
Length at Maturity, Sex Ratio, and Proportions of Maturity of the Giant Electric Ray, Narcine entemedor, in Its Septentrional Distribution

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Samples
2.2. Criteria for Evaluating Maturity
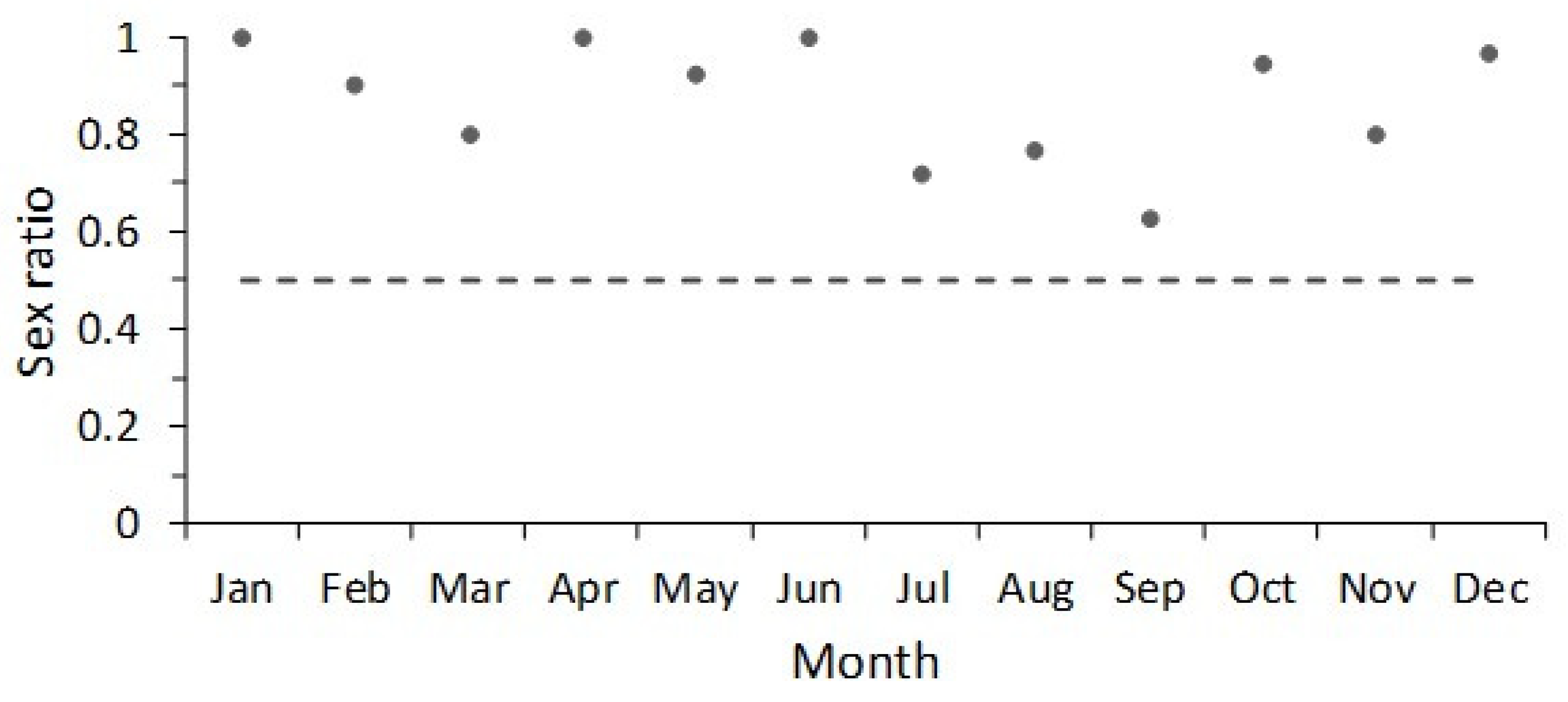
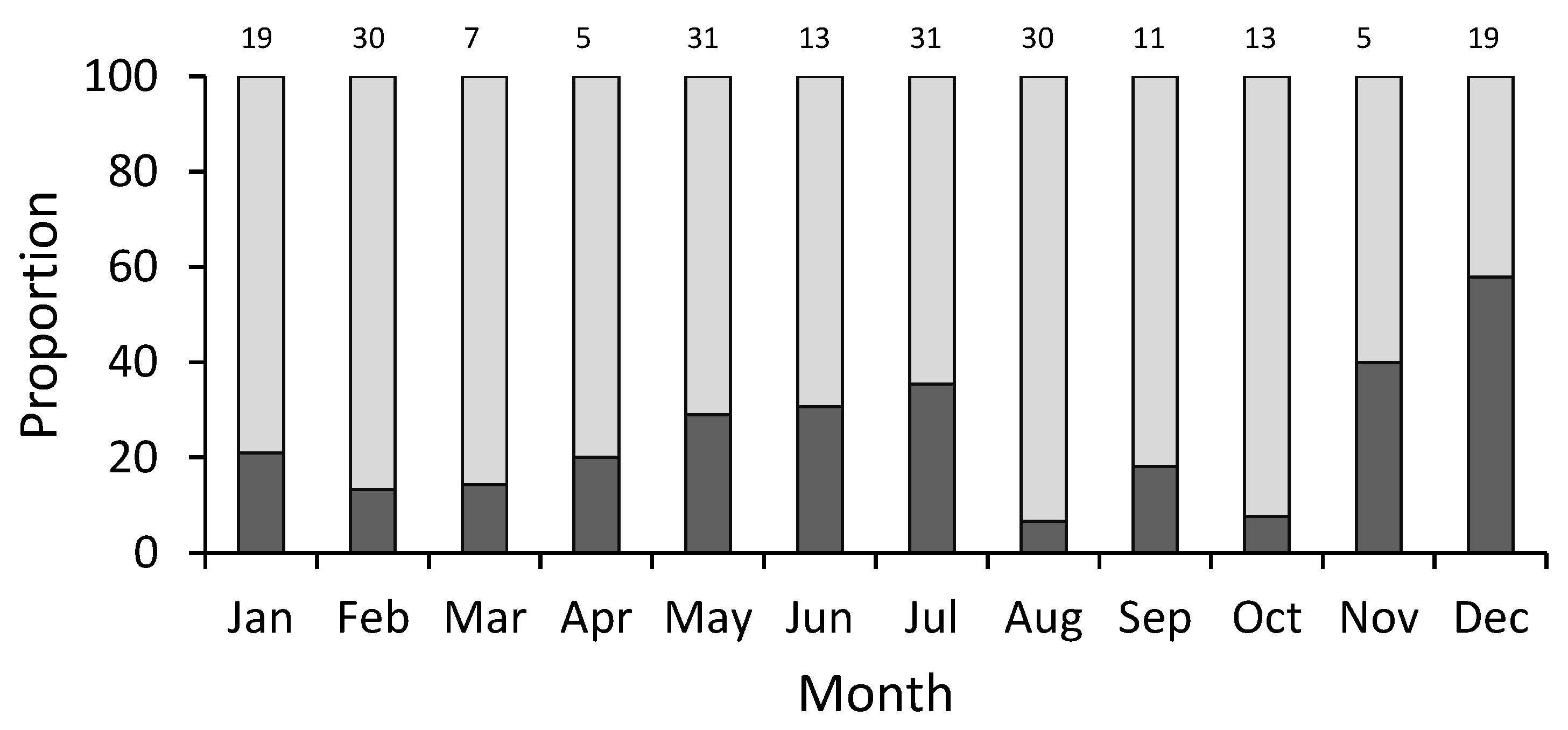
2.3. Sex Ratio
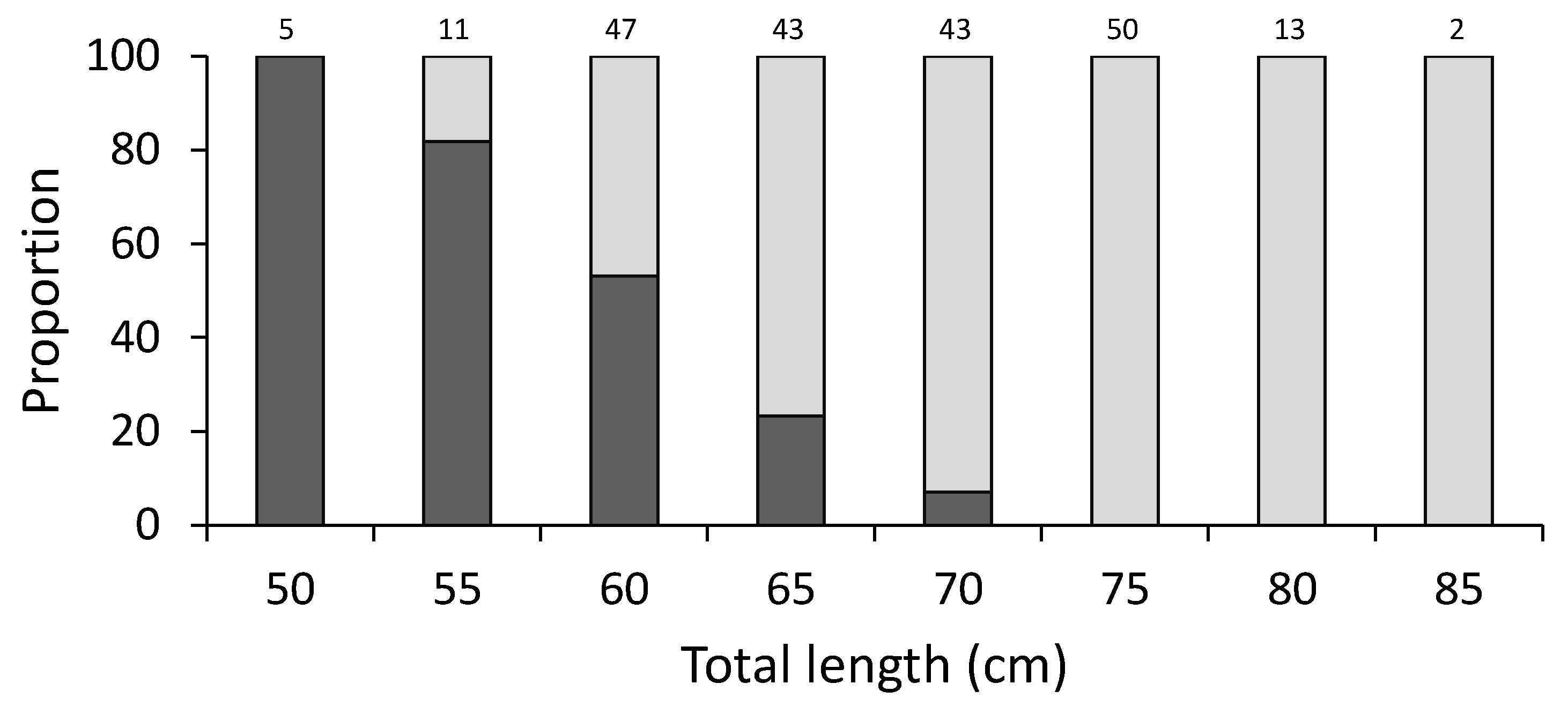
2.4. Length at Maturity
3. Results
4. Discussion
4.1. Length at Maturity
4.2. Features of the Reproductive Biology Affecting
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, Y.; Paloheimo, J.E. Estimating fish length and age at 50% maturity using a logistic type model. Aquat. Sci. 1994, 56, 206–219. [Google Scholar] [CrossRef]
- Cerdenares-Ladrón de Guevara, G.; Morales-Bojórquez, E.; Rodriguez-Sánchez, R. Age and growth of the sailfish Istiophorus platypterus (Istiophoridae) in the Gulf of Tehuantepec, Mexico. Mar. Biol. Res. 2011, 7, 488–499. [Google Scholar] [CrossRef]
- Jacob-Cervantes, M.L.; Aguirre-Villaseñor, H. Inferencia multimodelo y selección de modelos aplicados a la determinación de L50 para la sardina crinuda Opisthonema libertate del sur del golfo de California. Cienc. Pesq. 2014, 22, 61–68. [Google Scholar]
- Caddy, J.F.; Mahon, R. Reference Points for Fisheries Management; FAO Fisheries Technical Paper. No. 347; FAO: Rome, Italy, 1995. [Google Scholar]
- Midway, S.R.; Scharf, F.S. Histological analysis reveals larger size at maturity for southern flounder with implications for biological reference points. Mar. Coast. Fish. Dyn. Manag. Ecosyst. Sci. 2012, 4, 628–638. [Google Scholar] [CrossRef]
- Restrepo, V.R.; Watson, R.A. An approach to modeling crustacean egg-bearing fractions as a function of size and season. Can. J. Fish. Aquat. Sci. 1991, 48, 1431–1436. [Google Scholar] [CrossRef]
- Roa, R.; Ernst, B.; Tapia, F. Estimation of size at sexual maturity: An evaluation of analytical and resampling procedures. Fish. Bull. 1999, 97, 570–580. [Google Scholar]
- Gompertz, B. On the nature of the function expressive of the law of human mortality, and on a new mode of determining the value of life contingencies. Philos. Trans. R. Soc. Lond. 1825, 115, 513–583. [Google Scholar] [CrossRef]
- Lysack, W. 1979 Lake Winnipeg Fish Stock Assessment Program; Manitoba Department of Natural Resources; Fisheries Branch: Winnipeg, MB, Canada, 1980.
- White, T.; Hall, G.; Potter, C. Size and age compositions and reproductive biology of the nervous shark Carcharhinus cautus in a large subtropical embayment, including an analysis of growth during pre- and postnatal life. Mar. Biol. 2002, 1153–1164. [Google Scholar] [CrossRef]
- Flores, L.; Ernst, B.; Parma, A.M. Growth pattern of the sea urchin, Loxechinus albus (Molina, 1782) in Southern Chile: Evaluation of growth models. Mar. Biol. 2010, 157, 967–977. [Google Scholar] [CrossRef]
- Oviedo-Pérez, J.L.; Zea-De la Cruz, H.; Aguirre-Villaseñor, H.; Meiners-Mandujano, C.; Jiménez-Badillo, L.; González-Ocarranza, L. Talla de madurez sexual del tiburón Rhizoprionodon terraenovae en Veracruz, México. Cienc. Pesq. 2014, 22, 37–45. [Google Scholar]
- Katsanevakis, S.; Maravelias, C.D. Modelling fish growth: Multi-model inference as a better alternative to a priori using von Bertalanffy equation. Fish Fish. 2008, 9, 178–187. [Google Scholar] [CrossRef]
- Morales-Bojórquez, E.; Aguirre-Villaseñor, H.; Cerdenares-Ladrón De Guevara, G. Confrontación de hipótesis múltiples en pesquerías, teoría de la información y selección de modelos. Cienc. Pesq. 2014, 22, 9–10. [Google Scholar]
- Zhu, G.P.; Dai, X.J.; Song, L.M.; Xu, L.X. Size at sexual maturity of bigeye tuna Thunnus obesus (Perciformes: Scombridae) in the tropical waters: A comparative analysis. Turk. J. Fish. Aquat. Sci. 2011, 11, 149–156. [Google Scholar] [CrossRef]
- García-Rodríguez, A.; Hernández-Herrera, A.; Galván-Magaña, F.; Ceballos-Vázquez, B.P.; Pelamatti, T.; Tovar-Ávila, J. Estimation of the size at sexual maturity of the bat ray (Myliobatis californica) in Northwestern Mexico through a multi-model inference. Fish. Res. 2020, 231, 105712. [Google Scholar] [CrossRef]
- Hamlett, W.C. Reproductive Biology and Phylogeny of Chondrichthyes: Sharks, Batoids and Chimaeras; Hamlett, W.C., Ed.; Reproductive Biology and Phylogeny; Science Publishers, Inc.: Enfield, NH, USA, 2005; ISBN 9781578082711. [Google Scholar]
- Burgos-Vázquez, M.I.; Mejía-Falla, P.A.; Cruz-Escalona, V.H.; Brown-Peterson, N.J. Reproductive strategy of the giant electric ray in the Southern Gulf of California. Mar. Coast. Fish. 2017, 9, 577–596. [Google Scholar] [CrossRef] [Green Version]
- Villavicencio-Garayzar, C.J. Taxonomía, Abundancia Estacional, Edad y Crecimiento y Biología Reproductiva de Narcine entemedor Jordan y Starks (Chondrichthyes; Narcinidae), en Bahía Almejas, B.C.S, México. Ph.D. Thesis, Universidad Autónoma de Nuevo Leon, San Nicolás de los Garza, México, 2000. [Google Scholar]
- Robertson, D.R.; Allen, G. Peces Costeros del Pacífico Oriental Tropical: Un Sistema de Información en Línea. Available online: https://biogeodb.stri.si.edu/sftep/es/pages (accessed on 8 November 2021).
- Clarke, T.M.; Espinoza, M.; Ahrens, R.; Wehrtmann, I.S. Elasmobranch bycatch associated with the shrimp trawl fishery off the Pacific Coast of Costa Rica, Central America. Fish. Bull. 2016, 114, 1–17. [Google Scholar] [CrossRef]
- Márquez-Farías, J.F. The artisanal ray fishery in the Gulf of California: Development, fisheries research and management issues. IUCN/SSC Shark Spec. Group 2002, 14, 12–13. [Google Scholar]
- Ramírez-Amaro, S.R.; Cartamil, D.; Galvan-Magaña, F.; Gonzalez-Barba, G.; Graham, J.B.; Carrera-Fernandez, M.; Escobar-Sanchez, O.; Sosa-Nishizaki, O.; Rochin-Alamillo, A. The artisanal elasmobranch fishery of the Pacific Coast of Baja California, Mexico. Sci. Mar. 2013, 77, 473–487. [Google Scholar] [CrossRef]
- Zar, J. Biostatistical Analysis; Pearson Prentice-Hall: Hoboken, NJ, USA, 2010; ISBN 9788578110796. [Google Scholar]
- Cerdenares-Ladrón de Guevara, G.; Morales-Bojórquez, E.; Rodríguez-Jaramillo, C.; Hernández-Herrera, A.; Abitia-Cárdenas, A. Seasonal reproduction of sailfish Istiophorus platypterus from the Southeast Mexican Pacific. Mar. Biol. Res. 2013, 9, 407–420. [Google Scholar] [CrossRef]
- Brouwer, S.L.; Griffiths, M.H. Reproductive biology of carpenter seabream (Argyrozona argyrozona) (Pisces: Sparidae) in a marine protected area. Fish. Bull. 2005, 103, 258–269. [Google Scholar]
- Luquin-Covarrubias, M.A.; Morales-Bojórquez, E.; González-Peláez, S.S.; Hidalgo-De-La-Toba, J.Á.; Lluch-Cota, D.B. Modeling of growth depensation of geoduck clam Panopea globosa based on a multimodel inference approach. J. Shellfish. Res. 2016, 35, 379–387. [Google Scholar] [CrossRef]
- Haddon, M. Modelling and Quantitative Methods in Fisheries; Statistics/Biology; Chapman and Hall: Boca Raton, FL, USA, 2001; ISBN 9781584881773. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; ISBN 0387953647. [Google Scholar]
- Okamura, H.; Punt, A.E.; Semba, Y.; Ichinokawa, M. Marginal increment analysis: A new statistical approach of testing for temporal periodicity in fish age verification. J. Fish Biol. 2013, 82, 1239–1249. [Google Scholar] [CrossRef] [PubMed]
- Remya Mohan, S.; Harikrishnan, M.; Sherly Williams, E. Reproductive biology of a gobiid fish Oxyurichthys tentacularis (Valenciennes, 1837) inhabiting ashtamudi lake, S. India. J. Appl. Ichthyol. 2018, 34, 1099–1107. [Google Scholar] [CrossRef]
- Chelapurath Radhakrishnan, R.; Kuttanelloor, R.; Balakrishna, M.K. Reproductive biology of the endemic cyprinid fish Hypselobarbus thomassi (Day, 1874) from Kallada River in the Western Ghats, India. J. Appl. Ichthyol. 2020, 36, 604–612. [Google Scholar] [CrossRef]
- Chang, H.-Y.; Sun, C.-L.; Yeh, S.-Z.; Chang, Y.-J.; Su, N.-J.; DiNardo, G. Reproductive biology of female striped marlin Kajikia audax in the Western Pacific Ocean. J. Fish Biol. 2018, 92, 105–130. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.D.; Cailliet, G.M.; Melendez, E.M. Maturity and growth characteristics of a commercially exploited stingray, Dasyatis dipterura. Mar. Freshw. Res. 2007, 58, 54–66. [Google Scholar] [CrossRef]
- Jacobsen, I.P.; Bennett, M.B. Life history of the blackspotted whipray Himantura astra. J. Fish Biol. 2011, 78, 1249–1268. [Google Scholar] [CrossRef] [PubMed]
- Kume, G.; Furumitsu, K.; Yamaguchi, A. Age, growth and age at sexual maturity of fan ray Platyrhina sinensis (Batoidea: Platyrhinidae) in Ariake Bay, Japan. Fish. Sci. 2008, 74, 736–742. [Google Scholar] [CrossRef]
- Snelson, F.F., Jr.; Williams-Hooper, S.E.; Schmid, T.H. Biology of the bluntnose stingray, Dasyatis sayi, in Florida Coastal Lagoons. Bull. Mar. Sci. 1989, 45, 15–25. [Google Scholar]
- Consalvo, I.; Scacco, U.; Romanelli, M.; Vacchi, M. Comparative study on the reproductive biology of Torpedo torpedo (Linnaeus, 1758) and T. Marmorata (Risso, 1810) in the Central Mediterranean Sea. Sci. Mar. 2007, 71, 213–222. [Google Scholar] [CrossRef]
- Rolim, F.A.; Rotundo, M.M.; Vaske-Júnior, T. Notes on the reproductive biology of the brazilian electric ray Narcine brasiliensis (Elasmobranchii: Narcinidae). J. Fish Biol. 2016, 89, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Koob, T.J.; Callard, I.P. Reproductive endocrinology of female elasmobranchs: Lessons from the Little Skate (Raja erinacea) and Spiny Dogfish (Squalus acanthias). J. Exp. Zool. 1999, 284, 557–574. [Google Scholar] [CrossRef]
- Kadri, H.; Marouani, S.; Saïdi, B.; Bradai, M.N.; Bouaïn, A.; Morize, E. Age, growth, sexual maturity and reproduction of the thornback ray, Raja clavata (L.), of the Gulf of Gabès (South-Central Mediterranean Sea). Mar. Biol. Res. 2014, 10, 416–425. [Google Scholar] [CrossRef]
- Cannizzaro, L.; Garofalo, G.; Levi, D.; Rizzo, P.; Gancitano, S. Raja clavata (Linneo,1758) Nel Canale Di Sicilia: Crescita, distribuzione e abbondanza. Biol. Mar. Mediterr. 1995, 2, 257–262. [Google Scholar]
- Whittamore, J.M.; McCarthy, I.D. The population biology of the thornback ray, Raja clavata in Caernarfon Bay, North Wales. J. Mar. Biol. Assoc. U.K. 2005, 85, 1089–1094. [Google Scholar] [CrossRef]
- Maia, C.; Erzini, K.; Serra-Pereira, B.; Figueiredo, I. Reproductive biology of cuckoo ray Leucoraja naevus. J. Fish Biol. 2012, 81, 1285–1296. [Google Scholar] [CrossRef]
- McCully, S.R.; Scott, F.; Ellis, J.R. Lengths at maturity and conversion factors for skates (Rajidae) around the British Isles, with an analysis of data in the literature. ICES J. Mar. Sci. 2012, 69, 1812–1822. [Google Scholar] [CrossRef] [Green Version]
- Palma-Chávez, J.J. Biología Reproductiva de Las Rayas: Mariposa Gymnura marmorata (Cooper, 1863) y Torpedo Narcine entemedor (Jordan and Starks, 1895) Desembarcados En Santa Rosa-Salinas. Master’s Thesis, Universidad Laica Eloy Alfaro de Manabí, Manta, Ecuador, 2014. [Google Scholar]
- Ellis, J. Raja clavata. The IUCN Red List of Threatened Species 2016: E.T39399A103110667; 2016; Available online: http://dx.doi.org/10.2305/IUCN.UK.2016-3.RLTS.T39399A103110667.en (accessed on 8 November 2021).
- Ellis, J.; Dulvy, N.; Walls, R. Leucoraja naevus. The IUCN Red List of Threatened Species 2015: E.T161626A48949434; 2015; Available online: http://dx.doi.org/10.2305/IUCN.UK.2015-1.RLTS.T161626A48949434.en (accessed on 8 November 2021).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Abbreviation | Function | Source |
|---|---|---|---|
| Gompertz | GOM | [8] | |
| White | WHI | [10] |
| Female | Male | X2 | |
|---|---|---|---|
| January | 21 | 0 | 10.50 |
| February | 37 | 4 | 13.28 |
| March | 8 | 2 | 1.80 |
| April | 8 | 0 | 4.00 |
| May | 35 | 3 | 13.47 |
| June | 17 | 0 | 8.50 |
| July | 33 | 13 | 4.35 |
| August | 33 | 10 | 6.15 |
| September | 15 | 9 | 0.75 |
| October | 16 | 1 | 6.62 |
| November | 8 | 2 | 1.80 |
| December | 29 | 1 | 13.07 |
| Model | L50 | L95 | ε | AIC | ||
|---|---|---|---|---|---|---|
| GOM | 58.50 (56.2–60) | 0.231 (0.16–0.31) | 73.585 | 151.16 | ||
| WHI | 60.64 (58.6–62.2) | 70.37 (67.6–74.4) | 1 | 72.970 | 151.94 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Palafox, X.A.; Morales-Bojórquez, E.; Aguirre-Villaseñor, H.; Cruz-Escalona, V.H. Length at Maturity, Sex Ratio, and Proportions of Maturity of the Giant Electric Ray, Narcine entemedor, in Its Septentrional Distribution. Animals 2022, 12, 120. https://doi.org/10.3390/ani12010120
Pérez-Palafox XA, Morales-Bojórquez E, Aguirre-Villaseñor H, Cruz-Escalona VH. Length at Maturity, Sex Ratio, and Proportions of Maturity of the Giant Electric Ray, Narcine entemedor, in Its Septentrional Distribution. Animals. 2022; 12(1):120. https://doi.org/10.3390/ani12010120
Chicago/Turabian StylePérez-Palafox, Xchel Aurora, Enrique Morales-Bojórquez, Hugo Aguirre-Villaseñor, and Víctor Hugo Cruz-Escalona. 2022. "Length at Maturity, Sex Ratio, and Proportions of Maturity of the Giant Electric Ray, Narcine entemedor, in Its Septentrional Distribution" Animals 12, no. 1: 120. https://doi.org/10.3390/ani12010120