. ivanbureschi × T. macedonicus F1 hybrids (with T. ivanbureschi or T. macedonicus mtDNA) is largely similar, expressing some differences depending on trait, year of reproduction and male genotype involved in mating. Overall, the obtained results suggest that the loss of T. macedonicus mtDNA emerged in F2 or subsequent hybrid generations. Major cito-nuclear incompatibilities are often masked in the first generation and manifested in F2 or subsequent hybrid generations due to their accumulation, the phenomenon known as hybrid breakdown (e.g., [10,16]). The viability of F2 embryos, obtained from mutual hybrid crossings and backcrossing with both parental species, points out a possible decrease of fitness in the F2 generation. However, these results are only preliminary and should be tested further.
Females were monitored in the first two years of reproduction, so possibly results considering success in crossings with different male genotypes could change with the age of females. The postembryonic viability of F2 generation should be analyzed, as possible differences in survival can occur during larval development, metamorphosis or even during the postmetamorphic period. An experiment, which would include females and males of various ages of T. ivanbureschi, T. macedonicus and hybrids with both types of mtDNA, would enable females to choose between different male genotypes. The suggested experimental design could give another insight into processes that took place a long time ago, upon the initial secondary contact between two moderately related species of T. ivanbureschi and T. macedonicus.
Author Contributions
Conceptualization, T.V., A.I. and M.C.; methodology, T.V., A.I. and M.C.; software, M.C.; validation, T.V.; formal analysis, M.C. and T.V.; investigation, T.V., M.A., N.B. and M.C.; resources, T.V., A.I., M.A. and M.C.; data curation, T.V. and M.C.; writing—original draft preparation, T.V.; writing—review and editing, T.V., A.I., M.A., N.B. and M.C.; visualization, T.V. and A.I.; supervision, A.I. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Serbian Ministry of Education, Science and Technological Development, grant numbers 451-03-9/2021-14/ 200007 and 451-03-9/2021-14/ 200178. The APC was funded by the Institute for Biological Research “Siniša Stanković”, National Institute of the Republic of Serbia.
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Committee of the Institute for Biological Research ‘’Siniša Stanković”, National Institute of the Republic of Serbia decision Nos. 03-03/16 (date of approval: 1 April 2016) and 01-1949 (date of approval: 1 October 2018).
Data Availability Statement
Data is available upon request from the corresponding author.
Acknowledgments
We would like to thank S. Nikolić and J. Jovanović for their help in 2018. We thank M. Lakušić for her assistance in 2019. We also thank many undergraduate students of University of Belgrade—Faculty of biology for their technical assistance during the experiments. We would like to thank three anonymous reviewers for their constructive comments and suggestions and Enrico Lunghi for his editorial advices.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Barton, N.H.; Hewitt, G.M. Analysis of hybrid zones. Annu. Rev. Ecol. Syst. 1985, 16, 113–148. [Google Scholar] [CrossRef]
- Abbott, R.; Albach, D.; Ansell, S.; Arntzen, J.W.; Baird, S.J.; Bierne, N.; Boughman, J.; Brelsford, A.; Buerkle, C.A.; Buggs, R.; et al. Hybridization and speciation. J. Evol. Biol. 2013, 26, 229–246. [Google Scholar] [CrossRef] [Green Version]
- Arnold, M.L.; Hodges, S.A. Are natural hybrids fit or unfit relative to their parents? Trends Ecol. Evol. 1995, 10, 67–71. [Google Scholar] [CrossRef]
- Dowling, T.E.; Secor, C.L. The role of hybridization and introgression in the diversification of animals. Annu. Rev. Ecol. Syst. 1997, 28, 593–619. [Google Scholar] [CrossRef] [Green Version]
- Barton, N.H. The role of hybridization in evolution. Mol. Ecol. 2001, 10, 551–568. [Google Scholar] [CrossRef] [PubMed]
- Orr, H.A.; Turelli, M. The evolution of postzygotic isolation: Accumulating Dobzhansky-Muller incompatibilities. Evolution 2001, 55, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Coyne, J.A.; Orr, H.A. Speciation; Sinauer: Sunderland, MA, USA, 2004. [Google Scholar]
- Seehausen, O. Hybridization and adaptive radiation. Trends Ecol. Evol. 2004, 19, 198–207. [Google Scholar] [CrossRef]
- Endler, J.A. Geographic Variation, Speciation, and Clines; Princeton University Press: Princeton, NJ, USA, 1977. [Google Scholar]
- Burton, R.S. Hybrid breakdown in physiological response: A mechanistic approach. Evolution 1990, 44, 1806–1813. [Google Scholar] [CrossRef]
- Orr, H.A. The population genetics of speciation—The evolution of hybrid incompatibilities. Genetics 1995, 139, 1805–1813. [Google Scholar] [CrossRef]
- Burke, J.M.; Arnold, M.L. Genetics and the fitness of hybrids. Annu. Rev. Gen. 2001, 35, 31–52. [Google Scholar] [CrossRef] [Green Version]
- Burton, R.S.; Ellison, C.K.; Harrison, J.S. The sorry state of F2 hybrids: Consequences of rapid mitochondrial DNA evolution in allopatric populations. Am. Nat. 2006, 168, S14–S24. [Google Scholar] [CrossRef]
- Stelkens, R.; Seehausen, O. Genetic distance between species predicts novel trait expression in their hybrids. Evolution. 2009, 63, 884–897. [Google Scholar] [CrossRef] [Green Version]
- Arnold, M.L.; Martin, N.H. Hybrid fitness across time and habitats. Trends Ecol. Evol. 2010, 25, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Stelkens, R.B.; Schmid, C.; Seehausen, O. Hybrid breakdown in cichlid fish. PLoS ONE 2015, 10, e0127207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rometsch, S.J.; Torres-Dowdall, J.; Meyer, A. Evolutionary dynamics of pre- and postzygotic reproductive isolation in cichlid fishes. Philos. T. Roy. Soc. B. 2020, 375, 20190535. [Google Scholar] [CrossRef] [PubMed]
- Burton, R.S.; Barreto, F.S. A disproportionate role for mtDNA in Dobzhansky–Muller incompatibilities? Mol. Ecol. 2012, 21, 4942–4957. [Google Scholar] [CrossRef] [PubMed]
- Healy, T.M.; Burton, R.S. Strong selective effects of mitochondrial DNA on the nuclear genome. Proc. Natl. Acad. Sci. USA 2020, 117, 6616–6621. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, J.M.; Matute, D.R. The importance of intrinsic postzygotic barriers throughout the speciation process: Intrinsic barriers throughout speciation. Philos. T. Roy. Soc. B. 2020, 375, 20190533. [Google Scholar] [CrossRef]
- Steinfartz, S.; Vicario, S.; Arntzen, J.W.; Caccone, A. A Bayesian approach on molecules and behavior: Reconsidering phylogenetic and evolutionary patterns of the Salamandridae with emphasis on Triturus newts. J. Exp. Zool. Part B. 2007, 308, 139–162. [Google Scholar] [CrossRef] [PubMed]
- Wielstra, B.; McCartney–Melstad, E.; Arntzen, J.W.; Butlin, R.K.; Shaffer, H.B. Phylogenomics of the adaptive radiation of Triturus newts supports gradual ecological niche expansion towards an incrementally aquatic lifestyle. Mol. Phylogenet. Evol. 2019, 133, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Arntzen, J.W.; Wielstra, B.; Wallis, G.P. The modality of nine Triturus newt hybrid zones assessed with nuclear, mitochondrial and morphological data. Biol. J. Linn. Soc. 2014, 113, 604–622. [Google Scholar] [CrossRef] [Green Version]
- Arntzen, J.W.; Wallis, G.P. Restricted gene flow in a moving hybrid zone of the newts Triturus cristatus and T. marmoratus in western France. Evolution. 1991, 45, 805–826. [Google Scholar] [CrossRef]
- Wielstra, B.; Arntzen, J.W. Postglacial species displacement in Triturus newts deduced from asymmetrically introgressed mitochondrial DNA and ecological niche models. BMC Evol. Biol. 2012, 12, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wielstra, B.; Arntzen, J.W. Extensive cytonuclear discordance in a crested newt from the Balkan Peninsula glacial refugium. Biol. J. Linn. Soc. 2020, 130, 578–585. [Google Scholar] [CrossRef]
- Wielstra, B.; Burke, T.; Butlin, R.K.; Arntzen, J.W. A signature of dynamic biogeography: Enclaves indicate past species replacement. Proc. R. Soc. B Biol. Sci. 2017, 284, 20172014. [Google Scholar] [CrossRef] [Green Version]
- Wielstra, B.; Burke, T.; Butlin, R.K.; Avcı, A.; Üzüm, N.; Bozkurt, E.; Kurtuluş, O.; Arntzen, J.W. A genomic footprint of hybrid zone movement in crested newts. Evol. Lett. 2017, 1, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Wielstra, B. Historical hybrid zone movement: More pervasive than appreciated. J. Biogeogr. 2019, 46, 1300–1305. [Google Scholar] [CrossRef] [Green Version]
- Arntzen, J.W.; López-Delgado, J.; van Riemsdijk, I.; Wielstra, B. A genomic footprint of a moving hybrid zone in marbled newts. J. Zool. Syst. Evol. Res. 2021, 59, 459–465. [Google Scholar] [CrossRef]
- López-Delgado, J.; van Riemsdijk, I.; Arntzen, J.W. Tracing species replacement in Iberian marbled newts. Ecol. Evol. 2021, 11, 402–414. [Google Scholar] [CrossRef]
- Brede, E.G.; Thorpe, R.S.; Arntzen, J.W.; Langton, T.E.S. A morphometric study of a hybrid newt population (Triturus cristatus/T. carnifex): Beam Brook Nurseries, Surrey, UK. Biol. J. Linn. Soc. 2000, 70, 685–695. [Google Scholar] [CrossRef] [Green Version]
- Meilink, W.R.; Arntzen, J.W.; van Delft, J.J.; Wielstra, B. Genetic pollution of a threatened native crested newt species through hybridization with an invasive congener in the Netherlands. Biol. Conserv. 2015, 184, 145–153. [Google Scholar] [CrossRef] [Green Version]
- Wielstra, B.; Burke, T.; Butlin, R.K.; Schaap, O.; Shaffer, H.B.; Vrieling, K.; Arntzen, J.W. Efficient screening for ‘genetic pollution’ in an anthropogenic crested newt hybrid zone. Conserv. Genet. Resour. 2016, 8, 553–560. [Google Scholar] [CrossRef]
- Arntzen, J.W.; Jehle, R.; Wielstra, B. Genetic and morphological data demonstrate hybridization and backcrossing in a pair of salamanders at the far end of the speciation continuum. Evol. Appl. 2021, 14, 2784–2793. [Google Scholar] [CrossRef] [PubMed]
- Cogălniceanu, D.; Stănescu, F.; Arntzen, J.W. Testing the hybrid superiority hypothesis in crested and marbled newts. J. Zool. Syst. Evol. Res. 2020, 58, 275–283. [Google Scholar] [CrossRef]
- Vučić, T.; Tomović, L.; Ivanović, A. The distribution of crested newts in Serbia: An overview and update. Bull. Nat. Hist. Mus. 2020, 13, 237–252. [Google Scholar] [CrossRef]
- Wielstra, B.; Arntzen, J.W. Kicking Triturus arntzeni when it’s down: Large–scale nuclear genetic data confirm that newts from the type locality are genetically admixed. Zootaxa 2014, 3802, 381–388. [Google Scholar] [CrossRef]
- Arntzen, J.W.; Üzüm, N.; Ajduković, M.D.; Ivanović, A.; Wielstra, B. Absence of heterosis in hybrid crested newts. PeerJ. 2018, 6, e5317. [Google Scholar] [CrossRef]
- Vučić, T.; Vukov, T.D.; Tomašević Kolarov, N.; Cvijanović, M.; Ivanović, A. The study of larval tail morphology reveals differentiation between two Triturus species and their hybrids. Amphib–Reptilia 2018, 39, 87–97. [Google Scholar] [CrossRef]
- Vučić, T.; Sibinović, M.; Vukov, T.D.; Tomašević Kolarov, N.; Cvijanović, M.; Ivanović, A. Testing the evolutionary constraints of metamorphosis: The ontogeny of head shape in Triturus newts. Evolution. 2019, 73, 1253–1264. [Google Scholar] [CrossRef]
- Cvijanović, M.; Ivanović, A.; Kalezić, M.L.; Zelditch, M.L. The ontogenetic origins of skull shape disparity in the Triturus cristatus group. Evol. Dev. 2014, 16, 306–317. [Google Scholar] [CrossRef] [Green Version]
- Vučić, T.; Ivanović, A.; Nikolić, S.; Jovanović, J.; Cvijanović, M. Reproductive characteristics of two Triturus species (Amphibia: Caudata). Arch. Biol. Sci. 2020, 72, 321–328. [Google Scholar] [CrossRef]
- Džukić, G.; Vukov, T.D.; Kalezić, M.L. The Tailed Amphibians of Serbia; Serbian Academy of Science and Arts: Belgrade, Serbia, 2016. [Google Scholar]
- Cvijanović, M.; Ivanović, A.; Tomašević Kolarov, N.; Džukić, G.; Kalezić, M.L. Early ontogeny shows the same interspecific variation as natural history parameters in the crested newt (Triturus cristatus superspecies)(Caudata, Salamandridae). Contrib. Zool. 2009, 78, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Furtula, M.; Ivanović, A.; Džukić, G.; Kalezić, M.L. Egg size variation in crested newts from the western Balkans (Caudata, Salamandridae, Triturus cristatus superspecies). Zool. Stud. 2008, 47, 585–590. [Google Scholar]
- Furtula, M.; Todorović, B.; Simić, V.; Ivanović, A. Interspecific differences in early life-history traits in crested newts (Triturus cristatus superspecies, Caudata, Salamandridae) from the Balkan Peninsula. J. Nat. Hist. 2009, 43, 469–477. [Google Scholar] [CrossRef]
- Lukanov, S.; Tzankov, N. Life history, age and normal development of the Balkan-Anatolian crested newt (Triturus ivanbureschi Arntzen and Wielstra, 2013) from Sofia district. North-West. J. Zool. 2016, 12, 22–32. [Google Scholar]
- Nolte, A.W.; Tautz, D. Understanding the onset of hybrid speciation. Trends Genet. 2010, 26, 54–58. [Google Scholar] [CrossRef]
- Halliday, T.R.; Verrell, P.A. Body size and age in amphibians and reptiles. J. Herpetol. 1988, 22, 253–265. [Google Scholar] [CrossRef]
- Arntzen, J.W. A growth curve for the newt Triturus cristatus. J. Herpetol. 2000, 34, 227–232. [Google Scholar] [CrossRef]
- Bugarčić, M.; Ivanović, A.; Cvijanović, M.; Vučić, T. Hybridization and early postmetamorphic growth in Triturus macedonicus. Amphib–Reptilia 2022. [Google Scholar] [CrossRef]
- Semlitsch, R.D. Critical elements for biologically based recovery plans of aquatic-breeding amphibians. Conserv. Biol. 2002, 16, 619–629. [Google Scholar] [CrossRef] [Green Version]
- Eddy, S.L.; Vaccaro, E.A.; Baggett, C.L.; Kiemnec-Tyburczy, K.M.; Houck, L.D. Sperm mass longevity and sperm storage in the female reproductive tract of Plethodon shermani (Amphibia: Plethodontidae). Herpetologica. 2015, 71, 177–183. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Duellman, W.E.; Trueb, L. Biology of Amphibians; JHU Press: Baltimore, MD, USA, 1994. [Google Scholar]
- Reyer, H.U.; Frei, G.; Som, C. Cryptic female choice: Frogs reduce clutch size when amplexed by undesired males. Proc. R. Soc. B. 1999, 266, 2101–2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, M.L.; Ballerini, E.S.; Brothers, A.N. Hybrid fitness, adaptation and evolutionary diversification: Lessons learned from Louisiana irises. Heredity. 2012, 108, 159–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, C.M.; Rankin, K.J.; McLean, C.A.; Stuart-Fox, D. Maternal reproductive output and F1 hybrid fitness may influence contact zone dynamics. J. Evol. Biol. 2021, 34, 680–694. [Google Scholar] [CrossRef]
- Arntzen, J.W.; Jehle, R.; Bardakci, F.; Burke, T.; Wallis, G.P. Asymmetric viability of reciprocal-cross hybrids between crested and marbled newts (Triturus cristatus and T. marmoratus). Evolution 2009, 63, 1191–1202. [Google Scholar] [CrossRef]
- Prokić, M.D.; Despotović, S.G.; Vučić, T.Z.; Petrović, T.G.; Gavrić, J.P.; Gavrilović, B.R.; Radovanović, T.B.; Saičić, Z.S. Oxidative cost of interspecific hybridization: A case study of two Triturus species and their hybrids. J. Exp. Biol. 2018, 221, jeb182055. [Google Scholar] [CrossRef] [Green Version]
- Petrović, T.G.; Vučić, T.Z.; Nikolić, S.Z.; Gavrić, J.P.; Despotović, S.G.; Gavrilović, B.R.; Radovanović, T.B.; Faggio, C.; Prokić, M.D. The effect of shelter on oxidative stress and aggressive behavior in crested newt larvae (Triturus spp.). Animals 2020, 10, 603. [Google Scholar] [CrossRef] [Green Version]
- Prokić, M.D.; Petrović, T.G.; Despotović, S.G.; Vučić, T.; Gavrić, J.P.; Radovanović, T.B.; Gavrilović, B.R. The effect of short-term fasting on the oxidative status of larvae of crested newt species and their hybrids. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2021, 251, 110819. [Google Scholar] [CrossRef]
- Kaplan, R.H.; Cooper, W.S. The evolution of developmental plasticity in reproductive characteristics: An application of the” adaptive coin-flipping” principle. Am. Nat. 1984, 123, 393–410. [Google Scholar] [CrossRef]
- Kaplan, R.H.; Salthe, S.N. The allometry of reproduction: An empirical view in salamanders. Am. Nat. 1979, 113, 671–689. [Google Scholar] [CrossRef]
- Morrison, C.; Hero, J.M. Geographic variation in life-history characteristics of amphibians: A review. J. Anim. Ecol. 2003, 72, 270–279. [Google Scholar] [CrossRef] [Green Version]
- Maurel, M.C.; Kanellopoulos-Langevin, C. Heredity—Venturing beyond genetics. Biol. Reprod. 2008, 79, 2–8. [Google Scholar] [CrossRef]
- Litvinchuk, S.N.; Rosanov, J.M.; Borkin, L.J. Correlations of geographic distribution and temperature of embryonic development with the nuclear DNA content in the Salamandridae (Urodela, Amphibia). Genome 2007, 50, 333–342. [Google Scholar] [CrossRef]
- Arntzen, J.W.; Hedlund, L. Fecundity of the newts Triturus cristatus, T. marmoratus and their natural hybrids in relation to species coexistence. Ecography. 1990, 13, 325–332. [Google Scholar] [CrossRef]
- Wilbur, H.M.; Rubenstein, D.I.; Fairchild, L. Sexual selection in toads: The roles of female choice and male body size. Evolution 1978, 32, 264–270. [Google Scholar] [CrossRef] [Green Version]
- Wirtz, P. Mother species–father species: Unidirectional hybridization in animals with female choice. Anim. Behav. 1999, 58, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Halliday, T.R. The courtship of European newts: An evolutionary perspective. In The Reproductive Biology of Amphibians; Taylor, D.H., Guttman, S.I., Eds.; Plenum Press: New York, NY, USA, 1977; pp. 185–232. [Google Scholar]
- Wielstra, B. Balanced lethal systems. Curr. Biol. 2020, 30, R742–R743. [Google Scholar] [CrossRef]
Figure 1.
Schematic presentation of the experimental design. The F1 hybrid females with T. ivanbureschi mtDNA (HI) were crossed and backcrossed in 2018 and 2019. The F1 hybrid females with T. macedonicus mtDNA (HM) were crossed and backcrossed in 2019 and 2020. * Sexually mature HM males were available only in 2019, and therefore not involved in crossings with HI females in their first reproduction in 2018.
Figure 1.
Schematic presentation of the experimental design. The F1 hybrid females with T. ivanbureschi mtDNA (HI) were crossed and backcrossed in 2018 and 2019. The F1 hybrid females with T. macedonicus mtDNA (HM) were crossed and backcrossed in 2019 and 2020. * Sexually mature HM males were available only in 2019, and therefore not involved in crossings with HI females in their first reproduction in 2018.

Figure 2.
Ontogenetic differences in body size (length—SVL and mass—BM) and reproductive traits in the first two reproductive years of F1 hybrid females with T. ivanbureschi mtDNA, HI (gray) and with T. macedonicus mtDNA, HM (black). Values of each trait are represented as mean ± standard error per each genotype and year of reproduction.
Figure 2.
Ontogenetic differences in body size (length—SVL and mass—BM) and reproductive traits in the first two reproductive years of F1 hybrid females with T. ivanbureschi mtDNA, HI (gray) and with T. macedonicus mtDNA, HM (black). Values of each trait are represented as mean ± standard error per each genotype and year of reproduction.

Figure 3.
Differences in the percentage of egg-laying females and viability of embryos, as a measure of reproductive success during the first two reproductive years of F1 hybrid females with T. ivanbureschi mtDNA, HI (gray) and with T. macedonicus mtDNA, HM (black).
Figure 3.
Differences in the percentage of egg-laying females and viability of embryos, as a measure of reproductive success during the first two reproductive years of F1 hybrid females with T. ivanbureschi mtDNA, HI (gray) and with T. macedonicus mtDNA, HM (black).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Differences in body size and egg volume between F1 hybrid females with T. ivanbureschi mtDNA (HI) and T. macedonicus mtDNA (HM) during the first two reproductive years. Analyzed traits: SVL—snout to vent length, BM—body mass, Vv—vitellus volume, Vj—jelly volume. Level of significances of pairwise comparisons: ns = not significant.
Table 1.
Differences in body size and egg volume between F1 hybrid females with T. ivanbureschi mtDNA (HI) and T. macedonicus mtDNA (HM) during the first two reproductive years. Analyzed traits: SVL—snout to vent length, BM—body mass, Vv—vitellus volume, Vj—jelly volume. Level of significances of pairwise comparisons: ns = not significant.
| Comparisons | Females | Eggs | ||
|---|---|---|---|---|
| SVL | BM | Vv | Vj | |
| within genotypes (I vs. II year) | ||||
| HI | 0.0002 | 0.0002 | ns | ns |
| HM | 0.0002 | 0.0004 | <0.0001 | <0.0001 |
| between genotypes (HI vs. HM) | ||||
| I year | ns | ns | 0.0011 | <0.0001 |
| II year | 0.0002 | 0.0002 | 0.0051 | ns |
Table 2.
Reproductive success (percentages of egg-laying females and viability of embryos) of F1 hybrid females with T. ivanburechi mtDNA (HI) and T. macedonicus mtDNA (HM) exposed to different males (see Figure 1 for various breeding crossings and abbreviations). Sexually mature HM males were not available in 2018 and therefore not involved in crossings with HI females in their first reproductive year.
Table 2.
Reproductive success (percentages of egg-laying females and viability of embryos) of F1 hybrid females with T. ivanburechi mtDNA (HI) and T. macedonicus mtDNA (HM) exposed to different males (see Figure 1 for various breeding crossings and abbreviations). Sexually mature HM males were not available in 2018 and therefore not involved in crossings with HI females in their first reproductive year.
| Breeding Crossings (♀ × ♂) | Egg-Laying Females (%) | Viability (%) | ||
|---|---|---|---|---|
| I Year | II Year | I Year | II Year | |
| HI × HI | 22 | 43 | 26 | 20 |
| HI × HM | / | 50 | / | 35 |
| HI × TI | 14 | 100 | 8 | 18 |
| HI × TM | 57 | 100 | 33 | 25 |
| HM × HM | 20 | 100 | 19 | 35 |
| HM × HI | 80 | 100 | 20 | 23 |
| HM × TI | 100 | 100 | 26 | 26 |
| HM × TM | 100 | 60 | 23 | 27 |
Table 3.
Pairwise comparison of reproductive success (percentages of egg-laying females and viability of embryos) of F1 hybrid females with T. ivanbureschi mtDNA (HI) and T. macedonicus mtDNA (HM) exposed to different males (see Figure 1 for various breeding crossings and abbreviations). The comparisons were done within HI and HM females separately. Sexually mature HM males were not available in 2018 and therefore not involved in crossings with HI females in their first reproductive year. Level of significances of pairwise comparisons: ns = not significant.
Table 3.
Pairwise comparison of reproductive success (percentages of egg-laying females and viability of embryos) of F1 hybrid females with T. ivanbureschi mtDNA (HI) and T. macedonicus mtDNA (HM) exposed to different males (see Figure 1 for various breeding crossings and abbreviations). The comparisons were done within HI and HM females separately. Sexually mature HM males were not available in 2018 and therefore not involved in crossings with HI females in their first reproductive year. Level of significances of pairwise comparisons: ns = not significant.
| Compared Breeding Crossings (♀ × ♂) | Egg-Laying Females (%) | Viability (%) | ||
|---|---|---|---|---|
| I Year | II Year | I Year | II Year | |
| HI × HI vs. HI × TI | ns | ns | ns | ns |
| HI × HI vs. HI × TM | ns | ns | ns | ns |
| HI × TI vs. HI × TM | ns | ns | ns | 0.0310 |
| HI × HI vs. HI × HM | / | ns | / | 0.0002 |
| HI × TI vs. HI × HM | / | ns | / | <0.0001 |
| HI × TM vs. HI × HM | / | ns | / | 0.0140 |
| HM × HM vs. HM × TI | 0.0320 | ns | ns | <0.0001 |
| HM × HM vs. HM × TM | 0.0320 | ns | ns | ns |
| HM × TI vs. HM × TM | ns | ns | ns | ns |
| HM × HM vs. HM × HI | ns | ns | ns | 0.0160 |
| HM × TI vs. HM × HM | ns | ns | ns | ns |
| HI × HI vs. HM × HM | ns | ns | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vučić, T.; Ivanović, A.; Ajduković, M.; Bajler, N.; Cvijanović, M. The Reproductive Success of Triturus ivanbureschi × T. macedonicus F1 Hybrid Females (Amphibia: Salamandridae). Animals 2022, 12, 443. https://doi.org/10.3390/ani12040443
AMA Style
Vučić T, Ivanović A, Ajduković M, Bajler N, Cvijanović M. The Reproductive Success of Triturus ivanbureschi × T. macedonicus F1 Hybrid Females (Amphibia: Salamandridae). Animals. 2022; 12(4):443. https://doi.org/10.3390/ani12040443
Chicago/Turabian StyleVučić, Tijana, Ana Ivanović, Maja Ajduković, Nikola Bajler, and Milena Cvijanović. 2022. "The Reproductive Success of Triturus ivanbureschi × T. macedonicus F1 Hybrid Females (Amphibia: Salamandridae)" Animals 12, no. 4: 443. https://doi.org/10.3390/ani12040443
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.