
The Cytogenetics of the Water Buffalo: A Review


1
National Research Council (CNR), Institute of Animal Production System in Mediterranean Environment (ISPAAM), Piazzale E. Fermi, 1, 8055 Portici, Italy
2
Department of Agriculture and Environmental Sciences, University of Milan, 20133 Milan, Italy
*
Author to whom correspondence should be addressed.
Animals 2021, 11(11), 3109; https://doi.org/10.3390/ani11113109
Submission received: 30 September 2021
/
Revised: 25 October 2021
/
Accepted: 26 October 2021
/
Published: 30 October 2021
(This article belongs to the Special Issue The Water Buffalo (Bubalus bubalis))
Abstract
:Simple Summary
Currently, there are two recognized genera of buffalo worldwide: the Syncerus (from the African continent), and the Bubalus (from the southwest Asian continent, Mediterranean area, southern America, and Australia). All species belonging to these genera have specific chromosome numbers and shapes. Because of such features, the study of chromosomes is a fascinating biological basis for differentiating the various species (and hybrids) of buffaloes and characterizing their karyotypes for evolutionary, clinical, and molecular studies. In this review, we report an update of the most important studies in which the evolutionary, clinical, and molecular cytogenetics of buffaloes were described—particularly those belonging to the river buffalo. In addition, we show new data on swamp buffalo chromosomes.
Abstract
The water buffalo (Bubalus bubalis), also known as the Asian buffalo, is an essential domestic bovid. Indeed, although its world population (~209 million heads) is approximately one-ninth that of cattle, the management of this species involves a larger human population than that involved with raising cattle. Compared with cattle, water buffalo have been understudied for many years, but interest in this species has been increasing, especially considering that the world population of these bovids grows every year—particularly that of the river buffalo. There are two genera of buffalo worldwide: the Syncerus (from the African continent), and the Bubalus (from the southwest Asian continent, Mediterranean area, southern America, and Australia). All species belonging to these two genera have specific chromosome numbers and shapes. Because of such features, the study of chromosomes is a fascinating biological basis for differentiating various species (and hybrids) of buffaloes and characterizing their karyotypes in evolutionary, clinical, and molecular studies. In this review, we report an update on essential cytogenetic studies in which various buffalo species were described from evolutionary, clinical, and molecular perspectives—particularly considering the river buffalo (Bubalus bubalis 2n = 50). In addition, we show new data on swamp buffalo chromosomes.
1. Introduction
Chromosomes are an exciting biological material for studying the evolution of species, because most mutations accumulate at the chromosome level. At the same time, abnormal chromosome constitution is responsible for defects in both body conformation and reproduction. The water buffalo (Bubalus bubalis), also known as the Asian buffalo, is a crucial domestic bovid. Indeed, although its world population (~208,098,759 heads) [1] is approximately one-ninth that of cattle, the management to raise this species requires a larger human population than that involved in cattle rearing. The water buffalo is raised for both milk (mainly) and meat production, although there are differences between countries. In particular, milk can be consumed directly for nutrition (as occurs in East Asian countries) or transformed into products for commercial use [1]. The buffalo species has been understudied for many years compared with cattle, but the interest in this species has increased over the past two decades, and its population has been growing by 1.8 million heads/year for river buffalo, while the swamp buffalo population has decreased in the past decade, by an average of 180,000 heads/year [2].
There are two genera of buffalo worldwide: the African Syncerus (Figure 1A), and the Bubalus, also known as the water buffalo (Figure 1B,C). The latter is mainly present in East Asian countries, the Mediterranean area, southern America, Australia, and has several breeds—especially in India, Pakistan, and China [1]. All species belonging to these two genera have specific chromosome numbers and shapes, making the chromosomes useful for differentiating the two genera and various buffalo species (and hybrids). These chromosome changes have been the result of karyotype evolution, determining several species. Furthermore, the study of chromosomes has been mainly used in animal cytogenetics to (1) verify the relationship between chromosome abnormalities and fertility [3,4,5,6,7]; (2) physically map both type I (expressed sequences) and type II (SSRs, microsatellite marker, STSs) loci, especially using fluorescence in situ hybridization (FISH) techniques [8,9,10,11,12]; (3) correctly identify the chromosomes involved in chromosomal abnormalities via chromosome banding techniques [13,14]; (4) reveal chromosome rearrangements occurring in some chromosomal abnormalities, especially using both FISH mapping [15,16,17] and comparative genome hybridization array (aCGH) techniques [18,19]; (5) compare related and unrelated genomes by using the Zoo-FISH technique [20,21,22,23,24,25,26,27,28], centromeric SAT sequences by FISH mapping [29], or detailed FISH mapping along chromosomes [30,31,32,33,34,35,36]; and (6) test the genome stability of several bovids, including the river buffalo, with both in vitro and in vivo (natural) exposure to potential mutagens [37,38,39,40,41,42,43,44,45], or affected by limb malformations [46].
Standard chromosomal nomenclatures have been proposed for domestic bovids, including the river buffalo, where a standard karyotype was published using six different banding techniques [47]. This is the only example of a standard karyotype in any mammalian species, including humans, using six different banding techniques. However, discrepancies in the standard chromosomal nomenclatures of domestic bovids were observed when comparing the G- and R-banding patterns of some chromosomes. The differences in these standard nomenclatures were observed using both chromosome banding and FISH mapping techniques, mainly using marker chromosomes (biarmed pairs) [48,49,50]. A final agreement for a standard chromosomal nomenclature of all of the most critical domestic bovids (including the river buffalo) was found only when the G-Q and R-banding techniques were performed on cattle chromosomes by two different cytogenetic labs also using chromosome-specific markers via FISH mapping [51,52]. The same markers used in cattle chromosomes were also assigned to river buffalo, sheep, and goat chromosomes via FISH mapping [53,54], further supporting the agreement of the results obtained at the ISCNDB2000 [52].
In this review, we report the most critical data on the studies that characterized the evolutionary, clinical, and molecular cytogenetics of buffaloes—particularly those involving the water buffalo. In addition, we also show and discuss new data on swamp buffalo chromosomes.
2. Evolutionary Cytogenetics
The Bovidae are a family belonging to the Cetartiodactyla order, suborder Ruminantia. They comprise 149 different species grouped into 7 subfamilies [55]. The Bovinae comprise several species, including those in the genera Bos, Bubalus, and Syncerus, the latter belonging to the tribe Bovini, in which domesticated and undomesticated species are included [56]. The Robertsonian translocation (rob) has been the most common chromosomal mechanism characterizing the evolutionary history of many autosome chromosomes from bovid species. Still, complex chromosome rearrangements are also followed by sex chromosomes [36,57].
2.1. Autsosomes
Rob occurs when two non-homologous acrocentric chromosomes are joined along the centromeres. This fusion is generally accompanied by loss of constitutive heterochromatin (HC) (C-bands) localized at the centromeric regions, as shown in river buffalo [58] (Figure 2). This occurred frequently during the karyotype evolution of bovid autosomes, including the buffalo species. The number of chromosomes (2n) decreases during this process, but the number of chromosome arms (fundamental number; FN) is conserved. This simple discovery by Wurster and Benirschke [59] was later confirmed by comparative chromosome banding [60,61,62,63,64] and comparative FISH mapping analyses among bovid species [24,30,31,65,66,67,68].
With analyses of both chromosome banding comparisons and FISH mapping data, researchers have concluded that the Bovinae subfamily might be ancestral to the remaining subfamily (for details, see [57]). Rob occurred during the evolution of buffaloes, with their autosomal karyotypes resulting in various species having different diploid numbers, chromosome shapes, and chromosome linkage associations. All known species, subspecies, and hybrids evolved in this way, except for the buffalo Anoa depressicornis, which has the same 2n and FN in the two known species (Table 1).
As shown in Table 1, two genera of buffaloes exist worldwide: Syncerus (African buffalo), and Bubalus (Asian buffalo or water buffalo). The African buffalo has two subspecies: the Syncerus caffer caffer (2n = 52; FN = 60), and the Syncerus caffer nanus (2n = 54; FN = 60), with the presence of four and three biarmed pairs, respectively. By considering the biarmed pairs through the use of chromosome banding comparison with cattle chromosomes (ancestral bovid karyotype) [62] and the standard chromosomal nomenclatures [47,52], the four and three biarmed pairs of S. c. caffer and S. c. nanus, respectively, were established to be homologous to cattle chromosomes 1;13, 2;3, 5;20, and 11;29; and 1;13, 2;3, and 5;20, respectively (Table 1). For this reason, crossbreeds between the two species are possible, although the hybrids (2n = 53) could have reproductive problems because of the trivalent configurations during meiosis, and subsequent unbalanced gametes and embryos.
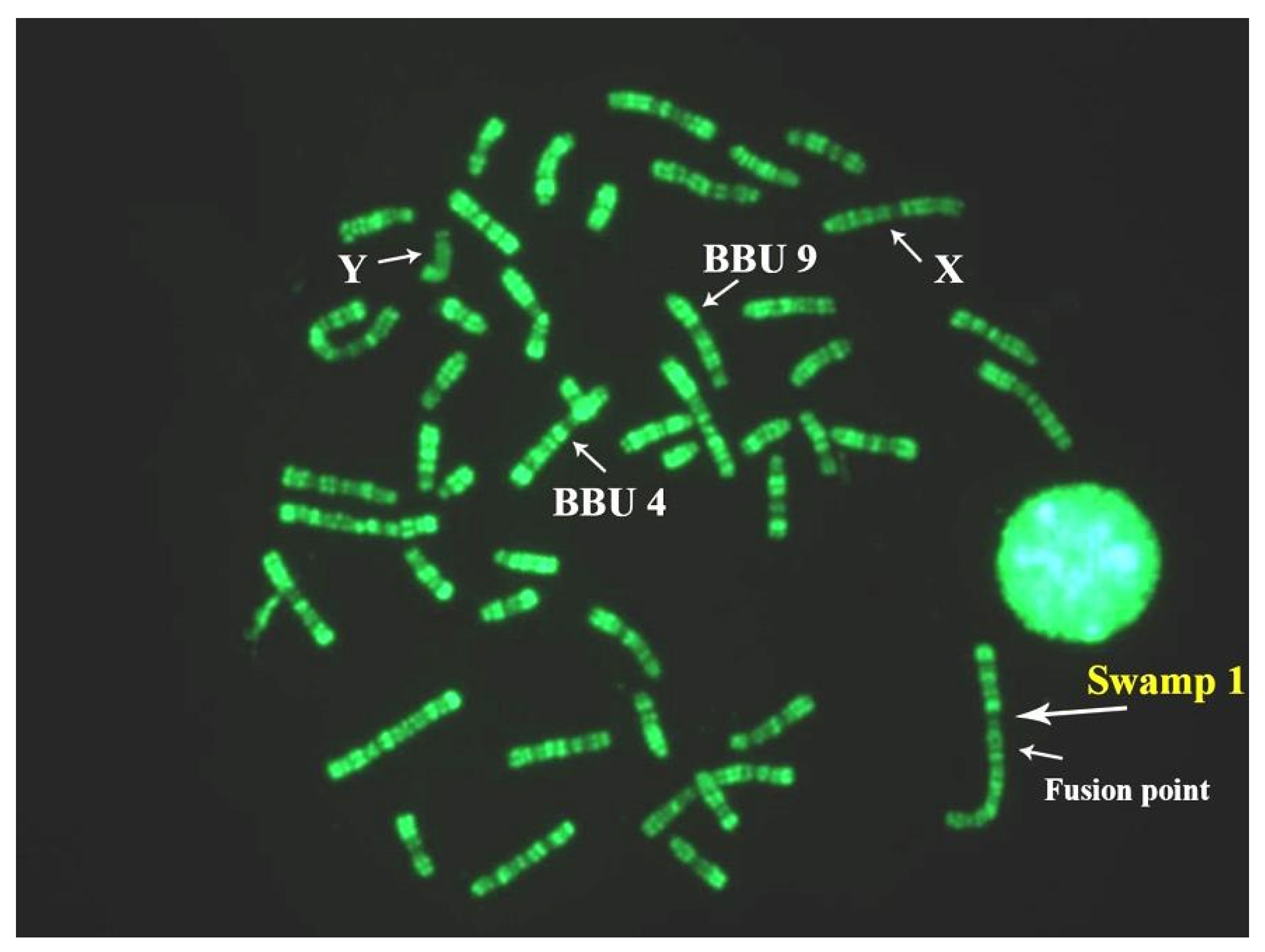
The Asian buffalo (BBU) includes two species: B. bubalis, and B. mindorensis. Both species are probably derived from Bubalus arnee, the ancestral water buffalo, which is currently considered an endangered species [1,2]. B. bubalis includes two subspecies: the river type (B. b. bubalis, 2n = 50, FN = 60), and the swamp type (B. b. carabanensis; 2n = 48, FN = 58) [2,69]—or simply B. bubalis (the river type) and B. kerabau (the swamp type), as recently reported [70]. Moreover, in the Asian buffalo, the diploid number varies between the two subspecies but, differently from the African buffalo—where the two subspecies differentiate from one additional centric fusion—the Asian buffaloes (B. bubalis) show the same number of biarmed pairs, but a different diploid number (Table 1). This phenomenon was explained by high-resolution chromosome banding comparisons between the river and swamp buffaloes [61]. Indeed, it was possible to demonstrate that river (2n = 50) and swamp (2n = 48) buffaloes are differentiated by a tandem fusion translocation that occurs between the telomeres of river buffalo chromosome 4p (BBU4p) and the centromere of river buffalo chromosome 9 (BBU9). The largest swamp buffalo chromosome 1 originated during this process [61]. This fusion was later confirmed in another study [71].
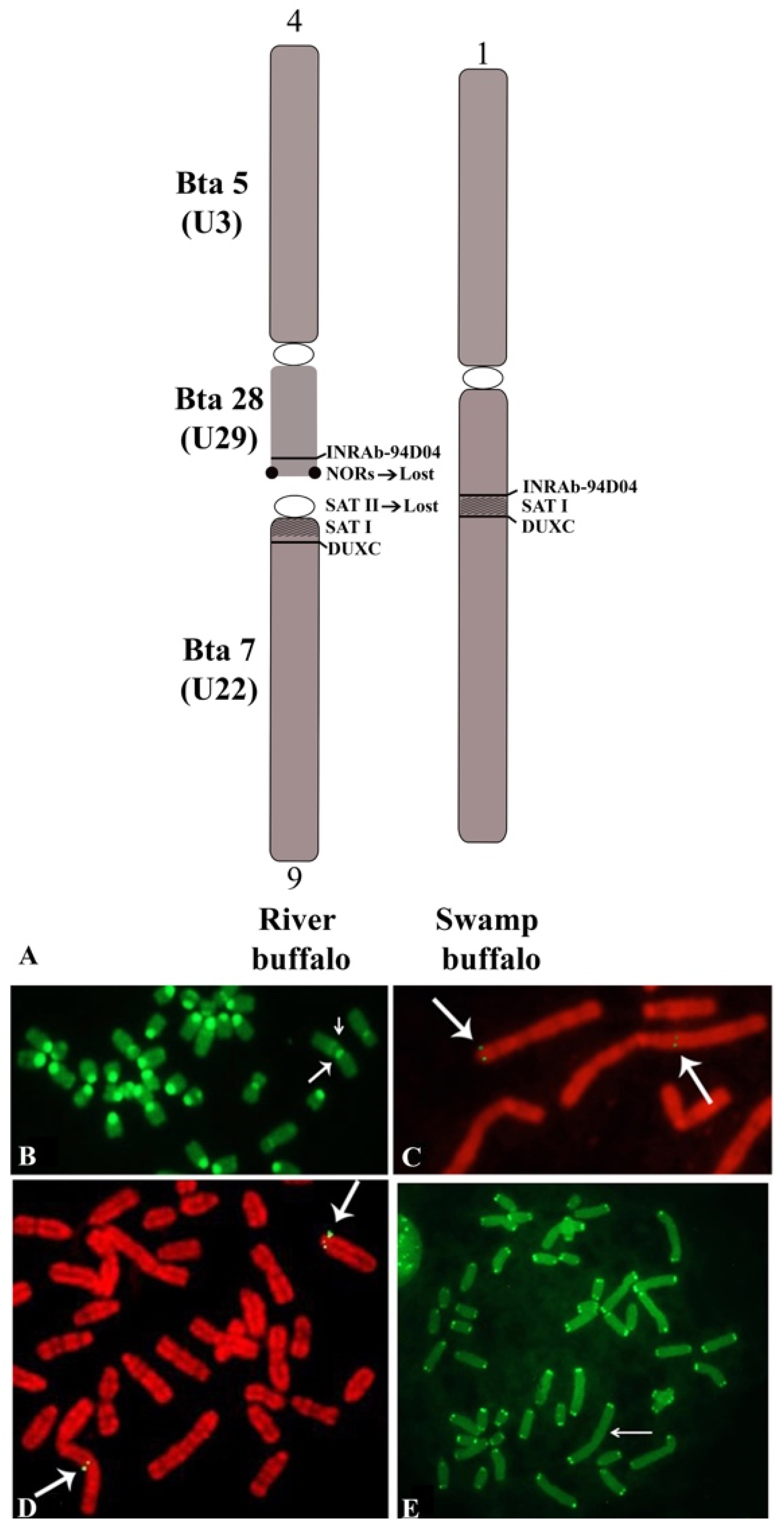
Considering the biarmed pairs between the two subspecies, we can see that all five biarmed pairs of the river buffalo are also present in swamp buffalo. The BBU4 (biarmed) chromosome was fused with the BBU9 (acrocentric) chromosome, resulting in a large swamp buffalo chromosome 1 (Table 1). During this tandem fusion translocation, the nucleolus organizer chromosomes (NORs) available at the telomeres of river buffalo chromosome 4p, along with some centromeric sequences of river buffalo chromosome 9, were lost (Figure 3A) [61,72,73]. The visualization of the centromeric positive C-band in the large swamp buffalo chromosome 1, along with a proximal and pale positive C-band, is allowed by the CBA banding technique (Figure 3B). The proximal and pale positive band observed in swamp buffalo chromosome 1 is likely to be part of the centromere of BBU9 (SAT-I sequences). To ensure that no other chromosome regions were lost during this fusion, we used two probes mapping distally to BBU4p (homologous to BTA28) (INRA_94D04, present in the ARS UCD1.1 genome at 47 Mb of BTA28) and very proximally (pericentromeric) to BBU9 (homologous to BTA7) (DUXC, present in the ARS UCD1.1 genome at 82 Kb of BTA7) [74]. Clear hybridization signals were observed at the distal BBU4p and pericentromeric region of BBU9, as well as the corresponding regions (fusion point) of swamp buffalo chromosome 1 (Figure 3A,C,D). This result is a confirmation that only the NORs present at BBU4p telomeres and some centromeric regions of BBU9 were lost during the tandem fusion. Furthermore, it was shown by a PNA telomeric probe that there were clear signals only at the telomeres of swamp buffalo chromosome 1, but they were absent at the fusion point (Figure 3E). This fusion could have been facilitated by the presence of a positive C-band distally located—almost telomeric—at BBU4p, which is often heteromorphic within the chromosome pair, as previously reported [58].
The two subspecies can interbreed, and their hybrids (2n = 49; Figure 4) are fertile. The hypothesis that the swamp buffalo was derived from the river type is supported by data, as suggested by mitochondrial DNA studies in water buffaloes [75]. However, closer genetic affinities between wild and swamp buffaloes than between wild and river buffaloes [76] have been revealed in studies on wild buffalo (B. arnee). Nevertheless, more information about the real natural karyotype evolution of these species can only be provided by cytogenetic studies of wild buffalo (B. arnee).
Crosses between river and swamp buffaloes are often undertaken to increase milk production in the swamp buffalo population, especially using Indian river buffalo breeds such as Murrah and Jafarabadi. However, the existence of riverine river–swamp hybrids with 49 chromosomes among the Indian Chilika buffalo population has been recorded for the first time. This feature was also confirmed by mitochondrial haplotype sharing between Chilika and Indian/Chinese swamp buffalo populations in a network analysis of swamp buffalo hybrids [77]. Indeed, milk production was noticeably increased by the cross between the two subspecies, although the results obtained using different breeds of both swamp and river buffalo were different (from 20% more to triple milk production just in F1 hybrids) [78,79].
Still, two issues remain crucial considering this matter: (1) the greater increase in milk production obtained from the hybrids must be sustained with adequate food nutrition, and this is often difficult considering that swamp buffaloes are raised in tiny farms with few animals (one or two heads on average) and few available food resources; (2) the hybrids have 49 chromosomes, and for this reason, reproductive problems can arise due to the presence of the trivalent configuration during meiosis, with possible subsequent anomalous meiotic segregations. In this process, abnormal gametes and embryos are produced which, consequently, can die in early embryonic life, as found in cattle carrying robs (1;29) [80,81]. Indeed, a higher frequency of abnormal pairings in the hybrids (both F1 and F2 backcrosses) at meiosis was revealed by synaptonemal complex (SC) analyses of river and swamp buffaloes and their hybrids [82].
Studies on gametes (semen in particular) and embryos using specific chromosome molecular markers and the FISH technique, as performed in a river buffalo bull carrying a chromosome abnormality via sperm FISH [83], could give more precise indications of the crosses between the two water buffalo subspecies.
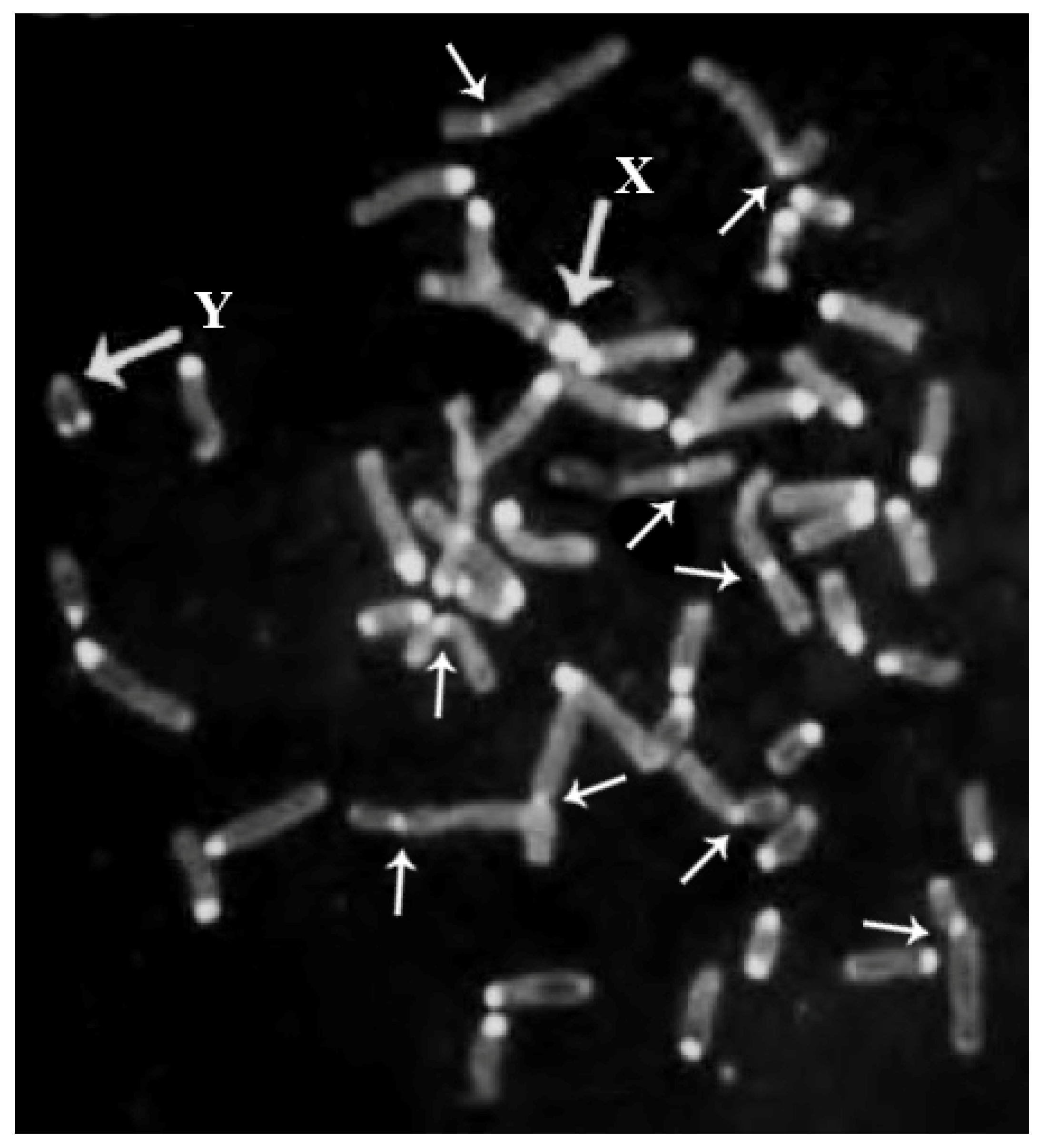
The river buffalo karyotype (2n = 50; Figure 5) has five biarmed pairs (submetacentric chromosomes). The remaining chromosomes are acrocentric, including both the X, being the largest acrocentric chromosome, and Y, the latter being one of the smallest acrocentric chromosomes. The five biarmed pairs originated via five centric fusion translocations involving the following 10 cattle (ancestral bovid) homologous chromosomes, according to both the standard river buffalo karyotype [47] and ISCNDB2000 [52]: BBU1 (1;27), BBU2 (2;23), BBU3 (8;19), BBU4 (5;28), BBU5 (16;29) (Table 1) (Figure 4). HC (C-band) losses occur along the centromeres involved in these fusions [58] (Figure 1).
In B. mindorensis (tamaraw) (2n = 46, FN = 56), six biarmed chromosomes resulted from centric fusions, five of which were identical to those of swamp buffalo, with one additional rob (4;14) [72] (Table 1). Therefore, B. mindorensis was probably derived from swamp buffalo.
In the Bubalus genus, we also have the anoa buffalo, with two subspecies: the B. depressicornis depressicornis (lowland anoa) and B. depressicornis quarlesi (mountain anoa), both with 2n = 48 and FN = 60 [65]. These species are the smallest buffaloes in the world. The karyotypes of these species are analogous to that of B. bubalis, with six biarmed chromosome pairs. Among those, four are identical to those present in Bubalus, the river type (BBU1, BBU2, BBU3, and BBU4), while the other two biarmed pairs of anoa are different because they involve cattle homologous chromosomes 11;20 and 17;15 [65] (Table 1). However, strong evidence is provided by phylogenetic and molecular dating analyses that the lowland anoa, river buffalo, and swamp buffalo are three distinct taxa that speciated rapidly during the Pleistocene epoch [70].
Other chromosome regions that play an essential role in the karyotype evolution of bovids are NORs. These are specific chromosome regions in which genes codifying for ribosomal RNA are intensely present. They are visualized using Ag-NOR staining [11] or FISH mapping using specific ribosomal probes [73]. The former allows visualization of only the active NORs (NORs that have organized at least one nucleolus), while the latter includes all NORs. NORs in bovids are generally available at the telomeres of autosomes—regions usually uninvolved in chromosome rearrangement (in contrast to centromeric regions). Additionally, all bovid chromosome arms were conserved. Therefore, we expected to find NORs at identical homologous chromosomes in various bovid species, but this was not the case. Indeed, NORs differ in number and chromosome position between species, especially when considering different genera [57,65]. In Table 2, the data on the NORs of the buffalo species and cattle (for comparison) are summarized. As shown, the number of NORs differs between river (with six nucleolus organizer (NO) chromosomes) and swamp (with five NO chromosomes) buffaloes. In the latter, the NORs present at the telomeres of BBU4p were lost during tandem fusion translocation originating from swamp buffalo chromosome 1, as reported above [61]. However, the remaining five NORs were conserved in the same NO chromosomes of the two subspecies. When compared with cattle, we observed that only two NORs were conserved in homologous chromosomes between cattle and water buffalo (3 and 19 of cattle corresponding to the buffalo chromosomes 6 and 3p, respectively) (Table 2). In S. c. caffer, only four NO chromosomes (2p, 22, 25, and 28) were found in the homologous chromosomes of the water buffalo [65], as shown in Table 2.
All species belonging to the subfamily Bovinae (including the B. bubalis) conserved the same autosomes 9 and 14, whereas in the other bovid species of the remaining subfamilies, these two chromosomes evolved via a simple translocation event from the proximal BTA9 region to the proximal BTA14 region [57,84]. In addition, some micro-rearrangements occurring among domestic bovids (including the river buffalo) have been found using a combination of bioinformatics techniques and the physical mapping of DNA markers using the FISH technique [85,86].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 2.
Number of nucleolus organizer regions (NORs) and nucleolus organizer (NO) chromosomes in cattle and buffaloes. Homologous cattle NO chromosomes in buffaloes are reported between parentheses according to [52,65,87].
| Species | NORs (Number of Chromosome Pairs) | NO-Chromosomes (Cattle Homologous Chromosomes Are Reported between Parentheses) |
|---|---|---|
| Bos taurus (cattle) | 5 | 2, 3, 4, 11, 19 |
| Bubalus b. (river) | 6 | 3p (19), 4p (28), 6(3), 21(22), 23(26), 24(25) |
| Bubalus b. (swamp) | 5 | 3p (19), 6(3), 20(22), 22(26), 24(25) |
| Syncerus caffer caffer | 4 | 2p (3), 22(22), 25(25), 28(28) |
2.2. Sex Chromosomes
Unlike the autosomes, the sex chromosomes of bovids evolved via complex chromosomal rearrangements, especially when comparing the Bovinae subfamily with the remaining members of the Bovidae family. Indeed, the X and Y chromosomes of domestic bovids differ in shape, size, and gene order [36,57]. In particular, the X chromosome in Bovinae, including the water buffalo, differs from the X chromosome in Caprinae by at least four chromosomal transpositions—including the centromere, with an inversion [35,57,88]. The water buffalo X chromosome is the largest acrocentric chromosome of the buffalo karyotype, and it is larger than cattle X (submetacentric) due to the presence of a large block of centromeric HC (C-band), with an additional positive C-band proximally located, whereas the cattle X chromosome is C-band negative [57]. In previous studies, it was hypothesized that the X chromosomes of cattle and river buffalo differ by pericentric inversion [62,89].
After high-resolution chromosome banding and detailed FISH mapping using several markers along the X chromosomes of both species, the gene order in the X chromosomes was the same. Still, the X chromosomes were different due to centromere repositioning (or transposition), with HC loss from river buffalo to cattle X chromosomes [35,57,88]. This finding is more likely when considering that the buffalo X chromosome is similar to the ancestral bovid karyotype [57].
Moreover, the Y chromosomes of cattle and buffaloes differ in size (more prominent in buffaloes) and shape (small submetacentric in cattle and small acrocentric in buffalo). These features occur because of a pericentric inversion differentiating the two chromosomes, with loss of HC from the buffalo to the cattle Y chromosome, as demonstrated by FISH mapping with some molecular markers [33,57]. This chromosome is generally heterochromatic (C-band positive) or has a telomeric positive C-band (Figure 2), depending on the degree of chromosomal denaturation in the C-banding technique [57]. Furthermore, the difference in size between the Y chromosomes of cattle and buffalo is only due to HC being more significant in the buffalo Y chromosome.
3. Clinical Cytogenetics
Clinical cytogenetics is a crucial research field of animal cytogenetics. In this research field, the study of the relationships between chromosomal abnormalities and their effects on fertility and body conformation is attempted. Since the discovery of rob (1;29) in a Swedish red cattle breed [90] and the demonstration of its deleterious effects on fertility [80,81], the clinical cytogenetics applied to domestic animals have spread throughout the world to study many domestic species, including the water buffalo. However, the water buffalo (essentially, the river buffalo) has been studied by only a few cytogenetic laboratories.
Numerical autosomal abnormalities have rarely been found in domestic animals, because the animals have abnormal body conformation. For this reason, these abnormalities are eliminated directly by the breeders. Conversely, numerical sex chromosome abnormalities are more tolerated by the species, because one of their X chromosomes is genetically inactivated [91], although some genes seem to escape gene inactivation [92]. However, numerical sex chromosome abnormalities are often correlated with sterility or low fertility, especially in females [3,4,5,6,7]. Balanced chromosomal abnormalities are generally correlated with reduced fertility due to unbalanced gametes originating during meiosis to form trivalent (centric fusions) or quadrivalent (reciprocal translocations) configurations. Subsequently, unbalanced gametes and embryos generally die in early embryonic life [3,4,5,6,7].
In Table 3, the most critical chromosomal abnormalities found so far in river buffalo and their effects on fertility are summarized. As shown in Table 3, only three autosomal chromosome abnormalities have been found in river buffaloes [93,94,95]: The first case was found in a cow with reduced fertility [93]. The second was found in a famous bull known as Magnifico [94]. A complex chromosomal rearrangement originated in both chromosomal abnormalities: fission of river buffalo chromosome 1 (BBU1) and subsequent centric fusion between chromosome BBU1p and BBU23 in the cow [93], and with BBU18 in the bull [94]. The cow had reduced fertility, producing only two lactations and calves in five years of reproductive life. One male calf had a normal karyotype, and the female carried the same translocation. After that, both females were eliminated from the reproduction farm.
The bull was famous for its high genetic value, and the use of its semen in artificial inbreeding (AI) resulted in its numerous progeny. When its chromosomal abnormality was also found in several progeny animals [94], the Italian National Breeders of Buffalo Species Association (ANASB) stakeholders decided to remove this bull from reproduction. However, in the analysis of the total and motile sperm fractions of this bull via sperm FISH, translocation was suggested to have minimal effects on the aneuploidy of its gametes. Therefore, there were also minimal effects on the reproductive abilities of the bulls [83]. These studies are demonstrations that it is crucial to obtain the banded karyotype of all bulls or males addressed to reproduction before their use as reproducers, especially in AI. The third case was reported in a Murrah buffalo bull, which had mosaicism (2n = 50, XY/2n = 50, XY, 3q-), with a partial deletion of the BBUq arm in some cells [95]. Still, the most common chromosomal abnormalities found in river buffalo involve sex chromosomes, in which mosaicism XX/XY (freemartin) constitutes the majority of cases (Table 3).
In detail, four cases of X-chromosome trisomy were found in both Murrah (two cases) and Italian Mediterranean (two cases) breeds (Table 3). In three cases, females were sterile because of severe damage to internal sex adducts [96,97,98]. In one case, the female had reduced fertility because it had only two lactations in 10 years [99] (Table 3).
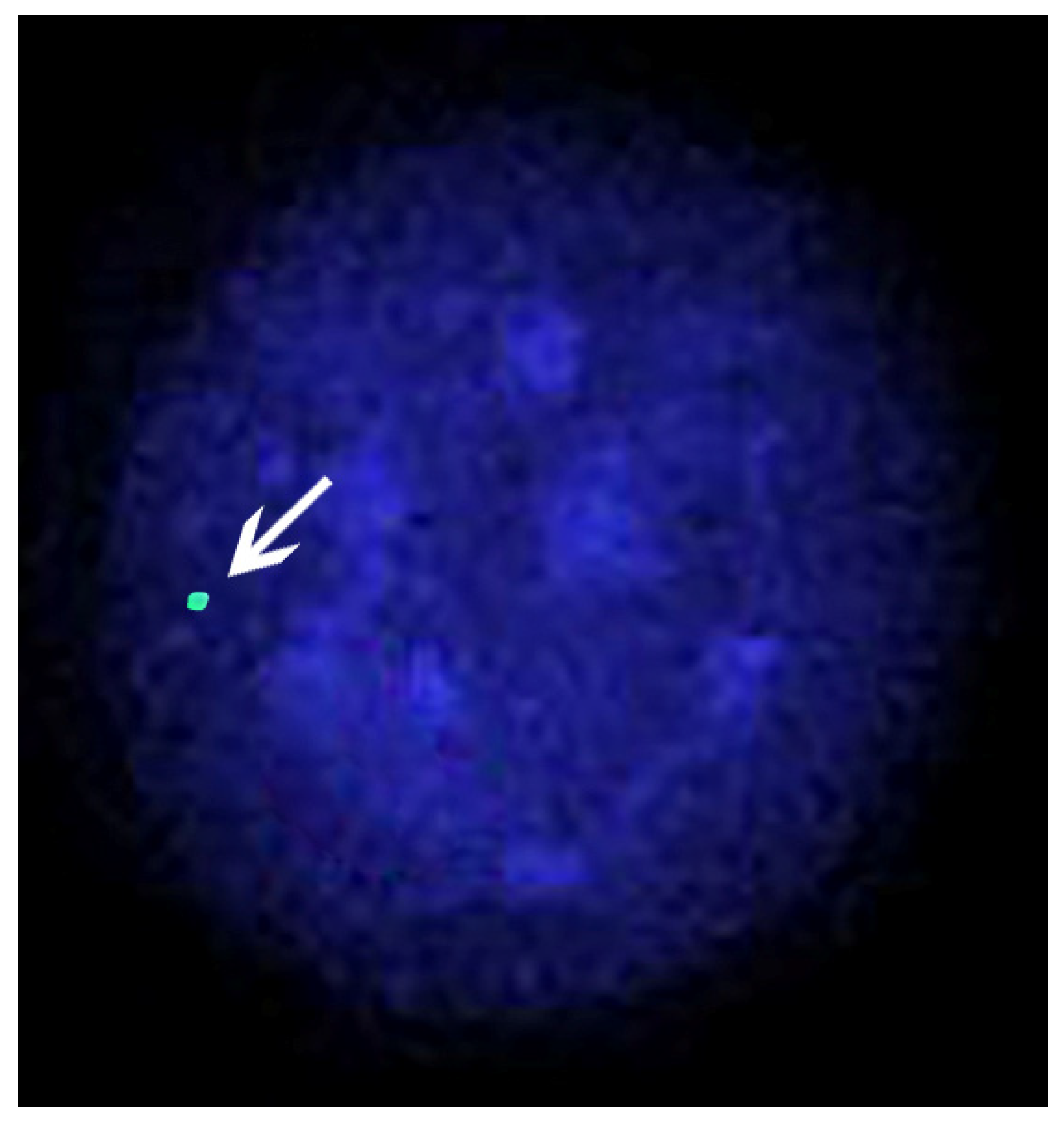
Four cases of X-chromosome monosomy were found in river buffaloes: two were in the Murrah breed [100,101], and two were in the Italian Mediterranean breed [97,98]. In all cases, the individuals were affected by gonadal dysgenesis (Table 3). In Figure 6, an interphase nucleus of a river buffalo female affected by X-chromosome monosomy is shown.
As shown in Table 3, most sex chromosome abnormalities are XX-XY mosaicisms (freemartin). This type of chromosomal abnormality occurs when placental anastomoses originate between heterosexual twins. Approximately 90% of these twins are freemartin in cattle, and most females are generally sterile as a result of severe damage to their internal sex organs [3,102]. These deleterious effects on internal sex adducts are because of the Y chromosome and the male-determining regions occurring in the blood cells. This phenomenon comprises the following two subsequent events: (1) placental anastomosis occurs 20 days before sex differentiation, and (2) male cell differentiation occurs one week earlier in males than in females [103]. For this reason, deleterious effects on the internal sex organs are much more frequent in female twins than in males, although some damage to male organs has also been reported in male twins [104,105]. However, the phenomenon of freemartinism is related to the percentage of twins being relatively high in cattle (varying from 2 to 4% in dairy breeds, reaching higher values (6%) in older cows) [106]. In buffaloes, the percentage of twins is meager (0.14%; [107]) but, unlike cattle, most females are found to be freemartin from a single birth. Indeed, as shown in Table 3, only four twins (two births) were born in both males and females, and all remaining cases were only female freemartins from a single birth. This finding means that the males died during early embryonic life and were adsorbed.
These data lead us to two important conclusions: (1) in buffaloes, the actual percentage of twins (at least in early embryonic life) is much higher than that observed; and (2) since most of the cases are from single birth, the breeder cannot realize that the female is a freemartin case, and keeps it in the farm as a normal individual for years, causing severe economic damage to the farm. Only after the female reaches reproductive age with no pregnancy, despite the presence of a bull, can veterinarian controls (e.g., rectal palpation or eco-graphic analyzes) and proper cytogenetic and genetic controls (e.g., PCR with specific male markers) be used to reveal freemartinism. However, male traits (e.g., prominent withers, larger horn base circumference, tight pelvis) were observed (Table 3); thus, both breeders and veterinarian doctors should pay careful attention to the external body traits of females at a young age in order to perform an earlier diagnosis of freemartinism via cytogenetic or molecular (PCR) analyses.
At least for the cytogenetic controls performed in the Italian Mediterranean breed of all studied females with reproductive problems, ~20% were found to be affected by sex chromosome abnormalities [97,98] (Table 3). These data underline the necessity of carefully investigating all females with reproductive problems in order to drastically reduce the economic damage caused by keeping females who will never produce calves or lactations for years. In this case, a minimum of 50 cells must be studied for each animal with reduced fertility using the C-banding technique, because sex chromosomes can be easily detected using this technique [11,57]. Indeed, the X chromosome is the largest acrocentric chromosome with the largest centromeric positive C-band, with an additional one located proximally (Figure 1). At the same time, the Y chromosome appears as completely heterochromatic (C-band positive) or with a positive C-band only situated distally (Figure 1), while all remaining acrocentric chromosomes are C-band positive in the centromeric regions [57,58] (Figure 1). Additional banding (preferably R-banding) and FISH (if necessary) techniques must be applied to animals affected by numerical autosome and structural chromosome abnormalities involving autosomes and sex chromosomes. However, using C- and R-banding techniques for all males addressed to reproduction and females with reproductive problems is highly suggested.
Chromosomes have also been used to establish the level of genome stability (or instability) in the cells of animals exposed to mutagens in vivo or in vitro, by using cytogenetic tests such as the CA test (chromatid or chromosome breaks), SCE test (sister chromatid exchange), MN test (micronuclei), comet assay test, and telomere test by RLTL. Some of these techniques have been applied in river buffalo, and are reported in Table 4; the type of test and the main results achieved are also indicated.
Table 3.
Cases of river buffaloes (breed/country) per year affected by chromosomal abnormalities, their phenotypic effects, and references.
Table 3.
Cases of river buffaloes (breed/country) per year affected by chromosomal abnormalities, their phenotypic effects, and references.
| Breed/Country | Chromosome Abnormality | Sex | Phenotypic Effects | References |
|---|---|---|---|---|
| Murrah/India | XX/XY mosaicism (Freemartin) | F | Cell mosaicism found in a a triplet birth; female with internal sex damage | [108] |
| Murrah/India | X-trisomy (2n = 51, XXX) | F | Normal body conformation, reduced fertility (only two lactations in 10 years) | [99] |
| Murrah/India | X-trisomy (2n = 51, XXX) | F | Sterile (damages to internal sex structures) | [96] |
| Murrah/India | X-monosomy (2n = 49, X) | Gonadal disgenesis (sterility) | [100] | |
| Murrah/India | X-monosomy (2n = 49, X) | Gonadal disgenesis (sterility) | [101] | |
| Italian Mediterranean/ Italy | X-monosomy (2n = 49, X) | F | Normal body conformation and external genitalia; small uterine body; ovaries not detectable; sterile | [109] |
| Ital. Mediterranean/ Italy | XY-sex reversal (2n = 50, XY) | F | Slight hypoplasia of derivative Muller’s ducts, as well as small cervix uteri and ovary structure; sterile | [110] |
| Ital. Mediterranean/ Italy | X-trisomy (2n = 51, XXX) | F | Normal weight but presence of male traits (prominent withers, large horn base circumference); normal vagina and clitoris; atrophy of internal sex adducts; sterile | [97] |
| Ital. Mediterranean/ Italy | XY-sex reversal (2n = 50, XY) | F | Normal weight with presence of male traits (prominent withers, large horn base circumference); close vagina; absence of internal sex adducts | [97] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (Freematrtin) twin | F | Normal body conformation and external genitalia; atrophy of internal sex adducts; sterile | [111] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (Freemartin), twin | M | Normal body conformation and external genitalia | [111] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (Freemartin) | F | Presence of some male traits (tight pelvis, large horn base circumference); normal external genitalia; serious atrophy of Muller’s ducts; small ovaries; sterile | [111] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (Freemartin) | F | Normal body conformation; normal vulva, vagina, and clitoris; atrophy of internal sex adducts; ovaries not detectable; sterile. | [111] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (Freemartin) | F | Normal body conformation and external genitalia; atrophy of internal sex adducts; ovaries not detectable; sterile | [111] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (Freemartin) twin | M | Normal body conformation and penis; one testis much smaller than the other. | [111] |
| Ital. Mediterranean /Italy | XX/XY mosaicism (Freemartin) twin | F | Body conformation with some male traits (tight pelvis) and unusual horns (thin); normal vulva, vagina, and clitoris; lack of internal sex adducts; sterile. | [111] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (Freemartin) | F | Normal body conformation; normal vulva, vagina, and clitoris; atrophy of internal sex adducts; ovaries not detectable; sterile. | [111] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (Freemartin) | F | Normal body conformation and external genitalia; atrophy of internal sex adducts; sterile | [111] |
| Murrah/India | XXY-syndrome 2n = 50,Y, rob(X;X) | M | Testicular hypoplasia; azoospermic | [112,113] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (freemartin) | M | Normal body conformation and external genitalia, with one testis smaller than the other; fertile (with progeny) | [98] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (freemartin) | F | Normal body conformation and external genitalia; closed vagina with lack of internal sex adducts; sterile | [98] |
| Ital. Mediterranean/ Italy | X-monosomy (2n = 49, X) | F | Normal body conformation; clitoris larger than normal; small uterine cervix; uteri horns very thin; gonads absent; sterile | [98] |
| Ital. Mediterranean/ Italy | X-trisomy (2n = 51, XXX) | Normal body conformation; atrophy of internal sex adducts; sterile | [98] | |
| Ital Mediterranean/ Italy | XX/XY mosaicism (freemartin) | F | Normal body conformation and external genitalia; atrophy of internal sex adducts; small ovaries; sterile | [98] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (freemartin) | F | Presence of male traits (tight pelvis) and thin horns; normal external genitalia and vagina; lack of internal sex adducts; sterile | [98] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (freemartin) | F | Normal body conformation and external genitalia; absence of internal sex adducts with closed vagina; sterile | [98] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (freemartin) | F | Normal body conformation and external genitalia; atrophy of internal sex adducts; small ovaries; sterile | [98] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (freemartin) | F | Normal body conformation and external genitalia; small uterine body; sterile | [98] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (freemartin) | F | Normal body conformation and external genitalia; small uterine body and ovaries; sterile | [98] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (freemartin) | F | Normal body conformation and external genitalia; small uterine body; sterile | [98] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (freemartin) | F | Normal vulva with large clitoris; atrophy of internal sex adducts; sterile | [98] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (freemartin) | F | Some male traits (pelvis slightly thin); normal external genitalia; closed vagina with absence of internal sex adducts; sterile | [98] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (freemartin) | F | Head with slight male traits, but with horns smaller than normal (horn base circumference 26 cm); uterine body incomplete; presence of a small left uterine horn without ovary; sterile | [98] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (freemartin) | F | Head with male traits, normal external genitalia; internal sex adducts showing a very small uterine body developed with a draft right uterine horn and atrophic uterine left horn with a draft ovary; sterile | [98] |
| Ital. Mediterranean/ Italy | XX/XY mosaicism (freemartin) | F | Normal body conformation and external genitalia, with a small clitoris, and atrophic internal adducts, with both uterine body and uterine horns being as fibroid cords without lumen | [98] |
| Ital. Mediterranean/ Italy | rob(1p;23), 2n = 50 | F | Reduced fertility (only two lactations in five years); second female calf carrying the same abnormality | [98] |
| Ital. Mediterranean/ Italy | rob(1p;18), 2n = 50 | M | Famous bull eliminated from reproduction; the abnormality was also found in some of the progeny; sperm FISH in the motile fraction sperm revealed limited effect on aneuploidy in the gametes | [94] [83] |
| Murrah/India | Mosaicism 20 = 50, XY/2n = 50, XY, 3q- | M | Phenotypically normal and fertile bull aged between 40 and 42 months. Not yet used for reproduction | [95] |
Table 4.
Cytogenetic test applied to river buffalo cells or sperm after in vivo (natural exposure) or in vitro exposure to potential mutagens; results and references.
Table 4.
Cytogenetic test applied to river buffalo cells or sperm after in vivo (natural exposure) or in vitro exposure to potential mutagens; results and references.
| Breed/Country | Cytogenetic Test | In Vivo/In Vitro Exposure | Results | Reference |
|---|---|---|---|---|
| Ital. Mediterranean/ Italy | SCE | In vitro | Normal SCE baseline in blood cells | [114] |
| Mediterranean/Egypt | Chromosome breaks, SCE | In vivo | Increased chromosomal fragility in buffaloes raised in polluted areas | [115] |
| Ital. Mediterranean/ Italy | SCE | In vitro | Chromosome instability induced in the X chromosome by exposure to Mitomicin C | [116] |
| Murah, Jaffarabadi, Italian Mediterranea/Brazil | Chromosome breaks | In vivo | Fragility in the X chromosome | [117] |
| Mediterranean/Egypt | Chromosome breaks, SCE, micronuclei | In vitro | Clastogenic effects of the fasciolicide drug Fasinex | [45] |
| Ital. Mediterranean/ Italy | DNA polymerase alpha inhibition by aphidicolin | In vitro | Fragile sites mainly found in specific chromosomes | [118] |
| Ital. Mediterranean/ Italy | Chromosome breaks, SCE, aneuploidy | In vitro | Chromosomal instability in calves with limb malformation (transversal hemimelia—TH) | [46] |
| Murrah/India | DNA sperm fragmentation | In vivo | Normal baseline of buffalo DNA sperm fragmentation | [119] |
| Ital. Mediterranean/ Italy | Chromosome breaks, SCE | In vivo exposure to dioxins | Chromosome fragility | [42] |
| Murrah/India | DNA sperm fragmentation | In vitro (titanium oxide (TiO2) nanoparticles (NPs exposure) | Cytotoxic effect on buffalo spermatozoa | [44] |
| Ital. Mediterranean/ Italy | SCE | In vitro | No chromosomal fragility in cells exposed to furocumarin extracts | [43] |
| Italian Mediterranean/ Italy | Chromosome breaks, SCE, micronuclei, RLTL | In vivo | No chromosomal fragility in blood cells of buffaloes raised in urban and rural areas | [41] |
4. Molecular Cytogenetics
Molecular cytogenetics was first applied to human chromosomes when specific DNA probes were hybridized using a new and advanced technique—fluorescence in situ hybridization (FISH) [120,121,122]. The DNA probes are made with cDNA, which is only applicable when the target gene is multicopy. Nevertheless, more generally, they may also be made with genomic DNA of different sizes, such as cosmids (DNA insert sizes of 20–40 kb) or, more commonly, bacterial artificial chromosomes (BACs; DNA insert sizes of 100–300 kb). Therefore, with these DNA probes, stronger hybridization signals are able to be visualized. Depending on the type of probe (mapping a specific chromosome region with the classical FISH), entire chromosomes, chromosome arms (painting probes; Zoo-FISH), specific chromosome regions, or whole chromosomes will be fluorescent in the dark field (Figure 3c–e). Chromosome painting probes are generally obtained by cell sorter chromosomes or by chromosome microdissection. The only (partial) chromosome libraries available in river buffalo are those obtained via chromosome microdissection [25,123].
Generally, the FISH technique allows us to (1) physically map genetic loci on specific chromosome regions [10,124,125]; (2) compare entire chromosome regions between related and unrelated species (Zoo-FISH) [20,21,22,23,24,25,26,27,28,126]; (3) establish the gene order along the chromosomes using detailed FISH-mapping data, which are adequate to detect chromosomal abnormalities (e.g., inversions, deletion, reciprocal, and Robertsonian translocations) (reviewed in [3,4,5,6,7]), or to compare different species on the basis of the gene order in homologous chromosomes or chromosome regions [24,32,34,35,36,127,128]; and (4) detect chromosome aneuploidy or trisomy in both metaphase [97,109] and interphase nuclei of somatic (Figure 6) and germinal cells [4,83,129].
The FISH technique was first applied to domestic animals to physically map the beta-casein gene on cattle chromosomes [124], or to correctly identify a chromosomal abnormality in cattle [130]. Comparative FISH mapping was first applied to domestic bovids to map the major histocompatibility complex (MHC) locus on cattle and river buffalo [67], and the omega and trophoblast interferon genes in cattle, river buffalo, sheep, and goats [66]. In these studies and others that followed, the high degree of chromosome and loci conservation among related bovids was confirmed [32,33,57,68]. The FISH technique was used to physically map several loci in river buffalo chromosomes, especially after the publication of the standard river buffalo karyotype [47]. The most complete cytogenetic map for the river buffalo was reported in [10], where 308 loci were mapped. However, the FISH technique has also been used in river buffalo to (1) compare human and river buffalo genomes via Zoo-FISH, using human chromosome libraries as probes, which is helpful in detecting the human chromosome regions conserved in river buffalo chromosomes [23]; (2) resolve evolutionary events in which the river buffalo was differentiated from other bovids [35,57,68,88]; (3) correctly identify the river buffalo chromosomes involved in chromosomal abnormalities [93,94,109,110,131]; (4) support the radiation hybrid (RH) maps performed in river buffalo [132,133,134]; and (5) check the fertility degree of bull semen carrying a chromosomal abnormality via sperm FISH [83].
As reported previously, Zoo-FISH is helpful for checking the conserved chromosome regions between related and unrelated species [20,21,22,23,24,25,26,27,28]. Still, the gene order between species is unknown, especially between distantly related mammals (i.e., humans–bovids). This issue can be easily resolved via FISH mapping of individual loci along the chromosomes, allowing us to reveal the correct gene order in chromosome regions between related and unrelated species in order to precisely establish conserved chromosome regions in several species, including the water buffalo [35,57,68,88].
5. Conclusions
The water buffalo remains of interest for its strategic and economic importance to the large human population raising this species. The chromosomes remain a fascinating material to differentiate not only the various species of buffalo in the world, but also the results of crosses between different subspecies. From an evolutionary point of view, cytogenetic studies of wild buffalo (B. arnee) should be undertaken in order to better establish the origins of various species of water buffaloes. Crosses between river and swamp buffaloes are important to noticeably increase milk production, although problems can arise as a result of the limited food available in swamp buffalo farms supporting higher milk production, as well as reproductive problems during the meiosis of hybrids with 49 chromosomes, as discussed above. Cytogenetic controls—at least for all males addressed to reproduction and females with reproductive problems (e.g., lack of estrus in reproductive age, larger than normal interbirth interval)—should be implemented to add genetic and economic value to the water buffalo food chain. Collaboration between breeders, veterinarian doctors, and cytogenetic labs from various countries is essential in order to reach this goal. More progress in the genetic improvement of buffalo cytogenetics can be obtained using the CGH array, as done in cattle, to reveal genomic DNA losses during the formation of chromosomal abnormalities. It is still essential to extend the water buffalo cytogenetic maps in order to noticeably increase the number of mapped loci (type I and type II). Such an extension will better anchor hybrid radiation maps to specific chromosome regions, and help to allocate genetic sequences of milk and meat production genes.
Author Contributions
Conceptualization, L.I. and P.P.; writing—original draft preparation, L.I., A.I. and P.P.; writing—review and editing, L.I. and A.I. All authors have read and agreed to the published version of the manuscript.
Funding
The study has been supported by the project PON1_486 GENOBU.
Institutional Review Board Statement
The Institutional Review Board statement was not required.
Data Availability Statement
Data are contained within the article.
Acknowledgments
We are grateful to L.C., Cruz, Philippine Carabao Center, Munoz, Nueva Ecija, Philippines, for blood samples of river swamp buffalo hybrids.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Minervino, A.; Zava, M.; Vecchio, D.; Borghese, A. Bubalus bubalis: A Short Story. Front. Vet. Sci. 2020, 7, 570413. [Google Scholar] [CrossRef]
- Zhang, Y.; Colli, L.; Barker, J. Asian water buffalo: Domestication, history and genetics. Anim. Genet. 2020, 51, 177–191. [Google Scholar] [CrossRef]
- Ducos, A.; Revay, T.; Kovacs, A.; Hidas, A.; Pinton, A.; Bonnet-Garnier, A.; Molteni, L.; Slota, E.; Switonski, M.; Arruga, M.V.; et al. Cytogenetic screening of livestock populations in Europe: An overview. Cytogenet. Genome Res. 2008, 120, 26–41. [Google Scholar] [CrossRef]
- Iannuzzi, A.; Parma, P.; Iannuzzi, L. Chromosome Abnormalities and Fertility in Domestic Bovids: A Review. Animals 2021, 11, 802. [Google Scholar] [CrossRef] [PubMed]
- Szczerbal, I.; Switonski, M. Clinical Cytogenetics of the Dog: A Review. Animals 2021, 11, 947. [Google Scholar] [CrossRef] [PubMed]
- Bugno-Poniewierska, M.; Raudsepp, T. Horse Clinical Cytogenetics: Recurrent Themes and Novel Findings. Animals 2021, 11, 831. [Google Scholar] [CrossRef]
- Donaldson, B.; Villagomez, D.; King, W.A. Classical, Molecular, and Genomic Cytogenetics of the Pig, a Clinical Perspective. Animals 2021, 11, 1257. [Google Scholar] [CrossRef]
- Hayes, H.; Petit, E.; Lemieux, N.; Dutrillaux, B. Chromosomal localization of the ovine beta-casein gene by non-isotopic in situ hybridization and R-banding. Cytogenet. Cell Genet. 1992, 61, 286–288. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L.; Di Meo, G.P.; Perucatti, A.; Schibler, L.; Incarnato, D.; Gallagher, D.; Eggen, A.; Ferretti, L.; Cribiu, E.P.; Womack, J. The river buffalo (Bubalus bubalis, 2n = 50) cytogenetic map: Assignment of 64 loci by fluorescence in situ hybridization and R-banding. Cytogenet. Genome Res. 2003, 102, 65–75. [Google Scholar] [CrossRef]
- Di Meo, G.P.; Perucatti, A.; Floriot, S.; Hayes, H.; Schibler, L.; Incarnato, D.; Di Berardino, D.; Williams, J.; Cribiu, E.; Eggen, A.; et al. An extended river buffalo (Bubalus bubalis, 2n = 50) cytogenetic map: Assignment of 68 autosomal loci by FISH-mapping and R-banding and comparison with human chromosomes. Chromosome Res. 2008, 16, 827–837. [Google Scholar] [CrossRef] [Green Version]
- Iannuzzi, L.; Di Berardino, D. Tools of the trade: Diagnostics and research in domestic animal cytogenetics. J. Appl. Genet. 2008, 49, 357–366. [Google Scholar] [CrossRef] [PubMed]
- De Lorenzi, L.; Genualdo, V.; Perucatti, A.; Iannuzzi, A.; Iannuzzi, L.; Parma, P. Physical mapping of 20 unmapped fragments of the btau_4.0 genome assembly in cattle, sheep and river buffalo. Cytogenet. Genome Res. 2013, 140, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L.; Rangel-Figueiredo, T.; Di Meo, G.P.; Ferrara, L. A new Robertsonian translocation in cattle, rob(15;25). Cytogenet. Cell Genet. 1992, 59, 280–283. [Google Scholar] [CrossRef]
- Iannuzzi, L.; Rangel-Figueiredo, T.; Di Meo, G.P.; Ferrara, L. A new centric fusion translocation in cattle, rob(16;18). Hereditas 1993, 119, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L.; Molteni, L.; Di Meo, G.P.; Perucatti, A.; Lorenzi, L.; Incarnato, D.; De Giovanni, A.; Succi, G.; Gustavsson, I. A new balanced autosomal reciprocal translocation in cattle revealed by banding techniques and human-painting probes. Cytogenet. Cell Genet. 2001, 94, 225–228. [Google Scholar] [CrossRef]
- Di Meo, G.P.; Perucatti, A.; Chaves, R.; Adega, F.; De Lorenzi, L.; Molteni, L.; De Giovanni, A.; Incarnato, D.; Guedes-Pinto, H.; Eggen, A.; et al. Cattle rob(1;29) originating from complex chromosome rearrangements as revealed by both banding and FISH-mapping techniques. Chromosome Res. 2006, 14, 649–655. [Google Scholar] [CrossRef]
- Molteni, L.; Perucatti, A.; Iannuzzi, A.; Di Meo, G.P.; De Lorenzi, L.; De Giovanni, A.; Incarnato, D.; Succi, G.; Cribiu, E.; Eggen, A.; et al. A new case of reciprocal translocation in a young bull: Rcp(11;21)(q28;q12). Cytogenet. Genome Res. 2007, 116, 80–84. [Google Scholar] [CrossRef] [Green Version]
- De Lorenzi, L.; Rossi, E.; Genualdo, V.; Gimelli, S.; Lasagna, E.; Perucatti, A.; Iannuzzi, A.; Parma, P. Molecular characterization of Xp chromosome deletion in a fertile cow. Sex. Dev. 2012, 6, 298–302. [Google Scholar] [CrossRef]
- De Lorenzi, L.; Iannuzzi, A.; Rossi, E.; Bonacina, S.; Parma, P. Centromere Repositioning in Cattle (Bos taurus) Chromosome 17. Cytogenet. Genome Res. 2017, 151, 191–197. [Google Scholar] [CrossRef]
- Hayes, H. Chromosome painting with human chromosome-specific DNA libraries reveals the extent and distribution of conserved segments in bovine chromosomes. Cytogenet. Cell Genet. 1995, 71, 168–174. [Google Scholar] [CrossRef]
- Chowdhary, B.P.; Frönicke, L.; Gustavsson, I.; Scherthan, H. Comparative analysis of the cattle and human genomes: Detection of ZOO-FISH and gene mapping-based chromosomal homologies. Mamm. Genome 1996, 7, 297–302. [Google Scholar] [CrossRef]
- Frönicke, L.; Chowdhary, B.P.; Scherthan, H.; Gustavsson, I. A comparative map of the porcine and human genomes demonstrates ZOO-FISH and gene mapping-based chromosomal homologies. Mamm. Genome 1996, 7, 285–290. [Google Scholar] [CrossRef]
- Iannuzzi, L.; Di Meo, G.P.; Perucatti, A.; Bardaro, T. ZOO-FISH and R-banding reveal extensive conservation of human chromosome regions in euchromatic regions of river buffalo chromosomes. Cytogenet. Cell Genet. 1998, 82, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L.; Di Meo, G.P.; Perucatti, A.; Incarnato, D. Comparison of the human with the sheep genomes by use of human chromosome-specific painting probes. Mamm. Genome 1999, 10, 719–723. [Google Scholar] [CrossRef] [PubMed]
- Pauciullo, A.; Perucatti, A.; Cosenza, G.; Iannuzzi, A.; Incarnato, D.; Genualdo, V.; Di Berardino, D.; Iannuzzi, L. Sequential cross-species chromosome painting among river buffalo, cattle, sheep and goat: A useful tool for chromosome abnormalities diagnosis within the family Bovidae. PLoS ONE 2014, 9, e110297. [Google Scholar] [CrossRef] [Green Version]
- Iannuzzi, A.; Pereira, J.; Iannuzzi, C.; Fu, B.; Ferguson-Smith, M. Pooling strategy and chromosome painting characterize a living zebroid for the first time. PLoS ONE 2017, 12, e0180158. [Google Scholar] [CrossRef] [Green Version]
- Romanenko, S.A.; Serdyukova, N.A.; Perelman, P.L.; Pavlova, S.V.; Bulatova, N.S.; Golenishchev, F.N.; Stanyon, R.; Graphodatsky, A.S. Intrachromosomal Rearrangements in Rodents from the Perspective of Comparative Region-Specific Painting. Genes 2017, 8, 215. [Google Scholar] [CrossRef] [Green Version]
- Romanenko, S.A.; Fedorova, Y.E.; Serdyukova, N.A.; Zaccaroni, M.; Stanyon, R.; Graphodatsky, A.S. Evolutionary rearrangements of X chromosomes in voles (Arvicolinae, Rodentia). Sci. Rep. 2020, 10, 13235. [Google Scholar] [CrossRef]
- Escudeiro, A.; Adega, F.; Robinson, T.J.; Heslop-Harrison, J.S.; Chaves, R. Analysis of the Robertsonian (1;29) fusion in Bovinae reveals a common mechanism: Insights into its clinical occurrence and chromosomal evolution. Chromosome Res. 2021. [Google Scholar] [CrossRef]
- Iannuzzi, L.; Gallagher, D.S., Jr.; Di Meo, G.P.; Yang, Y.; Womack, J.E.; Davis, S.K.; Taylor, J.F. Comparative FISH-mapping of six expressed gene loci to river buffalo and sheep chromosomes. Cytogenet. Cell Genet. 1999, 84, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L.; Di Meo, G.P.; Perucatti, A.; Schibler, L.; Incarnato, D.; Ferrara, L.; Bardaro, T.; Cribiu, E.P. Sixteen type I loci from six human chromosomes were comparatively fluorescence in-situ mapped to river buffalo (Bubalus bubalis) and sheep (Ovis aries) chromosomes. Chromosome Res. 2000, 8, 447–450. [Google Scholar] [CrossRef]
- Di Meo, G.P.; Perucatti, A.; Schibler, L.; Incarnato, D.; Ferrara, L.; Cribiu, E.P.; Iannuzzi, L. Thirteen type I loci from HSA4q, HSA6p, HSA7q and HSA12q were comparatively FISH-mapped in four river buffalo and sheep chromosomes. Cytogenet. Cell Genet. 2000, 90, 102–105. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, G.P.; Perucatti, A.; Floriot, S.; Incarnato, D.; Rullo, R.; Caputi Jambrenghi, A.; Ferretti, L.; Vonghia, G.; Cribiu, E.; Eggen, A.; et al. Chromosome evolution and improved cytogenetic maps of the Y chromosome in cattle, zebu, river buffalo, sheep and goat. Chromosome Res. 2005, 13, 349–355. [Google Scholar] [CrossRef]
- Di Meo, G.P.; Goldammer, T.; Perucatti, A.; Genualdo, V.; Iannuzzi, A.; Incarnato, D.; Rebl, A.; Di Berardino, D.; Iannuzzi, L. Extended cytogenetic maps of sheep chromosome 1 and their cattle and river buffalo homoeologues: Comparison with the OAR1 RH map and human chromosomes 2, 3, 21 and 1q. Cytogenet. Genome Res. 2011, 133, 16–24. [Google Scholar] [CrossRef]
- Perucatti, A.; Genualdo, V.; Iannuzzi, A.; Rebl, A.; Di Berardino, D.; Goldammer, T.; Iannuzzi, L. Advanced comparative cytogenetic analysis of X chromosomes in river buffalo, cattle, sheep, and human. Chromosome Res. 2012, 20, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Proskuryakova, A.A.; Kulemzina, A.I.; Perelman, P.L.; Yudkin, D.V.; Lemskaya, N.A.; Okhlopkov, I.M.; Kirillin, E.V.; Farré, M.; Larkin, D.M.; Roelke-Parker, M.E.; et al. Comparative Chromosome Mapping of Musk Ox and the X Chromosome among Some Bovidae Species. Genes 2019, 10, 857. [Google Scholar] [CrossRef] [Green Version]
- Perucatti, A.; Di Meo, G.P.; Albarella, S.; Ciotola, F.; Incarnato, D.; Jambrenghi, A.C.; Peretti, V.; Vonghia, G.; Iannuzzi, L. Increased frequencies of both chromosome abnormalities and SCEs in two sheep flocks exposed to high dioxin levels during pasturage. Mutagenesis 2006, 21, 67–75. [Google Scholar] [CrossRef]
- Perucatti, A.; Genualdo, V.; Pauciullo, A.; Iorio, C.; Incarnato, D.; Rossetti, C.; Vizzarri, F.; Palazzo, M.; Casamassima, D.; Iannuzzi, L.; et al. Cytogenetic tests reveal no toxicity in lymphocytes of rabbit (Oryctolagus cuniculus, 2n = 44) feed in presence of verbascoside and/or lycopene. Food Chem. Toxicol. 2018, 114, 311–315. [Google Scholar] [CrossRef]
- Di Meo, G.P.; Perucatti, A.; Genualdo, V.; Caputi-Jambrenghi, A.; Rasero, R.; Nebbia, C.; Iannuzzi, L. Chromosome fragility in dairy cows exposed to dioxins and dioxin-like PCBs. Mutagenesis 2011, 26, 269–272. [Google Scholar] [CrossRef]
- Genualdo, V.; Perucatti, A.; Iannuzzi, A.; Di Meo, G.P.; Spagnuolo, S.M.; Caputi-Jambrenghi, A.; Coletta, A.; Vonghia, G.; Iannuzzi, L. Chromosome fragility in river buffalo cows exposed to dioxins. J. Appl. Genet. 2012, 53, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, A.; Perucatti, A.; Genualdo, V.; Rossetti, C.; Iorio, C.; Caputi Jambrenghi, A.; Giannico, F.; Andreassi, M.G.; Iannuzzi, L. Cytogenetic and Genomic Investigations in River Buffaloes Raised in Farms Located in Urban and Rural Areas of Campania Region (Southern-Italy). Ann. Carcinog. 2019, 4, 1020. [Google Scholar]
- Genualdo, V.; Perucatti, A.; Pauciullo, A.; Iannuzzi, A.; Incarnato, D.; Spagnuolo, M.S.; Solinas, N.; Bullitta, S.; Iannuzzi, L. Analysis of chromosome damage by sister chromatid exchange (SCE) and redox homeostasis characterization on sheep flocks from Sardinian pasturelands. Sci. Total Environ. 2015, 527–528, 393–400. [Google Scholar] [CrossRef]
- Iannuzzi, A.; Perucatti, A.; Genualdo, V.; Pauciullo, A.; Melis, R.; Porqueddu, C.; Marchetti, M.; Usai, M.; Iannuzzi, L. Sister chromatid exchange test in river buffalo lymphocytes treated in vitro with furocoumarin extracts. Mutagenesis 2016, 31, 547–551. [Google Scholar] [CrossRef]
- Pawar, K.; Kaul, G. Toxicity of titanium oxide nanoparticles causes functionality and DNA damage in buffalo (Bubalus bubalis) sperm in vitro. Toxicol. Ind. Health 2014, 30, 520–533. [Google Scholar] [CrossRef]
- Ahmed, S.; Othman, O.E. Clastogenic effects of the fasciolicide drug fasinex on river buffalo lymphocyte cultures in vitro. Mutat. Res. 2003, 541, 115–121. [Google Scholar] [CrossRef]
- Albarella, S.; Ciotola, F.; Dario, C.; Iannuzzi, L.; Barbieri, V.; Peretti, V. Chromosome instability in Mediterranean Italian buffaloes affected by limb malformation (transversal hemimelia). Mutagenesis 2009, 24, 471–474. [Google Scholar] [CrossRef] [Green Version]
- Iannuzzi, L. Standard karyotype of the river buffalo (Bubalus bubalis L., 2n = 50). Report of the committee for the standardization of banded karyotypes of the river buffalo. Cytogenet. Cell Genet. 1994, 67, 102–113. [Google Scholar] [CrossRef]
- Ansari, H.A.; Maher, D.W.; Pearce, P.D.; Broad, T.E. Resolving ambiguities in the karyotype of domestic sheep (Ovis aries). II. G-, Q-, and R-banded idiograms, and chromosome-specific molecular markers. Chromosoma 1996, 105, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L.; Gallagher, D.S.; Womack, J.E.; Meo, G.P.; Shelling, C.P.; Groenen, M.A. FISH mapping of the alpha-S2 casein gene on river buffalo and cattle chromosomes identifies a nomenclature discrepancy in the bovine karyotype. Chromosome Res. 1996, 4, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L.; Gallagher, D.S.; Di Meo, G.P.; Diamond, G.; Bevins, C.L.; Womack, J.E. High-resolution FISH mapping of beta-defensin genes to river buffalo and sheep chromosomes suggests a chromosome discrepancy in ffttle standard karyotypes. Cytogenet. Cell Genet. 1996, 75, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Hayes, H.; Di Meo, G.P.; Gautier, M.; Laurent, P.; Eggen, A.; Iannuzzi, L. Localization by FISH of the 31 Texas nomenclature type I markers to both Q- and R-banded bovine chromosomes. Cytogenet. Cell Genet. 2000, 90, 315–320. [Google Scholar] [CrossRef]
- Di Berardino, D.; Di Meo, G.P.; Gallagher, D.S.; Hayes, H.; Iannuzz, L. (Eds.) ISCNDB2000, International System for Chromosome Nomenclature of Domestic Bovids. Cytogenet. Cell Genet. 2001, 92, 283–299. [Google Scholar]
- Iannuzzi, L.; Di Meo, G.P.; Hayes, H.; Perucatti, A.; Incarnato, D.; Gautier, M.; Eggen, A. FISH-mapping of 31 type I loci (Texas markers) to river buffalo chromosomes. Chromosome Res. 2001, 9, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, G.P.; Perucatti, A.; Gautier, M.; Hayes, H.; Incarnato, D.; Eggen, A.; Iannuzzi, L. Chromosome localization of the 31 type I Texas bovine markers in sheep and goat chromosomes by comparative FISH-mapping and R-banding. Anim. Genet. 2003, 34, 294–296. [Google Scholar] [CrossRef] [Green Version]
- Matthee, C.A.; Davis, S.K. Molecular insights into the evolution of the family Bovidae: A nuclear DNA perspective. Mol. Biol. Evol. 2001, 18, 1220–1230. [Google Scholar] [CrossRef] [Green Version]
- MacEachern, S.; McEwan, J.; Goddard, M. Phylogenetic reconstruction and the identification of ancient polymorphism in the Bovini tribe (Bovidae, Bovinae). BMC Genom. 2009, 10, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iannuzzi, L.; King, W.A.; Di Berardino, D. Chromosome evolution in domestic bovids as revealed by chromosome banding and FISH-mapping techniques. Cytogenet. Genome Res. 2009, 126, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L.; Di Berardino, D.; Gustavsson, I.; Ferrara, L.; Di Meo, G.P. Centromeric loss in translocation of centric fusion type in cattle and water buffalo. Hereditas 1987, 106, 73–81. [Google Scholar] [CrossRef]
- Wurster, D.H.; Benirschke, K. Chromosome studies in the superfamily Bovoidea. Chromosoma 1968, 25, 152–171. [Google Scholar] [CrossRef]
- Buckland, R.A.; Evans, H.J. Cytogenetic aspects of phylogeny in the Bovidae. I. G-banding. Cytogenet. Cell Genet. 1978, 21, 42–63. [Google Scholar] [CrossRef]
- Di Berardino, D.; Iannuzzi, L. Chromosome banding homologies in Swamp and Murrah buffalo. J. Hered. 1981, 72, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, D.S., Jr.; Womack, J.E. Chromosome conservation in the Bovidae. J. Hered. 1992, 83, 287–298. [Google Scholar] [CrossRef]
- Hayes, H.; Petit, E.; Dutrillaux, B. Comparison of RBG-banded karyotypes of cattle, sheep, and goats. Cytogenet. Cell Genet. 1991, 57, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Iannuzzi, L.; Di Meo, G.P. Chromosomal evolution in bovids: A comparison of cattle, sheep and goat G- and R-banded chromosomes and cytogenetic divergences among cattle, goat and river buffalo sex chromosomes. Chromosome Res. 1995, 3, 291–299. [Google Scholar] [CrossRef]
- Gallagher, D.S.; Davis, S.K.; De Donato, M.; Burzlaff, J.D.; Womack, J.E.; Taylor, J.F.; Kumamoto, A.T. A molecular cytogenetic analysis of the tribe Bovini (Artiodactyla: Bovidae: Bovinae) with an emphasis on sex chromosome morphology and NOR distribution. Chromosome Res. 1999, 7, 481–492. [Google Scholar] [CrossRef]
- Iannuzzi, L.; Gallagher, D.S.; Ryan, A.M.; Di Meo, G.P.; Womack, J.E. Chromosomal localization of omega and trophoblast interferon genes in cattle and river buffalo by sequential R-banding and fluorescent in situ hybridization. Cytogenet. Cell Genet. 1993, 62, 224–227. [Google Scholar] [CrossRef]
- Iannuzzi, L.; Gallagher, D.S.; Womack, J.E.; Di Meo, G.P.; Skow, L.C.; Ferrara, L.C. Chromosomal localization of the major histocompatibility complex in cattle and river buffalo by fluorescent in situ hybridization. Hereditas 1993, 118, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L.; Di Meo, G.P.; Gallagher, D.S.; Ryan, A.M.; Ferrara, L.; Womack, J.E. Chromosomal localization of omega and trophoblast interferon genes in goat and sheep by fluorescent in situ hybridization. J. Hered. 1993, 84, 301–304. [Google Scholar] [CrossRef]
- Kumar, S.; Nagarajan, M.; Sandhu, J.S.; Kumar, N.; Behl, V.; Nishanth, G. Mitochondrial DNA analyses of Indian water buffalo support a distinct genetic origin of river and swamp buffalo. Anim. Genet. 2007, 38, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Curaudeau, M.; Rozzi, R.; Hassanin, A. The genome of the lowland anoa (Bubalus depressicornis) illuminates the origin of river and swamp buffalo. Mol. Phylogenetics Evol. 2021, 161, 107170. [Google Scholar] [CrossRef]
- Bongso, T.A.; Hilmi, M. Chromosome banding homologies of a tandem fusion in river, swamp and crossbred buffaloes (Bubalus bubalis). Can. J. Genet. Cytol. 1982, 24, 667–673. [Google Scholar] [CrossRef]
- Tanaka, K.; Matsuda, Y.; Masangkay, J.S.; Solis, C.D.; Anunciado, R.V.; Kuro-o, M.; Namikawa, T. Cytogenetic analysis of the tamaraw (Bubalus mindorensis): A comparison of R-banded karyotype and chromosomal distribution of centromeric satellite DNAs, telomeric sequence, and 18S-28S rRNA genes with domestic water buffaloes. J. Hered. 2000, 91, 117–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degrandi, T.M.; Pita, S.; Panzera, Y.; De Oliveira, E.H.; Marques, J.R.; Figueiró, M.R.; Marques, L.C.; Vinadé, L.; Gunski, R.J.; Garnero, A. Karyotypic evolution of ribosomal sites in buffalo subspecies. Genet. Mol. Biol. 2014, 37, 375–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leidenroth, A.; Clapp, J.; Mitchell, L.M.; Coneyworth, D.; Dearden, F.L.; Iannuzzi, L.; Hewitt, J.E. Evolution of DUX gene macrosatellites in placental mammals. Chromosoma 2012, 121, 489–497. [Google Scholar] [CrossRef]
- Shaari, N.; Jaoi-Edward, M.; Loo, S.S.; Salisi, M.S.; Yusoff, R.; Ab Ghani, N.I.; Saad, M.Z.; Ahmad, H. Karyotypic and mtDNA based characterization of Malaysian water buffalo. BMC Genet. 2019, 20, 37. [Google Scholar] [CrossRef] [Green Version]
- Sarataphan, N.; Narongwanichgarn, W.; Maneerat, S. Phylogenetic analysis of a Thai wild water buffalo (Bubalus arnee) through mitochondrial control region. Int. J. Conserv. Sci. 2017, 8, 105–112. [Google Scholar]
- Singh, R.; Lava Kumar, S.; Mishra, S.K.; Gurao, A.; Niranjan, S.K.; Vohra, V.; Dash, S.K.; Rajesh, C.; Kataria, R.S. Mitochondrial sequence-based evolutionary analysis of riverine-swamp hybrid buffaloes of India indicates novel maternal differentiation and domestication patterns. Anim. Genet. 2020, 51, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Zeng, X.L.Q.; Qui, J.; Yang, C. Dairy buffalo breeding in country side of China. Ital. J. Anim. Sci. 2007, 6, 25–29. [Google Scholar] [CrossRef]
- Cruz, L.C. Trends in buffalo production in Asia. Ital. J. Anim. Sci. 2007, 6, 9–24. [Google Scholar] [CrossRef]
- Gustavsson, I. Cytogenetics, distribution and phenotypic effects of a translocation in Swedish cattle. Hereditas 1969, 63, 68–169. [Google Scholar] [CrossRef]
- Gustavsson, I. Chromosome aberrations and their influence on the reproductive performance of domestic animals Y: A review. Z. Tierz. Ziichtungsbiol. 1980, 97, 76–195. [Google Scholar] [CrossRef]
- Dai, K.; Gillies, C.B.; Dollin, A.E.; Hilmi, M. Synaptonemal complex analysis of hybrid and purebred water buffaloes (Bubalus bubalis). Hereditas 1994, 121, 171–184. [Google Scholar]
- Di Dio, C.; Longobardi, V.; Zullo, G.; Parma, P.; Pauciullo, A.; Perucatti, A.; Higgins, J.; Iannuzzi, A. Analysis of meiotic segregation by triple-color fish on both total and motile sperm fractions in a t(1p;18) river buffalo bull. PLoS ONE 2020, 15, e0232592. [Google Scholar] [CrossRef]
- Iannuzzi, L.; Di Meo, G.P.; Perucatti, A.; Schibler, L.; Incarnato, D.; Cribiu, E.P. Comparative FISH mapping in river buffalo and sheep chromosomes: Assignment of forty autosomal type I loci from sixteen human chromosomes. Cytogenet. Cell Genet. 2001, 94, 43–48. [Google Scholar] [CrossRef] [PubMed]
- De Lorenzi, L.; Planas, J.; Rossi, E.; Malagutti, L.; Parma, P. New cryptic karyotypic differences between cattle (Bos taurus) and goat (Capra hircus). Chromosome Res. 2015, 23, 225–235. [Google Scholar] [CrossRef]
- De Lorenzi, L.; Pauciullo, A.; Iannuzzi, A.; Parma, P. Cytogenetic Characterization of a Small Evolutionary Rearrangement Involving Chromosomes BTA21 and OAR18. Cytogenet. Genome Res. 2020, 160, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L.; Di Meo, G.P.; Perucatti, A. Identification of nucleolus organizer chromosomes and frequency of active NORs in river buffalo (Bubalus bubalis L.). Caryologia 1996, 49, 27–34. [Google Scholar] [CrossRef]
- Goldammer, T.; Brunner, R.M.; Rebl, A.; Wu, C.H.; Nomura, K.; Hadfield, T.; Maddox, J.F.; Cockett, N.E. Cytogenetic anchoring of radiation hybrid and virtual maps of sheep chromosome X and comparison of X chromosomes in sheep, cattle, and human. Chromosome Res. 2009, 17, 497–506. [Google Scholar] [CrossRef]
- Iannuzzi, L.; Di Berardino, D. Diagrammatic representation of RBA-banded chromosomes of swamp buffalo (Bubalus bubalis L.) and sex chromosome banding homologies with cattle (Bos taurus L.). Caryologia 1985, 38, 281–295. [Google Scholar] [CrossRef]
- Gustavsson, I.; Rockborn, G. Chromosome abnormality in three cases of lymphatic leukaemia in cattle. Nature 1964, 203, 990. [Google Scholar] [CrossRef]
- Lyon, M.F. Gene action in the X-chromosome of the mouse (Mus musculus L.). Nature 1961, 190, 372–373. [Google Scholar] [CrossRef]
- Burgoyne, P.S.; Ojarikre, O.A.; Turner, J.M. Evidence that postnatal growth retardation in XO mice is due to haploinsufficiency for a non-PAR X gene. Cytogenet. Genome Res. 2002, 99, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, G.P.; Perucatti, A.; Genualdo, V.; Iannuzzi, A.; Sarubbi, F.; Caputi-Jambrenghi, A.; Incarnato, D.; Peretti, V.; Vonghia, G.; Iannuzzi, L. A rare case of centric fission and fusion in a river buffalo (Bubalus bubalis, 2n = 50) cow with reduced fertility. Cytogenet. Genome Res. 2011, 132, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Albarella, S.; Ciotola, F.; Coletta, A.; Genualdo, V.; Iannuzzi, L.; Peretti, V. A new translocation t(1p;18) in an Italian Mediterranean river buffalo (Bubalus bubalis, 2n = 50) bull: Cytogenetic, fertility and inheritance studies. Cytogenet. Genome Res. 2013, 139, 17–21. [Google Scholar] [CrossRef]
- Kotikalapudi, R.; Patel, R.K.; Naik Sugali, N.; Kommuri, M. Structural chromosomal mosaicism due to partial monosomy (3q-) in a Murrah buffalo (Bubalus bubalis) bull. Int. J. Adv. Res. Dev. 2016, 1, 25–27. [Google Scholar]
- Prakash, B.; Balain, D.S.; Lathwal, S.S.; Malik, R.K. Trisomy-X in a sterile river buffalo. Vet. Rec. 1994, 134, 241–242. [Google Scholar] [CrossRef]
- Iannuzzi, L.; Di Meo, G.P.; Perucatti, A.; Incarnato, D.; Di Palo, R.; Zicarelli, L. Reproductive disturbances and sex chromosome abnormalities in two female river buffaloes. Vet. Rec. 2004, 154, 823–824. [Google Scholar] [CrossRef]
- Di Meo, G.P.; Perucatti, A.; Di Palo, R.; Iannuzzi, A.; Ciotola, F.; Peretti, V.; Neglia, G.; Campanile, G.; Zicarelli, L.; Iannuzzi, L. Sex chromosome abnormalities and sterility in river buffalo. Cytogenet. Genome Res. 2008, 120, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Yadav, B.R.; Balakrishnan, C.R. Trisomy of the X chromosome in a Murrah buffalo. Vet. Rec. 1982, 111, 184–185. [Google Scholar] [CrossRef]
- Yadav, B.R.; Kumar, P.; Tomer, O.S.; Kumar, S.; Balain, D.S. Monosomy X and gonadal dysgenesis in a buffalo heifer (Bubalus bubalis). Theriogenology 1990, 34, 99–105. [Google Scholar] [CrossRef]
- Prakash, B.; Balain, D.S.; Lathwal, S.S. A 49, XO sterile murrah buffalo (Bubalus bubalis). Vet. Rec. 1992, 130, 559–560. [Google Scholar] [CrossRef] [PubMed]
- Padula, A.M. The freemartin syndrome: An update. Anim. Reprod. Sci. 2005, 87, 93–109. [Google Scholar] [CrossRef]
- Ruvinsky, A.; Spicer, L.J. Developmental genetics: Sex determination and differentiation. In The Genetics of Cattle; Fries, R., Ruvinsky, A., Eds.; CABI: Wallingford, UK, 1999; pp. 456–461. [Google Scholar]
- Dunn, H.O.; McEntee, K.; Hall, C.E.; Johnson, R.H., Jr.; Stone, W.H. Cytogenetic and reproductive studies of bulls born co-twin with freemartins. J. Reprod. Fertil. 1979, 57, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seguin, B.E.; Zhang, T.Q.; Buoen, L.C.; Weber, A.F.; Ruth, G.R. Cytogenetic survey of Holstein bulls at a commercial artificial insemination company to determine prevalence of bulls with centric fusion and chimeric anomalies. J. Am. Vet. Med Assoc. 2000, 216, 65–67. [Google Scholar] [CrossRef]
- Komisarek, J.; Dorynek, Z. Genetic aspects of twinning in cattle. J. Appl. Genet. 2002, 43, 55–68. [Google Scholar] [PubMed]
- Rocha, L.; Dos Santos, D.; Tonhati, H.; Costa, R.B.; de Camargo, G. Twinning rate in buffaloes: A case report. Reprod. Domest. Anim. 2019, 54, 808–811. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, C.R.; Yadav, B.R.; Sarma, P.A.; Goswami, S.L. Sex chromosome chimaerism in heterosexual Murrah buffalo triplets. Vet. Rec. 1981, 109, 162. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L.; Di Meo, G.P.; Perucatti, A.; Zicarelli, L. Sex chromosome monosomy (2n = 49,X) in a river buffalo (Bubalus bubalis). Vet. Rec. 2000, 147, 690–691. [Google Scholar]
- Iannuzzi, L.; Di Meo, G.P.; Perucatti, A.; Di Palo, R.; Zicarelli, L. 50,XY gonadal dysgenesis (Swyer’s syndrome) in a female river buffalo (Bubalus bubalis). Vet. Rec. 2001, 148, 634–635. [Google Scholar] [CrossRef]
- Iannuzzi, L.; Di Meo, G.P.; Perucatti, A.; Ciotola, F.; Incarnato, D.; Di Palo, R.; Peretti, V.; Campanile, G.; Zicarelli, L. Freemartinism in river buffalo: Clinical and cytogenetic observations. Cytogenet. Genome Res. 2005, 108, 355–358. [Google Scholar] [CrossRef]
- Patel, R.K.; Singh, K.M.; Soni, K.J.; Chauhan, J.B. Novel cytogenetic finding: An unusual X;X-translocation in Mehsana buffalo (Bubalus bubalis). Cytogenet. Genome Res. 2006, 115, 186–188. [Google Scholar] [CrossRef]
- Chauhan, J.B.; Patel, R.K.; Singh, K.M.; Soni, K.J. Impact of a novel cytogenetic finding (unusual X:X translocation) on fertility of a buffalo (Bubalus bubalis). Buffalo Bull. 2009, 28, 151–153. [Google Scholar]
- Iannuzzi, L.; Perucatti, A.; Di Meo, G.P.; Ferrara, L. Sister chromatid exchange in chromosomes of river buffalo (Bubalus bubalis L.). Caryologia 1988, 41, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.; Mahrous, K.; El-Sobhy, H. Cytogenetic study of buffalo under pollution of environmental conditions. Mutat. Res. 1998, 419, 21–26. [Google Scholar] [CrossRef]
- Iannuzzi, L.; Di Meo, G.P.; Perucatti, A.; Ferrara, L. Mitomycin C-induced sister chromatid exchange in X chromosomes of Bovidae. J. Hered. 1990, 81, 78–80. [Google Scholar] [CrossRef] [PubMed]
- Pires, R.M.; Reichert, R.H.; Kasahara, S. Cytogenetics of three breeds of river buffalo (Bubalus bubalis L.), with evidence of a fragile site on the X chromosome. Theriogenology 1998, 49, 529–538. [Google Scholar] [CrossRef]
- Nicodemo, D.; Coppola, G.; Pauciullo, A.; Cosenza, G.; Ramunno, L.; Ciotola, F.; Peretti, V.; Di Meo, G.P.; Iannuzzi, L.; Rubes, J.; et al. Chromosomal expression and localization of aphidicolin-induced fragile sites in the standard karyotype of river buffalo (Bubalus bubalis). Cytogenet. Genome Res. 2008, 120, 178–182. [Google Scholar] [CrossRef]
- Pawar, K.; Kaul, G. Assessment of buffalo (Bubalus bubalis) sperm DNA fragmentation using a sperm chromatin dispersion test. Reprod. Domest. Anim. 2011, 46, 964–969. [Google Scholar] [CrossRef]
- Pinkel, D.; Gray, J.W.; Trask, B.; Van Den Engh, G.; Fuscoe, J.; Van Dekken, H. Cytogenetic analysis by in situ hybridization with fluorescently labeled nucleic acid probes. Cold Spring Harb. Symp. Quant. Biol. 1986, 51 Pt 1, 151–157. [Google Scholar] [CrossRef]
- Trask, B.; Pinkel, D. Fluorescence in situ hybridization with DNA probes. Methods Cell Biol. 1990, 33, 383–400. [Google Scholar]
- Trask, B.J. Fluorescence in situ hybridization: Applications in cytogenetics and gene mapping. Trends Genet. 1991, 7, 149–154. [Google Scholar] [CrossRef]
- Pauciullo, A.; Perucatti, A.; Iannuzzi, A.; Incarnato, D.; Genualdo, V.; Di Berardino, D.; Iannuzzi, L. Development of a sequential multicolor-FISH approach with 13 chromosome-specific painting probes for the rapid identification of river buffalo (Bubalus bubalis, 2n = 50) chromosomes. J. Appl. Genet. 2014, 55, 397–401. [Google Scholar] [CrossRef] [Green Version]
- Hayes, H.; Elduque, C.; Gautier, M.; Schibler, L.; Cribiu, E.; Eggen, A. Mapping of 195 genes in cattle and updated comparative map with man, mouse, rat and pig. Cytogenet. Genome Res. 2003, 102, 16–24. [Google Scholar] [CrossRef]
- Roy, R.; Gautier, M.; Hayes, H.; Laurent, P.; Osta, R.; Zaragoza, P.; Eggen, A.; Rodellar, C. Assignment of the fatty acid synthase (FASN) gene to bovine chromosome 19 (19q22) by in situ hybridization and confirmation by somatic cell hybrid mapping. Cytogenet. Cell Genet. 2001, 93, 141–142. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L.; Gallagher, D.S.; Di Meo, G.P.; Schlapfer, J.; Perucatti, A.; Amarante, M.R.; Incarnato, D.; Davis, S.K.; Taylor, J.F.; Womack, J.E. Twelve loci from HSA10, HSA11 and HSA20 were comparatively FISH-mapped on river buffalo and sheep chromosomes. Cytogenet. Cell Genet. 2001, 93, 124–126. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L.; Di Meo, G.P.; Perucatti, A.; Incarnato, D.; Lopez-Corrales, N.; Smith, J. Chromosomal localization of four HSA2 type I loci in river buffalo (Bubalus bubalis, 2n = 50) chromosomes 2q and 12. Mamm. Genome 2000, 11, 241–242. [Google Scholar] [CrossRef]
- Di Meo, G.P.; Perucatti, A.; Uboldi, C.; Roperto, S.; Incarnato, D.; Roperto, F.; Williams, J.; Eggen, A.; Ferretti, L.; Iannuzzi, L. Comparative mapping of the fragile histidine triad (FHIT) gene in cattle, river buffalo, sheep and goat by FISH and assignment to BTA22 by RH-mapping: A comparison with HSA3. Anim. Genet. 2005, 36, 363–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genualdo, V.; Turri, F.; Pizzi, F.; Castiglioni, B.; Marletta, D.; Iannuzzi, A. Sperm Nuclei Analysis and Nuclear Organization of a Fertile Boar-Pig Hybrid by 2D FISH on Both Total and Motile Sperm Fractions. Animals 2021, 11, 738. [Google Scholar] [CrossRef]
- Gallagher, D.S., Jr.; Basrur, P.K.; Womack, J.E. Identification of an autosome to X chromosome translocation in the domestic cow. J. Hered. 1992, 83, 451–453. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L. Methodologies applied to domestic animal chromosomes. Methods Mol. Biol. 2004, 240, 5–34. [Google Scholar]
- Amaral, M.E.; Grant, J.R.; Riggs, P.K.; Stafuzza, N.B.; Filho, E.A.; Goldammer, T.; Weikard, R.; Brunner, R.M.; Kochan, K.J.; Greco, A.J.; et al. A first generation whole genome RH map of the river buffalo with comparison to domestic cattle. BMC Genom. 2008, 9, 631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ianella, P.; Venancio, L.P.; Stafuzza, N.B.; Miziara, M.N.; Agarwala, R.; Schäffer, A.A.; Riggs, P.K.; Womack, J.E.; Amaral, M.E. First radiation hybrid map of the river buffalo X chromosome (BBUX) and comparison with BTAX. Anim. Genet. 2008, 39, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Goldammer, T.; Weikard, R.; Miziara, M.N.; Brunner, R.M.; Agarwala, R.; Schaffer, A.A.; Womack, J.E.; Amaral, M.E. A radiation hybrid map of river buffalo (Bubalus bubalis) chromosome 7 and comparative mapping to the cattle and human genomes. Cytogenet. Genome Res. 2007, 119, 235–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
African (A) and Asian (B,C) buffaloes, the latter of river (B) and swamp (C) types.

Figure 2.
CBA banding in a river buffalo male metaphase. Note the larger HC block (C-bands) in the X chromosome also showing a proximal positive C-band. The Y chromosome has a telomeric positive C-band, making it easily distinguishable from other autosomes showing all of them to be centromeric C-band positive. Note the small amounts of HC (C-bands) in the biarmed pairs (small arrows).
Figure 2.
CBA banding in a river buffalo male metaphase. Note the larger HC block (C-bands) in the X chromosome also showing a proximal positive C-band. The Y chromosome has a telomeric positive C-band, making it easily distinguishable from other autosomes showing all of them to be centromeric C-band positive. Note the small amounts of HC (C-bands) in the biarmed pairs (small arrows).

Figure 3.
(A) Schematic representation of tandem fusion translocation between river buffalo chromosomes 4p (telomere) and 9 (centromere) in the origin of swamp buffalo chromosome 1. Both NORs available in BBU4p telomeres and SAT-II sequences available at the centromere of BBU9 were lost during the tandem fusion. Only the SAT-I sequences were partially conserved during the fusion. Moreover, homologous cattle chromosomes and relative syntenic groups are reported on the left of BBU4 and BBU9. (B) Details of CBA banding in a male metaphase plate of a hybrid river swamp buffalo. Note a large positive C-band at the centromere of swamp buffalo chromosome 1 (large arrow) and a pale and proximal positive C-band (small arrow), which are probably part of the SAT-I sequences of BBU9 conserved during the fusion. (C) Details of a metaphase plate of hybrid river–swamp buffaloes showing the result of the FISH mapping, with the marker INRAb_94D present both at the telomeres of BBU4 and the fusion point of swamp buffalo chromosome 1 (arrows). (D) Details of a metaphase plate of hybrid river–swamp buffalo showing the result of the FISH mapping, with the marker DUXC present at both the pericentromeric region of BBU9 and the fusion point of swamp buffalo chromosome 1 (arrow). (E) River–swamp buffalo hybrid metaphase showing the result of FISH mapping with a PNA telomeric probe. Clear hybridization signals were observed only at the telomeres of all chromosomes, and absent at the fusion point in swamp buffalo chromosome 1 (arrow).
Figure 3.
(A) Schematic representation of tandem fusion translocation between river buffalo chromosomes 4p (telomere) and 9 (centromere) in the origin of swamp buffalo chromosome 1. Both NORs available in BBU4p telomeres and SAT-II sequences available at the centromere of BBU9 were lost during the tandem fusion. Only the SAT-I sequences were partially conserved during the fusion. Moreover, homologous cattle chromosomes and relative syntenic groups are reported on the left of BBU4 and BBU9. (B) Details of CBA banding in a male metaphase plate of a hybrid river swamp buffalo. Note a large positive C-band at the centromere of swamp buffalo chromosome 1 (large arrow) and a pale and proximal positive C-band (small arrow), which are probably part of the SAT-I sequences of BBU9 conserved during the fusion. (C) Details of a metaphase plate of hybrid river–swamp buffaloes showing the result of the FISH mapping, with the marker INRAb_94D present both at the telomeres of BBU4 and the fusion point of swamp buffalo chromosome 1 (arrows). (D) Details of a metaphase plate of hybrid river–swamp buffalo showing the result of the FISH mapping, with the marker DUXC present at both the pericentromeric region of BBU9 and the fusion point of swamp buffalo chromosome 1 (arrow). (E) River–swamp buffalo hybrid metaphase showing the result of FISH mapping with a PNA telomeric probe. Clear hybridization signals were observed only at the telomeres of all chromosomes, and absent at the fusion point in swamp buffalo chromosome 1 (arrow).

Figure 4.
RBA-banding in a metaphase plate of a hybrid river–swamp buffalo (2n = 49). BBU4, BBU9, swamp chromosome 1, X, and Y chromosomes are indicated (arrows).
Figure 4.
RBA-banding in a metaphase plate of a hybrid river–swamp buffalo (2n = 49). BBU4, BBU9, swamp chromosome 1, X, and Y chromosomes are indicated (arrows).

Figure 5.
RBA-banded river buffalo karyotype is arranged according to the standard river buffalo karyotype [47]. The numbers between parentheses are referred to as the cattle homologous chromosomes, according to ISCNDB2000, 2001.
Figure 5.
RBA-banded river buffalo karyotype is arranged according to the standard river buffalo karyotype [47]. The numbers between parentheses are referred to as the cattle homologous chromosomes, according to ISCNDB2000, 2001.

Figure 6.
The interphase nucleus of a river buffalo female affected by X-chromosome monosomy, and showing only one FITC signal (arrow) after FISH mapping with a specific marker (PGK) of the X chromosome.
Figure 6.
The interphase nucleus of a river buffalo female affected by X-chromosome monosomy, and showing only one FITC signal (arrow) after FISH mapping with a specific marker (PGK) of the X chromosome.

Table 1.
Diploid number (2n), fundamental number (FN), and cattle homologous chromosomes (ancestral bovid karyotype) involved in the autosomal biarmed pairs of both African (Syncerus) and Asiatic (Bubalus) buffaloes according to [47,52,62,65].
| Buffalo | Genus | Species | Sub-Species | 2n/FN | Biarmed Pairs (Cattle Homologous Chromosomes Are Reported between Parentheses) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| African | Syncerus | caffer | caffer | 52/60 | 1(1;13) | 2(2;3) | 3(5;20) | 4(11;29) | ||
| Syncerus | caffer | nanus | 54/60 | 1(1;13) | 2(2;3) | 3(5;20) | ||||
| Asiatic | Bubalus | bubalis | bubalis | 50/60 | 1(1;27) | 2(2;23) | 3(8;19) | 4(5;28) | 5(16;29) | |
| Bubalus | bubalis | carabenensis | 48/58 | 1(5;28;7) * | 2(1;27) | 3(2;23) | 4(8;19) | 5(16;29) | ||
| Bubalus | mindorensis | 46/58 | 1(5;28;7) * | 2(1;27) | 3(2;23) | 4(8;19) | 5(4;14) | 6(16;29) | ||
| Bubalus | depressicornis | Depressicornis | 48/60 | 1(1;27) | 2(2;23) | 3(8;19) | 4(5;28) | 5(11;20) | 6(17;15) | |
| Bubalus | depressicornis | quarlesi | 48/60 | 1(1;27) | 2(2;23) | 3(8;19) | 4(5;28) | 5(11;20) | 6(17;15) | |
* Centric fusion + tandem fusion.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Iannuzzi, A.; Parma, P.; Iannuzzi, L. The Cytogenetics of the Water Buffalo: A Review. Animals 2021, 11, 3109. https://doi.org/10.3390/ani11113109
AMA Style
Iannuzzi A, Parma P, Iannuzzi L. The Cytogenetics of the Water Buffalo: A Review. Animals. 2021; 11(11):3109. https://doi.org/10.3390/ani11113109
Chicago/Turabian StyleIannuzzi, Alessandra, Pietro Parma, and Leopoldo Iannuzzi. 2021. "The Cytogenetics of the Water Buffalo: A Review" Animals 11, no. 11: 3109. https://doi.org/10.3390/ani11113109
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.