
Occurrence of Rotavirus A Genotypes and Other Enteric Pathogens in Diarrheic Suckling Piglets from Spanish Swine Farms

 , , ,
, , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Approval Statement
2.2. Stool Samples
2.3. Nucleic Acid Extraction and qPCR or RT-qPCR
2.4. G and P Genotyping of RVA
2.5. Sequence Analysis
2.6. Nucleotide Sequence Accession Numbers
2.7. Statistical Analysis
3. Result
3.1. Detection of Enteropathogens
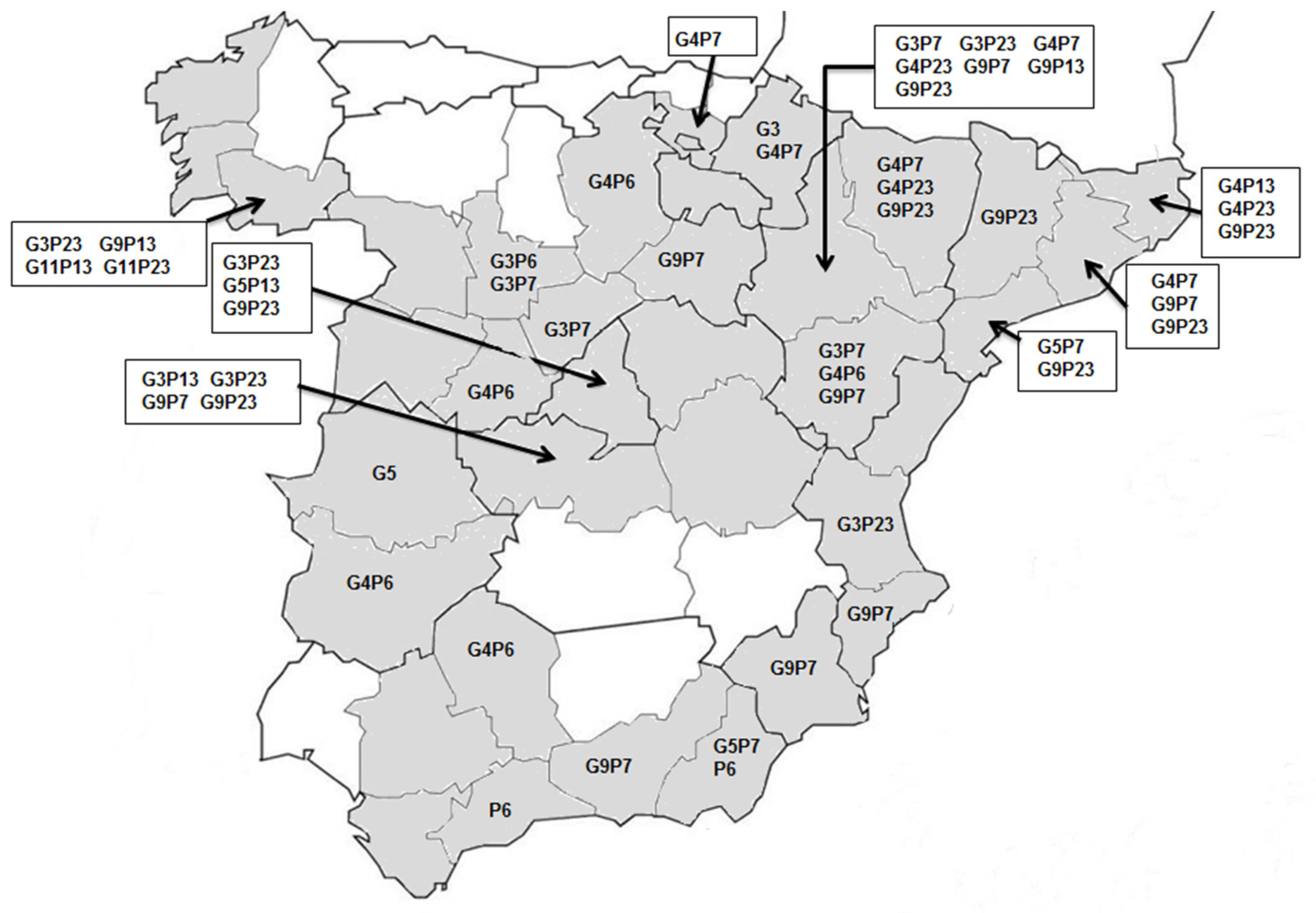
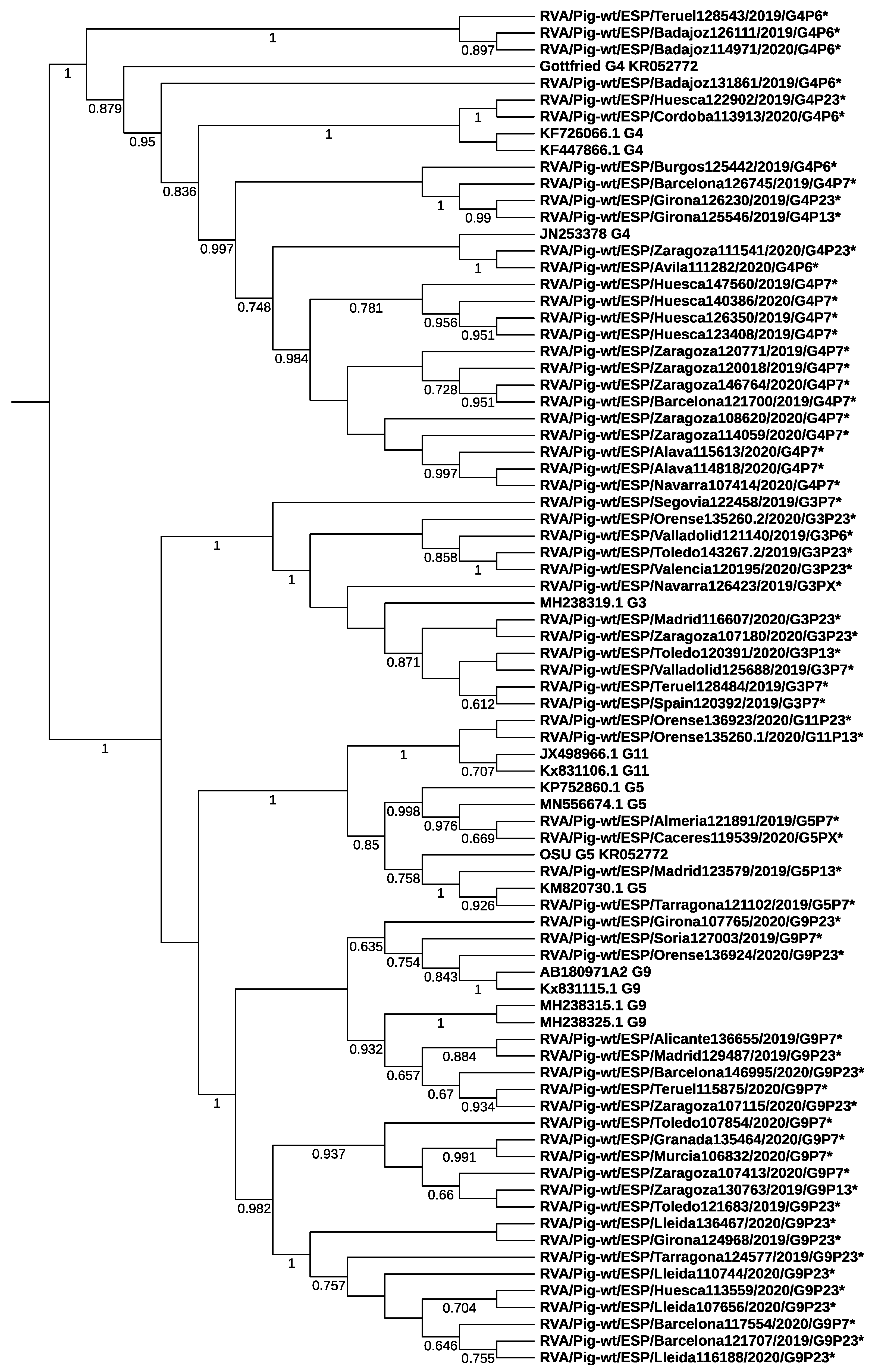
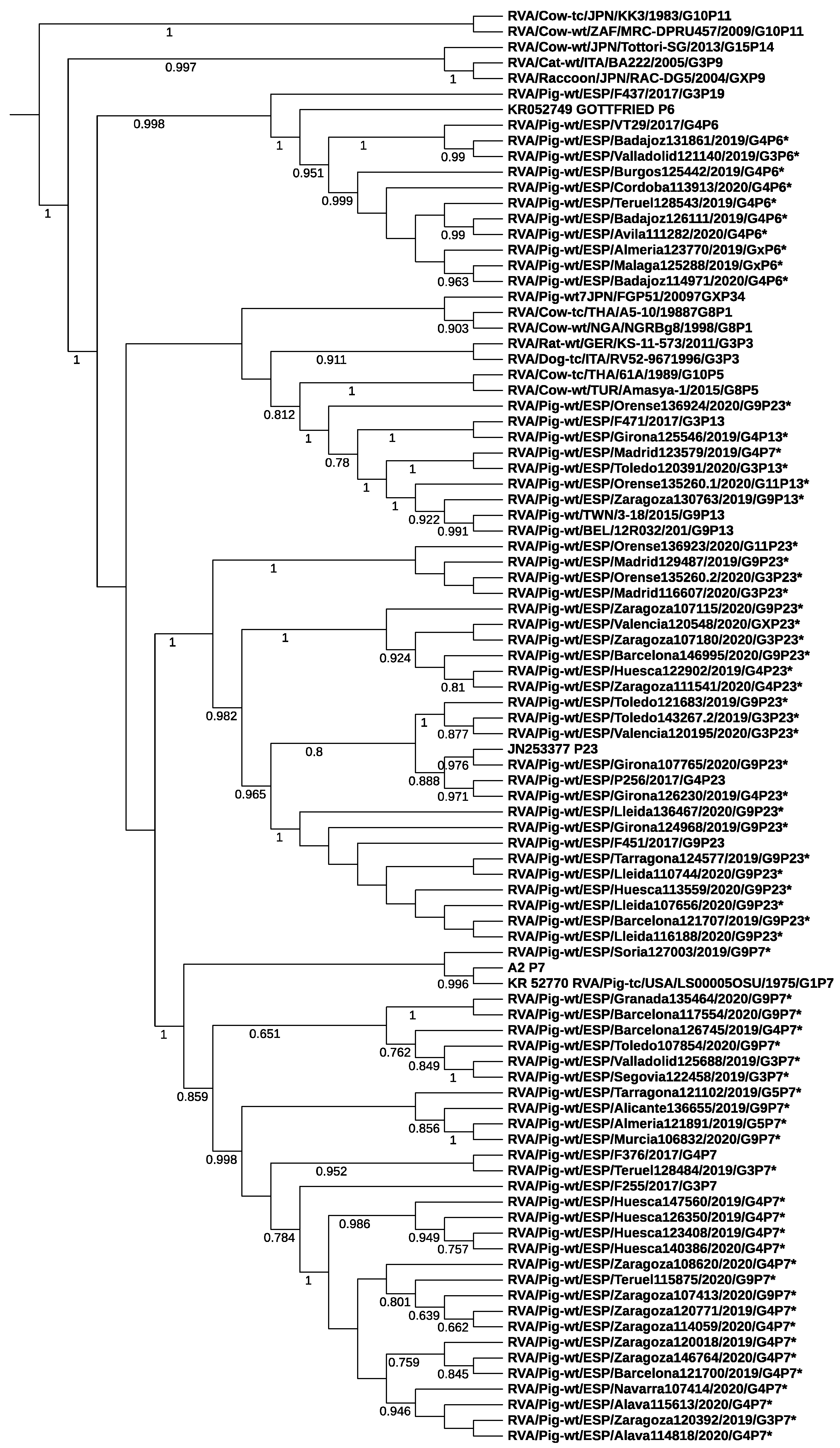
3.2. Rotavirus Genotypes
3.3. Differences among Field Isolates and Vaccine Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kongsted, H.; Stege, H.; Toft, N.; Nielsen, J.P. The effect of New Neonatal Porcine Diarrhoea Syndrome (NNPDS) on average daily gain and mortality in 4 Danish pig herds. BMC Vet. Res. 2014, 10, 90. [Google Scholar] [CrossRef]
- Westin, R.; Holmgren, N.; Hultgren, J.; Ortman, K.; Linder, A.; Algers, B. Post-mortem findings and piglet mortality in relation to strategic use of straw at farrowing. Prev. Vet. Med. 2015, 119, 141–152. [Google Scholar] [CrossRef]
- Sjölund, M.; Zoric, M.; Wallgren, P. Financial impact of disease on pig production: Part III. Gastrointestinal disorders. In Proceedings of the 6th European Symposium of Porcine Health Management, Sorrento, Italy, 8 May 2014; p. 189. [Google Scholar]
- Kongsted, H.; Pedersen, K.; Hjulsager, C.K.; Larsen, L.E.; Pedersen, K.S.; Jorsal, S.E.; Bækbo, P. Diarrhoea in neonatal piglets: A case control study on microbiological findings. Porc. Health Manag. 2018, 4, 17. [Google Scholar] [CrossRef]
- Mesonero-Escuredo, S.; Strutzberg-Minder, K.; Casanovas, C.; Segalés, J. Viral and bacterial investigations on the aetiology of recurrent pig neonatal diarrhoea cases in Spain. Porc. Health Manag. 2018, 4, 5. [Google Scholar] [CrossRef] [PubMed]
- Estes, M.K.; Cohen, J. Rotavirus gene structure and function. Microbiol. Rev. 1989, 53, 410–449. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, A.N.; Amimo, J.O.; Saif, L.J. Porcine rotaviruses: Epidemiology, immune responses and control strategies. Viruses 2017, 9, 48. [Google Scholar] [CrossRef] [PubMed]
- Vidal, A.; Martín-Valls, G.E.; Tello, M.; Mateu, E.; Martín, M.; Darwich, L. Prevalence of enteric pathogens in diarrheic and non-diarrheic samples from pig farms with neonatal diarrhea in the North East of Spain. Vet. Microbiol. 2019, 237, 108419. [Google Scholar] [CrossRef]
- Marthaler, D.; Homwong, N.; Rossow, K.; Culhane, M.; Goyal, S.; Collins, J.; Matthijnssens, J.; Ciarlet, M. Rapid detection and high occurrence of porcine rotavirus A, B, and C by RT-qPCR in diagnostic samples. J. Virol. Methods 2014, 209, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Moutelíková, R.; Prodělalová, J.; Dufková, L. Prevalence study and phylogenetic analysis of group C porcine rotavirus in the Czech Republic revealed a high level of VP6 gene heterogeneity within porcine cluster I1. Arch. Virol. 2014, 159, 1163–1167. [Google Scholar] [CrossRef]
- Midgley, S.E.; Bányai, K.; Buesa, J.; Halaihel, N.; Hjulsager, C.K.; Jakab, F.; Kaplon, J.; Larsen, L.E.; Monini, M.; Poljšak-Prijatelj, M.; et al. Diversity and zoonotic potential of rotaviruses in swine and cattle across Europe. Vet. Microbiol. 2012, 156, 238–245. [Google Scholar] [CrossRef]
- Papp, H.; László, B.; Jakab, F.; Ganesh, B.; De Grazia, S.; Matthijnssens, J.; Ciarlet, M.; Martella, V.; Bányai, K. Review of group A rotavirus strains reported in swine and cattle. Vet. Microbiol. 2013, 165, 190–199. [Google Scholar] [CrossRef]
- Matthijnssens, J.; Ciarlet, M.; Rahman, M.; Attoui, H.; Bányai, K.; Estes, M.K.; Gentsch, J.R.; Iturriza-Gómara, M.; Kirkwood, C.D.; Martella, V.; et al. Recommendations for the classification of group A rotaviruses using all 11 genomic RNA segments. Arch. Virol. 2008, 153, 1621–1629. [Google Scholar] [CrossRef]
- Suzuki, T.; Hasebe, A. A provisional complete genome-based genotyping system for rotavirus species C from terrestrial mammals. J. Gen. Virol. 2017, 98, 2647–2662. [Google Scholar] [CrossRef]
- Carvajal, A.; Argüello, H.; Martínez-Lobo, F.J.; Costillas, S.; Miranda, R.; de Nova, P.J.G.; Rubio, P. Porcine epidemic diarrhoea: New insights into an old disease. Porc. Health Manag. 2015, 1, 12. [Google Scholar] [CrossRef]
- Vlasova, A.N.; Wang, Q.; Jung, K.; Langel, S.N.; Malik, Y.S.; Saif, L.J. Porcine Coronaviruses. Emerg. Transbound. Anim. Viruses 2020, 79–110. [Google Scholar] [CrossRef]
- Fairbrother, J.M.; Nadeau, E. Colibacillosis. In Diseases of Swine, 11th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; pp. 807–834. [Google Scholar]
- Uzal, F.A.; Songer, J.G. Clostridial diseases. In Diseases of Swine, 11th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; pp. 792–806. [Google Scholar]
- Vidal, A.; Clilverd, H.; Cortey, M.; Martín-Valls, G.E.; Franzo, G.; Darwich, L.; Martín, M.; Mateu, E. Full-genome characterization by deep sequencing of rotavirus A isolates from outbreaks of neonatal diarrhoea in pigs in Spain. Vet. Microbiol. 2018, 227, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Halaihel, N.; Masía, R.M.; Fernández-Jiménez, M.; Ribes, J.M.; Montava, R.; De Blas, I.; Gironés, O.; Alonso, J.L.; Buesa, J. Enteric calicivirus and rotavirus infections in domestic pigs. Epidemiol. Infect. 2010, 138, 542–548. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Shimoike, T.; Takagi, H.; Murakami, K.; Todaka-Takai, R.; Park, Y.; Katayama, K. Amplification of all 11 RNA segments of group A rotaviruses based on reverse transcription polymerase chain reaction. Microbiol. Immunol. 2012, 56, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Isegawa, Y.; Nakagomi, O.; Nakagomi, T.; Ishida, S.; Uesugi, S.; Ueda, S. Determination of bovine rotavirus G and P serotypes by polymerase chain reaction. Mol. Cell. Probes 1993, 7, 277–284. [Google Scholar] [CrossRef]
- Gentsch, J.R.; Glass, R.I.; Woods, P.; Gouvea, V.; Gorziglia, M.; Flores, J.; Das, B.K.; Bhan, M.K. Identification of group a rotavirus gene 4 types by polymerase chain reaction. J. Clin. Microbiol. 1992, 30, 1365–1373. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed]
- McDonald, S.M.; Matthijnssens, J.; McAllen, J.K.; Hine, E.; Overton, L.; Wang, S.; Lemey, P.; Zeller, M.; Van Ranst, M.; Spiro, D.J.; et al. Evolutionary dynamics of human rotaviruses: Balancing reassortment with preferred genome constellations. PLoS Pathog. 2009, 5, e1000634. [Google Scholar] [CrossRef]
- Farzan, A.; Kircanki, J.; DeLay, J.; Soltes, G.; Songer, J.G.; Friendship, R.; Prescott, J.F. An investigation into the association between cpb2-encoding C. perfringens type A and diarrhea in neonatal piglets. Can. J. Vet. Res. 2013, 77, 45–53. [Google Scholar] [PubMed]
- Shi, Y.; Li, B.; Tao, J.; Cheng, J.; Liu, H. The Complex Co-infections of Multiple Porcine Diarrhea Viruses in Local Area Based on the Luminex xTAG Multiplex Detection Method. Front. Vet. Sci. 2021, 8, 602866. [Google Scholar] [CrossRef]
- Kongsted, H.; Jonach, B.; Haugegaard, S.; Angen, O.; Jorsal, S.E.; Kokotovic, B.; Larsen, L.E.; Jensen, T.K.; Nielsen, J.P. Microbiological, pathological and histological findings in four Danish pig herds affected by a new neonatal diarrhoea syndrome. BMC Vet. Res. 2013, 9, 206. [Google Scholar] [CrossRef]
- Larsson, J.; Aspán, A.; Lindberg, R.; Grandon, R.; Båverud, V.; Fall, N.; Jacobson, M. Pathological and bacteriological characterization of neonatal porcine diarrhoea of uncertain aetiology. J. Med. Microbiol. 2015, 64, 916–926. [Google Scholar] [CrossRef] [PubMed]
- Chan, G.; Farzan, A.; DeLay, J.; McEwen, B.; Prescott, J.F.; Friendship, R.M. A retrospective study on the etiological diagnoses of diarrhea in neonatal piglets in Ontario, Canada, between 2001 and 2010. Can. J. Vet. Res. 2013, 77, 254–260. [Google Scholar] [PubMed]
- Kylla, H.; Dutta, T.K.; Roychoudhury, P.; Mandakini, R.; Subudhi, P.K. Detection and characterization of genogroup 5 Rotavirus associated with piglet diarrhoea in the North East Region of India. Vet. Arhiv 2018, 88, 453–466. [Google Scholar] [CrossRef]
- Amimo, J.O.; Vlasova, A.N.; Saif, L.J. Detection and genetic diversity of porcine group A rotaviruses in historic (2004) and recent (2011 and 2012) swine fecal samples in Ohio: Predominance of the G9P[13] genotype in nursing piglets. J. Clin. Microbiol. 2013, 51, 1142–1151. [Google Scholar] [CrossRef]
- Lachapelle, V.; Sohal, J.S.; Lambert, M.C.; Brassard, J.; Fravalo, P.; Letellier, A.; L’Homme, Y. Genetic diversity of group A rotavirus in swine in Canada. Arch. Virol. 2014, 159, 1771–1779. [Google Scholar] [CrossRef]
- Arruda, P.H.; Madson, D.M.; Ramirez, A.; Rowe, E.; Lizer, J.T.; Songer, J.G. Effect of age, dose and antibiotic therapy on the development of Clostridium difficile infection in neonatal piglets. Anaerobe 2013, 22, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Andrés-Lasheras, S.; Bolea, R.; Mainar-Jaime, R.C.; Kuijper, E.; Sevilla, E.; Martín-Burriel, I.; Chirino-Trejo, M. Presence of Clostridium difficile in pig faecal samples and wild animal species associated with pig farms. J. Appl. Microbiol. 2017, 122, 462–472. [Google Scholar] [CrossRef]
- Zhou, W.; Ullman, K.; Chowdry, V.; Reining, M.; Benyeda, Z.; Baule, C.; Juremalm, M.; Wallgren, P.; Schwarz, L.; Zhou, E.; et al. Molecular investigations on the prevalence and viral load of enteric viruses in pigs from five European countries. Vet. Microbiol. 2016, 182, 75–81. [Google Scholar] [CrossRef]
- Marthaler, D.; Rossow, K.; Culhane, M.; Collins, J.; Goyal, S.; Ciarlet, M.; Matthijnssens, J. Identification, phylogenetic analysis and classification of porcine group C rotavirus VP7 sequences from the United States and Canada. Virology 2013, 446, 189–198. [Google Scholar] [CrossRef]
- Chepngeno, J.; Diaz, A.; Paim, F.C.; Saif, L.J.; Vlasova, A.N. Rotavirus C: Prevalence in suckling piglets and development of virus-like particles to assess the influence of maternal immunity on the disease development. Vet. Res. 2019, 50, 84. [Google Scholar] [CrossRef]
- Martella, V.; Bányai, K.; Lorusso, E.; Bellacicco, A.L.; Decaro, N.; Camero, M.; Bozzo, G.; Moschidou, P.; Arista, S.; Pezzotti, G.; et al. Prevalence of group C rotaviruses in weaning and post-weaning pigs with enteritis. Vet. Microbiol. 2007, 123, 26–33. [Google Scholar] [CrossRef]
- Stadler, J.; Zoels, S.; Fux, R.; Hanke, D.; Pohlmann, A.; Blome, S.; Weissenböck, H.; Weissenbacher-Lang, C.; Ritzmann, M.; Ladinig, A. Emergence of porcine epidemic diarrhea virus in southern Germany. BMC Vet. Res. 2015, 11, 142. [Google Scholar] [CrossRef] [PubMed]
- Grasland, B.; Bigault, L.; Bernard, C.; Quenault, H.; Toulouse, O.; Fablet, C.; Rose, N.; Touzain, F.; Blanchard, Y. Complete genome sequence of a porcine epidemic diarrhea S gene Indel strain isolated in France in December 2014. Genome Announc. 2015, 3, e00535-15. [Google Scholar] [CrossRef] [PubMed]
- Theuns, S.; Conceição-Neto, N.; Christiaens, I.; Zeller, M.; Desmarets, L.M.; Roukaerts, I.D.; Acar, D.D.; Heylen, E.; Matthijnssens, J.; Nauwynck, H.J. Complete genome sequence of a porcine epidemic diarrhea virus from a novel outbreak in Belgium, January 2015. Genome Announc. 2015, 3, e00506-15. [Google Scholar] [CrossRef]
- Dastjerdi, A.; Carr, J.; Ellis, R.J.; Steinbach, F.; Williamson, S. Porcine Epidemic Diarrhea Virus among Farmed Pigs, Ukraine. Emerg. Infect. Dis. 2015, 21, 2235–2237. [Google Scholar] [CrossRef] [PubMed]
- Steinrigl, A.; Fernández, S.R.; Stoiber, F.; Pikalo, J.; Sattler, T.; Schmoll, F. First detection, clinical presentation and phylogenetic characterization of porcine epidemic diarrhea virus in Austria. BMC Vet. Res. 2015, 11, 310. [Google Scholar] [CrossRef] [PubMed]
- Mesquita, J.R.; Hakze-van der Honing, R.; Almeida, A.; Lourenço, M.; van der Poel, W.H.M.; Nascimento, M.S.J. Outbreak of porcine epidemic diarrhea virus in Portugal, 2015. Transbound. Emerg. Dis. 2015, 62, 586–588. [Google Scholar] [CrossRef] [PubMed]
- Dortmans, J.C.F.M.; Li, W.; van der Wolf, P.J.; Buter, G.J.; Franssen, P.J.M.; van Schaik, G.; Houben, M.; Bosch, B.J. Porcine epidemic diarrhea virus (PEDV) introduction into a naive Dutch pig population in 2014. Vet. Microbiol. 2018, 221, 13–18. [Google Scholar] [CrossRef]
- Cortey, M.; Díaz, I.; Vidal, A.; Martín-Valls, G.; Franzo, G.; Gómez de Nova, P.J.; Darwich, L.; Puente, H.; Carvajal, A.; Martín, M.; et al. High levels of unreported intraspecific diversity among RNA viruses in faeces of neonatal piglets with diarrhoea. BMC Vet. Res. 2019, 15, 441. [Google Scholar] [CrossRef]
- Puente, H.; Argüello, H.; Mencía-Ares, Ó.; Gómez-García, M.; Rubio, P.; Carvajal, A. Detection and genetic diversity of porcine coronavirus involved in diarrhea outbreaks in Spain. Front. Vet. Sci. 2021, 8, 651999. [Google Scholar] [CrossRef] [PubMed]
- Wenske, O.; Rückner, A.; Piehler, D.; Schwarz, B.A.; Vahlenkamp, T.W. Epidemiological analysis of porcine rotavirus A genotypes in Germany. Vet. Microbiol. 2018, 214, 93–98. [Google Scholar] [CrossRef]
- Collins, P.J.; Martella, V.; Sleator, R.D.; Fanning, S.; O’Shea, H. Detection and characterisation of group A rotavirus in asymptomatic piglets in southern Ireland. Arch. Virol. 2010, 155, 1247–1259. [Google Scholar] [CrossRef]
- Chandler-Bostock, R.; Hancox, L.R.; Nawaz, S.; Watts, O.; Iturriza-Gomara, M.; Mellits, K.H. Genetic diversity of porcine group A rotavirus strains in the UK. Vet. Microbiol. 2014, 173, 27–37. [Google Scholar] [CrossRef]
- Theuns, S.; Desmarets, L.M.; Heylen, E.; Zeller, M.; Dedeurwaerder, A.; Roukaerts, I.D.; Van Ranst, M.; Matthijnssens, J.; Nauwynck, H.J. Porcine group A rotaviruses with heterogeneous VP7 and VP4 genotype combinations can be found together with enteric bacteria on Belgian swine farms. Vet Microbiol. 2014, 172, 23–34. [Google Scholar] [CrossRef]
- Winiarczyk, S.; Paul, P.S.; Mummidi, S.; Panek, R.; Gradzki, Z. Survey of porcine rotavirus G and P genotype in Poland and the United States using RT-PCR. J. Vet. Med. B 2002, 49, 373–378. [Google Scholar] [CrossRef]
- Steyer, A.; Poljšak-Prijatelj, M.; Barlič-Maganja, D.; Marin, J. Human, porcine and bovine rotaviruses in Slovenia: Evidence of interspecies transmission and genome reassortment. J. Gen. Virol. 2008, 89, 1690–1698. [Google Scholar] [CrossRef]
- Monini, M.; Zaccaria, G.; Ianiro, G.; Lavazza, A.; Vaccari, G.; Ruggeri, F.M. Full-length genomic analysis of porcine rotavirus strains isolated from pigs with diarrhea in Northern Italy. Infect. Genet. Evol. 2014, 25, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Rocha, T.G.; Silva, F.D.; Gregori, F.; Alfieri, A.A.; Buzinaro, M.D.; Fagliari, J.J. Longitudinal study of bovine rotavirus group A in newborn calves from vaccinated and unvaccinated dairy herds. Trop. Anim. Health Prod. 2017, 49, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Steele, A.D.; Victor, J.C.; Carey, M.E.; Tate, J.E.; Atherly, D.E.; Pecenka, C.; Diaz, Z.; Parashar, U.D.; Kirkwood, C.D. Experiences with rotavirus vaccines: Can we improve rotavirus vaccine impact in developing countries? Hum. Vaccines Immunother. 2019, 15, 1215–1227. [Google Scholar] [CrossRef] [PubMed]
- Moise, L.; Gutiérrez, A.H.; Khan, S.; Tan, S.; Ardito, M.; Martin, W.D.; De Groot, A.S. New Immunoinformatics Tools for Swine: Designing Epitope-Driven Vaccines, Predicting Vaccine Efficacy, and Making Vaccines on Demand. Front. Immunol. 2020, 11, 563362. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Enteropathogen | No. Piglets (n = 866) | No. Farms (n = 426) |
|---|---|---|
| C. perfringens | 99 (11.4) | 69 (16.2) |
| Cl. difficile | 13 (1.5) | 9 (2.1) |
| RVA | 25 (2.9) | 22 (5.2) |
| RVC | 3 (0.3) | 2 (0.5) |
| PEDV | 2 (0.2) | 1 (0.2) |
| C. perfringens + Cl. difficile | 121 (13.9) | 93 (21.8) |
| C. perfringens + RVA | 82 (9.5) | 72 (16.9) |
| C. perfringens + RVC | 31 (3.6) | 27 (6.3) |
| C. perfringens + PEDV | 7 (0.8) | 6 (1.4) |
| C. perfringens + Cl. difficile + RVA | 172 (19.9) | 129 (30.3) |
| C. perfringens + Cl. difficile + RVC | 160 (18.5) | 121 (28.4) |
| C. perfringens + Cl. difficile + PEDV | 7 (0.8) | 7 (1.6) |
| C. perfringens + RVA + RVC | 14 (1.6) | 13 (3) |
| C. perfringens + RVA + PEDV | 6 (0.7) | 5 (1.2) |
| C. perfringens + RVC + PEDV | 1 (0.1) | 1 (0.2) |
| C. perfringens + Cl. difficile + RVA + RVC | 65 (7.5) | 54 (12.7) |
| C. perfringens + Cl. difficile + RVA + PEDV | 4 (0.5) | 4 (0.9) |
| C. perfringens + Cl. difficile + RVC + PEDV | 3 (0.3) | 2 (0.5) |
| C. perfringens + RVA + RVC + PEDV | 2 (0.2) | 2 (0.5) |
| Cl. difficile + RVA | 6 (0.7) | 3 (0.7) |
| Cl. difficile + RVC | 3 (0.3) | 3 (0.7) |
| Cl. difficile + RVA + RVC | 4 (0.5) | 3 (0.7) |
| RVA + RVC | 3 (0.3) | 3 (0.7) |
| RVA + PEDV | 3 (0.3) | 3 (0.7) |
| RVA + RVC + PEDV | 3 (0.3) | 3 (0.7) |
| No detection | 27 (3.1) | 18 (4.2) |
| P—G Type | No. of Samples |
|---|---|
| G4P [7] | 14 |
| G9P [23] | 13 |
| G9P [7] | 8 |
| G4P [6] | 7 |
| G3P [7] | 4 |
| G3P [23] | 5 |
| G4P [23] | 3 |
| G3P [13] | 1 |
| G9P [13] | 2 |
| G5P [7] | 2 |
| G3P [6] | 1 |
| G4P [13] | 1 |
| G5P [13] | 1 |
| G11P [13] | 1 |
| G11P [23] | 1 |
| G3P [X] | 1 |
| G4P [X] | 1 |
| G5P [X] | 1 |
| XP [6] | 2 |
| XP [23] | 1 |
| Total | 70 |
| Genotype | No. Maximum of SNPs/nt/Sample | Index |
|---|---|---|
| G4 | 343/981/26 | 0.0134 |
| G9 | 274/981/23 | 0.0121 |
| G3 | 157/981/11 | 0.0145 |
| G5 | 202/981/4 | 0.0515 |
| G11 | 40/979/2 | 0.0204 |
| P [7] | 270/838/28 | 0.0115 |
| P [23] | 290/838/23 | 0.0150 |
| P [6] | 249/838/10 | 0.0297 |
| P [13] | 293/838/6 | 0.0583 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteagudo, L.V.; Benito, A.A.; Lázaro-Gaspar, S.; Arnal, J.L.; Martin-Jurado, D.; Menjon, R.; Quílez, J. Occurrence of Rotavirus A Genotypes and Other Enteric Pathogens in Diarrheic Suckling Piglets from Spanish Swine Farms. Animals 2022, 12, 251. https://doi.org/10.3390/ani12030251
Monteagudo LV, Benito AA, Lázaro-Gaspar S, Arnal JL, Martin-Jurado D, Menjon R, Quílez J. Occurrence of Rotavirus A Genotypes and Other Enteric Pathogens in Diarrheic Suckling Piglets from Spanish Swine Farms. Animals. 2022; 12(3):251. https://doi.org/10.3390/ani12030251
Chicago/Turabian StyleMonteagudo, Luis V., Alfredo A. Benito, Sofía Lázaro-Gaspar, José L. Arnal, Desirée Martin-Jurado, Rut Menjon, and Joaquín Quílez. 2022. "Occurrence of Rotavirus A Genotypes and Other Enteric Pathogens in Diarrheic Suckling Piglets from Spanish Swine Farms" Animals 12, no. 3: 251. https://doi.org/10.3390/ani12030251