
Investigating the Short-Term Effects of Cold Stress on Metabolite Responses and Metabolic Pathways in Inner-Mongolia Sanhe Cattle

 , ,
, ,  ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
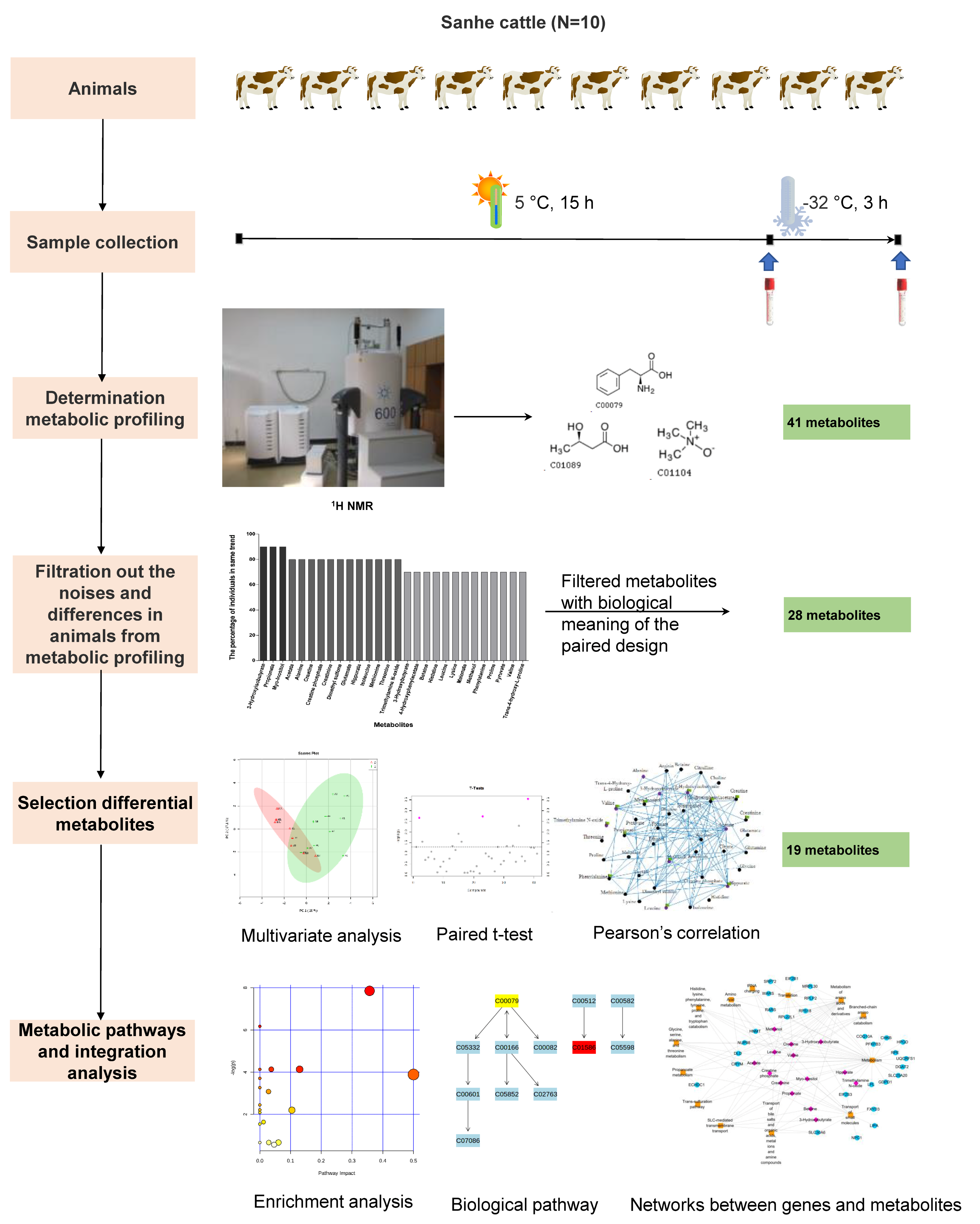
2.1. Animals and Sample Collection
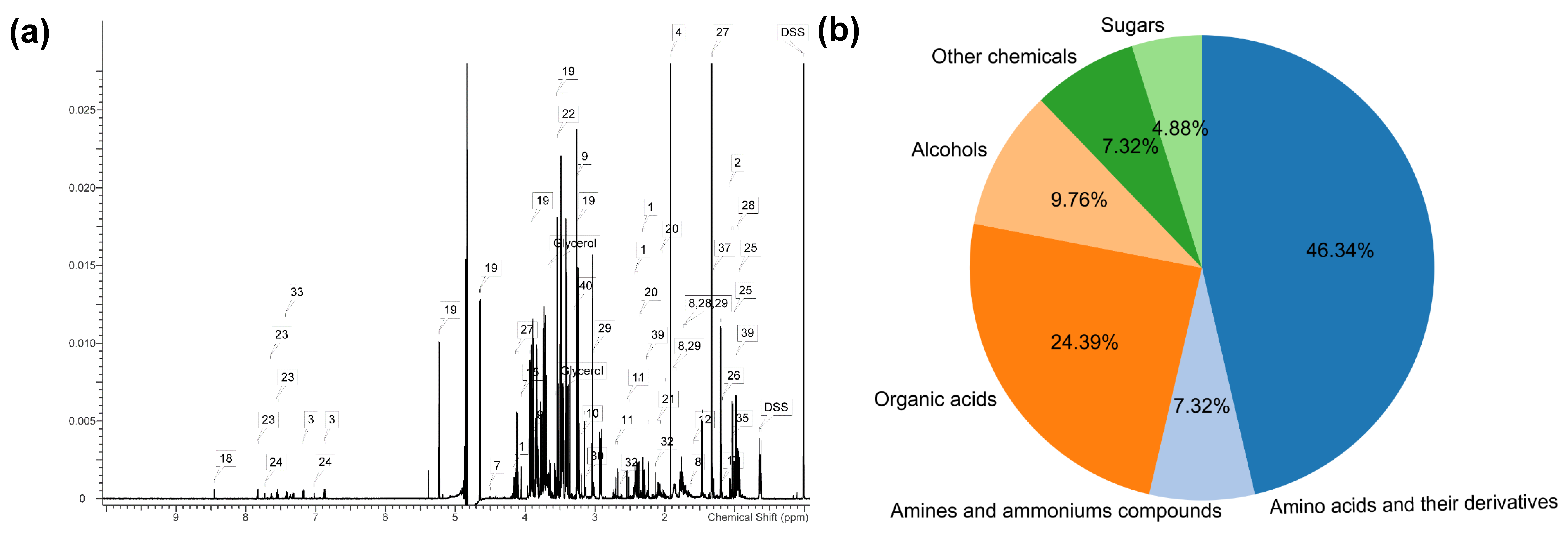
2.2. 1H Nuclear Magnetic Resonance (1H NMR) Analyses
2.3. Metabolome Analysis
2.4. Integration Analysis of Transcriptome and Metabolome Datasets
3. Results
3.1. The Serum Metabolome in Inner-Mongolia Sanhe Cattle
3.2. Pre-Selection of Candidate Metabolites Related to Severe Cold Stress in Sanhe Cattle
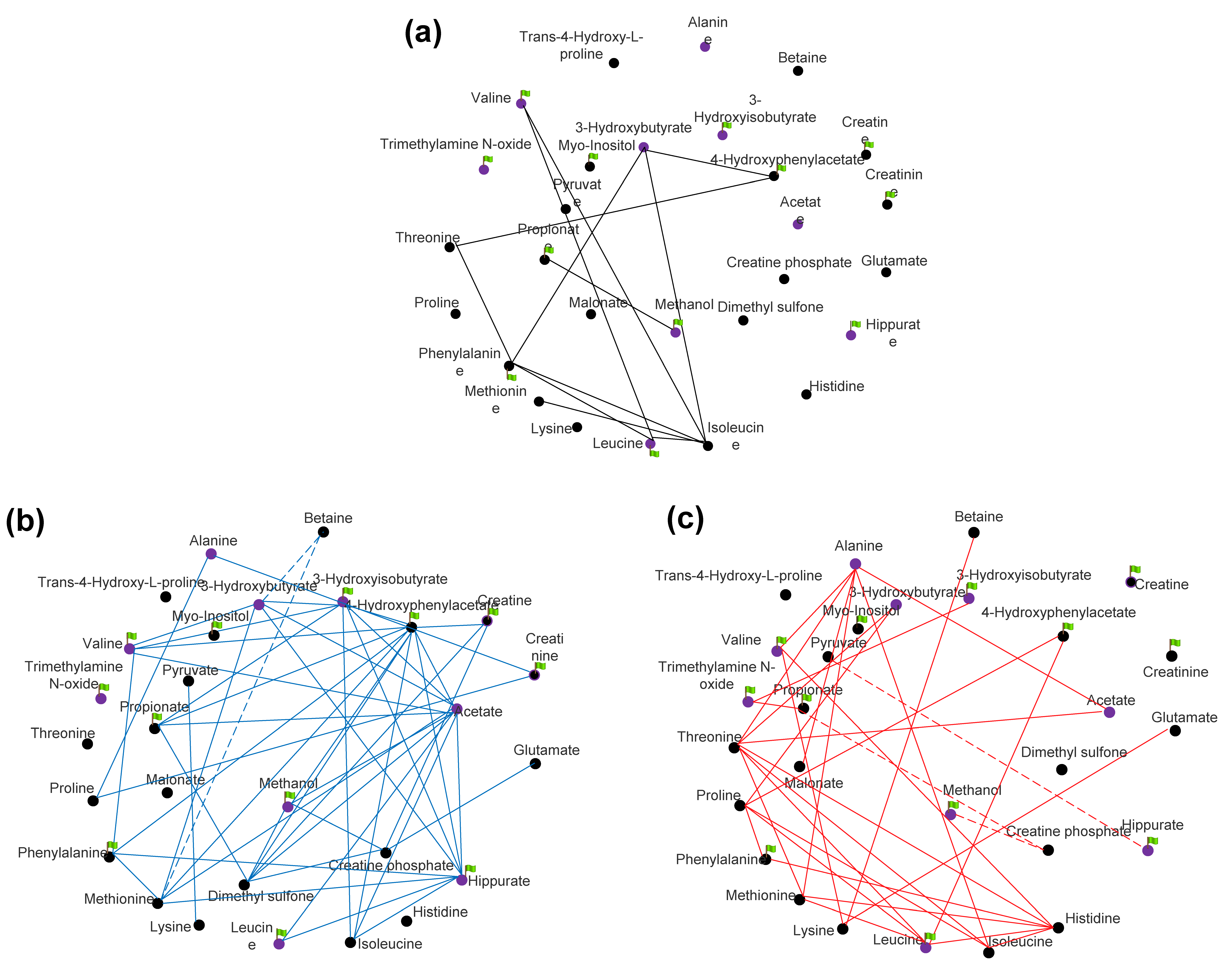
3.3. Metabolite Changes in the Serum of Sanhe Cattle after Severe Cold Stress
3.4. Changes of Correlation between Candidate Metabolites under Severe Cold Stress
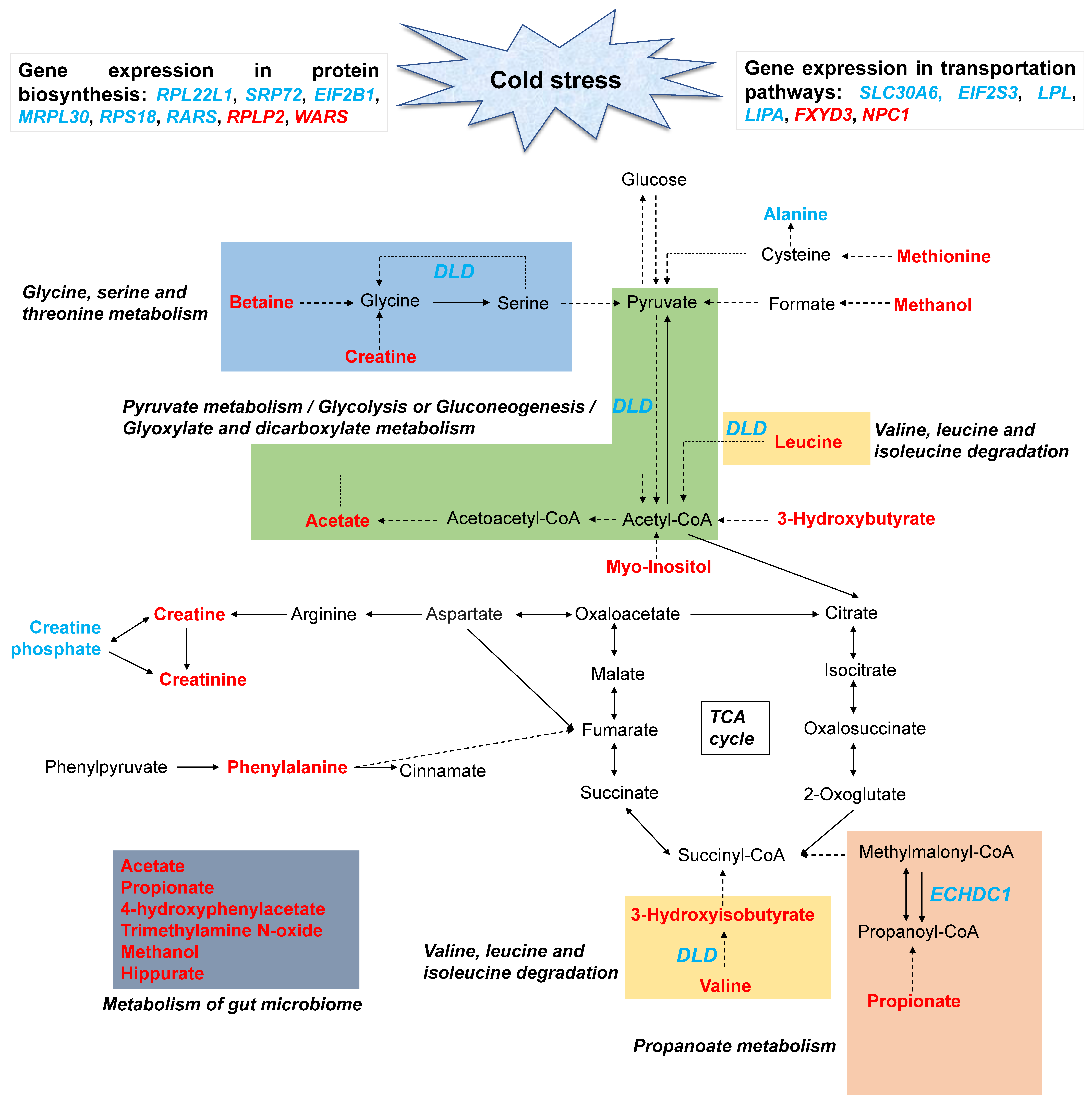
3.5. Key Metabolic Pathways Involved in Severe Cold Stress
3.6. Significant Biological Processes Related to Metabolites’ Regulation in Severe Cold Stress
4. Discussion
4.1. The Pre-Filtration for the Paired Metabolome Data
4.2. The Metabolic Pathways’ Response to Severe Cold Stress in Sanhe Cattle
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kang, H.J.; Lee, I.K.; Piao, M.Y.; Gu, M.J.; Yun, C.H.; Kim, H.J.; Kim, K.H.; Baik, M. Effects of Ambient Temperature on Growth Performance, Blood Metabolites, and Immune Cell Populations in Korean Cattle Steers. Asian-Australas J. Anim. Sci. 2016, 29, 436–443. [Google Scholar] [CrossRef] [Green Version]
- Young, B.A. Cold stress as it affects animal production. J. Anim. Sci. 1981, 52, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Uematsu, M.; Kitahara, G.; Osawa, T. Reproductive performance of Japanese Black cattle: Association with herd size, season, and parity in commercial cow-calf operations. Theriogenology 2016, 86, 2156–2161. [Google Scholar] [CrossRef] [PubMed]
- Hemsworth, P.H.; Barnett, J.L.; Beveridge, L.; Matthews, L.R. The welfare of extensively managed dairy cattle: A review. Appl. Anim. Behav. Sci. 1995, 42, 161–182. [Google Scholar] [CrossRef]
- Guo, J.; Yang, H.; Wang, Z. Research progress on effects of cold stress on function of immune system in cattle and poultry. J. Environ. Health 2010, 27, 649–651. [Google Scholar]
- Nguyen, P.; Greene, E.; Donoghue, A.; Huff, G.; Clark, F.; Dridi, S. A new insight into cold stress in poultry production. Adv. Food Technol. Nutr. Sci. Open J. 2016, 2, 1–2. [Google Scholar] [CrossRef]
- Liu, G.L.; Wang, J.Q.; Liu, W.Z.; Zhang, W.F.; Fu, D.P.; Wang, J.P. Study on heat stress and cold stress of dairy cows in different regions of China. China Dairy Cattle 2009, 8, 66–69. [Google Scholar]
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; Church, J.A.; Clarke, L.; Dahe, Q.; LDasgupta, P.; et al. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Swtzerland, 2015.
- Abbas, Z.; Hu, L.; Fang, H.; Sammad, A.; Kang, L.; Brito, L.F.; Xu, Q.; Wang, Y. Association Analysis of Polymorphisms in the 5’ Flanking Region of the HSP70 Gene with Blood Biochemical Parameters of Lactating Holstein Cows under Heat and Cold Stress. Animals 2020, 10, 2016. [Google Scholar] [CrossRef]
- Fang, H.; Kang, L.; Abbas, Z.; Hu, L.; Chen, Y.; Tan, X.; Wang, Y.; Xu, Q. Identification of key Genes and Pathways Associated With Thermal Stress in Peripheral Blood Mononuclear Cells of Holstein Dairy Cattle. Front. Genet. 2021, 12, 662080. [Google Scholar] [CrossRef]
- Xu, Q.; Wang, Y.C.; Liu, R.; Brito, L.F.; Kang, L.; Yu, Y.; Wang, D.S.; Wu, H.J.; Liu, A. Differential gene expression in the peripheral blood of Chinese Sanhe cattle exposed to severe cold stress. Genet. Mol. Res. 2017, 16. [Google Scholar] [CrossRef]
- Wang, X.; Xu, S.; Gao, X.; Ren, H.; Chen, J. Genetic Polymorphism of TLR4 Gene and Correlation with Mastitis in Cattle. J. Genet. Genom. 2007, 34, 406–412. [Google Scholar] [CrossRef]
- Wu, H.J.; Ma, X.L.; Liu, A.R.; Liu, Y.S.; Zhao, J.H.; Wang, D.S.; Liu, H.Z. Breeding history and current improvements of Sanhe Cattle in Inner Mongolia. China Cattle Sci. 2012, 38, 48–52. [Google Scholar]
- Hu, L.; Ma, Y.; Liu, L.; Kang, L.; Brito, L.F.; Wang, D.; Wu, H.; Liu, A.; Wang, Y.; Xu, Q. Detection of functional polymorphisms in the hsp70 gene and association with cold stress response in Inner-Mongolia Sanhe cattle. Cell Stress Chaperones 2019, 24, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O. Metabolomics—The link between genotypes and phenotypes. Plant Mol. Biol. 2002, 155–1711. [Google Scholar] [CrossRef]
- Bathe, O.F.; Shaykhutdinov, R.; Kopciuk, K.; Weljie, A.M.; McKay, A.; Sutherland, F.R.; Dixon, E.; Dunse, N.; Sotiropoulos, D.; Vogel, H.J. Feasibility of identifying pancreatic cancer based on serum metabolomics. Cancer Epidemiol. Biomark. Prev. 2011, 20, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Tiziani, S.; Kang, Y.; Choi, J.S.; Roberts, W.; Paternostro, G. Metabolomic high-content nuclear magnetic resonance-based drug screening of a kinase inhibitor library. Nat. Commun. 2011, 2, 545. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S. Metabolomics: Applications to food science and nutrition research. Trends Food Sci. Technol. 2008, 19, 482–493. [Google Scholar] [CrossRef]
- Xiao, Z.; Ge, C.; Zhou, G.; Zhang, W.; Liao, G. 1H NMR-based metabolic characterization of Chinese Wuding chicken meat. Food Chem. 2018, 274, 574–582. [Google Scholar] [CrossRef]
- Alreshidi, M.M.; Dunstan, R.H.; Macdonald, M.M.; Smith, N.D.; Gottfries, J.; Roberts, T.K. Metabolomic and proteomic responses of Staphylococcus aureus to prolonged cold stress. J. Proteom. 2015, 121, 44–55. [Google Scholar] [CrossRef]
- Colinet, H.; Renault, D.; Javal, M.; Berková, P.; Šimek, P.; Koštál, V. Uncovering the benefits of fluctuating thermal regimes on cold tolerance of drosophila flies by combined metabolomic and lipidomic approach. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2016, 1861, 1736–1745. [Google Scholar] [CrossRef]
- MacMillan, H.A.; Knee, J.M.; Dennis, A.B.; Udaka, H.; Marshall, K.E.; Merritt, T.J.S.; Sinclair, B.J. Cold acclimation wholly reorganizes the Drosophila melanogaster transcriptome and metabolome. Sci. Rep. 2016, 6, 28999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhao, T.; Qiu, Y.; Su, M.; Jiang, T.; Zhou, M.; Zhao, A.; Jia, W. Metabonomics Approach to Understanding Acute and Chronic Stress in Rat Models. J. Proteome Res. 2009, 8, 2511–2518. [Google Scholar] [CrossRef]
- Gandhi, S.; Devi, M.M.; Pal, S.; Tripathi, R.P.; Khushu, S. Metabolic regulatory variations in rats due to acute cold stress & Tinospora Cordifolia intervention: High resolution 1H NMR approach. Metabolomics 2012, 8, 444–453. [Google Scholar]
- Beckonert, O.; Keun, H.C.; Ebbels, T.M.D.; Bundy, J.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. Metabolic profiling, metabolomic and metabonomic procedures for NMR spectroscopy of urine, plasma, serum and tissue extracts. Nat. Protoc. 2007, 2, 2692. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Hu, R.; Wang, Z.; Peng, Q.; Dong, X.; Zhang, X.; Zou, H.; Pu, Q.; Xue, B.; Wang, L. Metabolomics Profiling of Serum and Urine in Three Beef Cattle Breeds Revealed Different Levels of Tolerance to Heat Stress. J. Agric. Food Chem. 2018, 66, 6926–6935. [Google Scholar] [CrossRef]
- Harrell, F.; Dupont, C. Hmisc: Harrell Miscellaneous. R Package Version 3.3.2. Available online: https://CRAN.R-project.org/package=Hmisc (accessed on 20 December 2020).
- Kamburov, A.; Cavill, R.; Ebbels, T.M.; Herwig, R.; Keun, H.C. Integrated pathway-level analysis of transcriptomics and metabolomics data with IMPaLA. Bioinformatics 2011, 27, 2917–2918. [Google Scholar] [CrossRef] [PubMed]
- Kohl, M.; Wiese, S.; Warscheid, B. Cytoscape: Software for visualization and analysis of biological networks. Methods Mol. Biol. 2011, 696, 291–303. [Google Scholar] [PubMed]
- Li, W.; Liu, R.; Ma, Y.; Li, J.; Wu, H.; Ai, L.; Yu, Y.; Xu, Q.; Wang, Y. Effects of Severe Cold Stress on Blood Biochemical Parameters and Related Gene Expression in Sanhe Cattle. Chin. J. Anim. Vet. Sci. 2015, 46, 8. [Google Scholar]
- Freitas, P.H.F.; Wang, Y.; Yan, P.; Oliveira, H.R.; Schenkel, F.S.; Zhang, Y.; Xu, Q.; Brito, L.F. Genetic Diversity and Signatures of Selection for Thermal Stress in Cattle and Other Two Bos Species Adapted to Divergent Climatic Conditions. Front. Genet. 2021, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Regier, N.G.; Naik, A.D.; Mulligan, E.A.; Nasreddine, Z.S.; Driver, J.A.; Sada, Y.H.F.; Moye, J. Cancer-related cognitive impairment and associated factors in a sample of older male oral-digestive cancer survivors. Psychooncology 2019, 28, 1551–1558. [Google Scholar] [CrossRef]
- Ke, C.; Qun, L.V.W.; Ching, L.S.S. A Tablet-Based Volunteer-Mediated Intervention for Cognitively Impaired Older People: A Pretest–Posttest. Res. Soc. Work Pract. 2020, 30, 288–297. [Google Scholar]
- Ferreira, R.A.; Teixeira, A.O.; Oliveira, R.F.; Cantarelli, V.S.; Gionbelli, M.P.; Abreu, M.L.T.; Moreira, R.H.R. Effect of air temperature physiology and productive performance of pigs during growing and finishing phases. S. Afr. J. Anim. Sci. 2018, 48, 627–635. [Google Scholar]
- Pate, R.T.; Luchini, D.; Murphy, M.R.; Cardoso, F.C. Effects of rumen-protected methionine on lactation performance and physiological variables during a heat stress challenge in lactating Holstein cows. J. Dairy Sci. 2020, 103, 2800–2813. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Wang, W.; Zheng, N.; Cheng, J.; Li, S.; Zhang, Y.; Wang, J. Identification of diagnostic biomarkers and metabolic pathway shifts of heat-stressed lactating dairy cows. J. Proteom. 2015, 125, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Gowda, H.; Ivanisevic, J.; Johnson, C.H.; Kurczy, M.E.; Benton, H.P.; Rinehart, D.; Nguyen, T.; Ray, J.; Kuehl, J.; Arevalo, B.; et al. Interactive XCMS Online: Simplifying Advanced Metabolomic Data Processing and Subsequent Statistical Analyses. Anal. Chem. 2014, 86, 6931–6939. [Google Scholar] [CrossRef]
- Southam, A.D.; Easton, J.M.; Stentiford, G.D.; Ludwig, C.; Arvanitis, T.N.; Viant, M.R. Metabolic Changes in Flatfish Hepatic Tumours Revealed by NMR-Based Metabolomics and Metabolic Correlation Networks. J. Proteome Res. 2008, 7, 5277–5285. [Google Scholar] [CrossRef]
- Pohjanen, E.; Thysell, E.; Jonsson, P.; Eklund, C.; Silfver, A.; Carlsson, I.B.; Lundgren, K.; Moritz, T.; Svensson, M.B.; Antti, H. A Multivariate Screening Strategy for Investigating Metabolic Effects of Strenuous Physical Exercise in Human Serum. J. Proteome Res. 2007, 6, 2113–2120. [Google Scholar] [CrossRef]
- Westerhuis, J.A.; van Velzen, E.J.; Hoefsloot, H.C.; Smilde, A.K. Multivariate paired data analysis: Multilevel PLSDA versus OPLSDA. Metabolomics 2010, 6, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Thiébaut, R. A novel approach for biomarker selection and the integration of repeated measures experiments from two assays. BMC Bioinform. 2012, 13, 325. [Google Scholar]
- Du, H.; Li, S.; Zhang, Y.; Guo, H.; Wu, L.; Liu, H.; Manyande, A.; Xu, F.; Wang, J. NMR Based Metabolomics Comparison of Different Blood Sampling Techniques in Awake and Anesthetized Rats. Molecules 2019, 24, 2542. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Su, M.; Qiu, Y.; Ni, Y.; Zhao, T.; Zhou, M.; Zhao, A.; Yang, S.; Zhao, L.; Jia, W. Metabolic Regulatory Network Alterations in Response to Acute Cold Stress and Ginsenoside Intervention. J. Proteome Res. 2007, 6, 3449–3455. [Google Scholar] [CrossRef]
- Jiao, S.; Nie, M.; Song, H.; Xu, D.; You, F. Physiological responses to cold and starvation stresses in the liver of yellow drum (Nibea albiflora) revealed by LC-MS metabolomics. Sci. Total Environ. 2020, 715, 136940. [Google Scholar] [CrossRef]
- Coloma-Garcia, W.; Mehaba, N.; Such, X.; Caja, G.; Salama, A.A.K. Effects of Cold Exposure on Some Physiological, Productive, and Metabolic Variables in Lactating Dairy Goats. Animals 2020, 10, 2383. [Google Scholar] [CrossRef] [PubMed]
- Marosi, K.; Kim, S.W.; Moehl, K.; Scheibye-Knudsen, M.; Cheng, A.; Cutler, R.; Camandola, S.; Mattson, M.P. 3-Hydroxybutyrate regulates energy metabolism and induces BDNF expression in cerebral cortical neurons. J. Neurochem. 2016, 139, 769–781. [Google Scholar] [CrossRef]
- Tian, H.; Zheng, N.; Wang, W.; Cheng, J.; Li, S.; Zhang, Y.; Wang, J. Integrated Metabolomics Study of the Milk of Heat-stressed Lactating Dairy Cows. Sci. Rep. 2016, 6, 24208. [Google Scholar] [CrossRef]
- Lu, Z.; He, X.; Ma, B.; Zhang, L.; Li, J.; Jiang, Y.; Zhou, G.; Gao, F. Serum metabolomics study of nutrient metabolic variations in chronic heat-stressed broilers. Br. J. Nutr. 2018, 119, 771–781. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Wang, C.; Hao, Y.; Gu, X.; Wang, H. Chronic Heat Stress Induces Acute Phase Responses and Serum Metabolome Changes in Finishing Pigs. Animals 2019, 9, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malyszko, J.; Urano, T.; Yan, D.; Serizawa, K.; Kozima, Y.; Takada, Y.; Takada, A. Foot shock-induced changes in blood and brain serotonin and related substances in rats. Jpn. J. Physiol. 1994, 44, 35–47. [Google Scholar] [CrossRef] [Green Version]
- Teague, C.R.; Dhabhar, F.S.; Barton, R.H.; Beckwith-Hall, B.; Powell, J.; Cobain, M.; Singer, B.; McEwen, B.S.; Lindon, J.C.; Nicholson, J.K.; et al. Metabonomic Studies on the Physiological Effects of Acute and Chronic Psychological Stress in Sprague−Dawley Rats. J. Proteome Res. 2007, 6, 2080–2093. [Google Scholar] [CrossRef]
- Gu, Z.; Li, L.; Tang, S.; Liu, C.; Fu, X.; Shi, Z.; Mao, H. Metabolomics Reveals that Crossbred Dairy Buffaloes Are More Thermotolerant than Holstein Cows under Chronic Heat Stress. J. Agric. Food Chem. 2018, 66, 12889–12897. [Google Scholar] [CrossRef]
- Wiltrout, D.W.; Satter, L.D. Contribution of Propionate to Glucose Synthesis in the Lactating and Nonlactating Cow1. J. Dairy Sci. 1972, 55, 307–317. [Google Scholar] [CrossRef]
- Abbas, Z.; Sammad, A.; Hu, L.; Fang, H.; Xu, Q.; Wang, Y. Glucose Metabolism and Dynamics of Facilitative Glucose Transporters (GLUTs) under the Influence of Heat Stress in Dairy Cattle. Metabolites 2020, 10, 312. [Google Scholar] [CrossRef]
- Dumas, M.-E.; Barton, R.H.; Toye, A.; Cloarec, O.; Blancher, C.; Rothwell, A.; Fearnside, J.; Tatoud, R.; Blanc, V.; Lindon, J.C.; et al. Metabolic profiling reveals a contribution of gut microbiota to fatty liver phenotype in insulin-resistant mice. Proc. Natl. Acad. Sci. USA 2006, 103, 12511. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Metabolite | Metabolic Pathway | No. | Metabolite | Metabolic Pathway |
|---|---|---|---|---|---|
| 1 | 3-Hydrxyisobutyrate | Amino acid | 22 | Trans-4-Hydroxy-l-proline | Amino acid |
| 2 | 4-Hydroxyphenylacetate | Amino acid | 23 | Betaine | Betaine metabolism |
| 3 | Alanine | Amino acid | 24 | Acetate | Carbohydrate |
| 4 | Arginine | Amino acid | 25 | Arabinose | Carbohydrate |
| 5 | Citrulline | Amino acid | 26 | Citrate | Carbohydrate |
| 6 | Creatine | Amino acid | 27 | Ethanol | Carbohydrate |
| 7 | Creatine phosphate | Amino acid | 28 | Formate | Carbohydrate |
| 8 | Creatinine | Amino acid | 29 | Glucose | Carbohydrate |
| 9 | Glutamate | Amino acid | 30 | Isopropanol | Carbohydrate |
| 10 | Glutamine | Amino acid | 31 | Lactate | Carbohydrate |
| 11 | Glycine | Amino acid | 32 | Methanol | Carbohydrate |
| 12 | Hippurate | Amino acid | 33 | Propionate | Carbohydrate |
| 13 | Histidine | Amino acid | 34 | Pyruvate | Carbohydrate |
| 14 | Isoleucine | Amino acid | 35 | Myo-Inositol | Carbohydrate |
| 15 | Leucine | Amino acid | 36 | Trimethylamine N-oxide | Gut microbiome-derived metabolism |
| 16 | Lysine | Amino acid | 37 | 3-Hydroxybutyrate | Lipid |
| 17 | Methionine | Amino acid | 38 | Acetone | Lipid |
| 18 | Phenylalanine | Amino acid | 39 | Choline | Lipid |
| 19 | Proline | Amino acid | 40 | Malonate | Lipid |
| 20 | Threonine | Amino acid | 41 | Dimethyl sulfone | Sulfur metabolism |
| 21 | Valine | Amino acid |
| Metabolite | VIP 1 | p-Value 2 | Correlation Coefficient Difference 3 | Regulation Status 4 |
|---|---|---|---|---|
| 3-Hydroxybutyrate | 2.44 | 0.072 | >1 | up |
| Trimethylamine N-oxide | 2.07 | 0.001 | NA | up |
| Methanol | 1.43 | 0.050 | >1 | up |
| Hippurate | 1.34 | 0.002 | >1 | up |
| Acetate | 1.33 | 0.400 | NA | up |
| Valine | 1.29 | 0.050 | >1 | up |
| 3-Hydroxyisobutyrate | 1.22 | 0.002 | >1 | up |
| Alanine | 1.08 | NA | NA | down |
| Leucine | 1.06 | 0.050 | NA | up |
| 4-Hydroxyphenylacetate | 0.84 | 0.027 | >1 | up |
| Betaine | 0.06 | 0.830 | >1 | up |
| Creatine | 0.92 | 0.049 | >1 | up |
| Creatine phosphate | 0.44 | 0.086 | >1 | down |
| Creatinine | 0.85 | 0.010 | NA | up |
| Dimethyl sulfone | 0.31 | 0.170 | >1 | up |
| Methionine | 0.43 | NA | >1 | up |
| Myo-Inositol | 0.62 | 0.050 | NA | up |
| Phenylalanine | 0.81 | 0.050 | NA | up |
| Propionate | 0.79 | 0.011 | >1 | up |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, L.; Brito, L.F.; Abbas, Z.; Sammad, A.; Kang, L.; Wang, D.; Wu, H.; Liu, A.; Qi, G.; Zhao, M.; et al. Investigating the Short-Term Effects of Cold Stress on Metabolite Responses and Metabolic Pathways in Inner-Mongolia Sanhe Cattle. Animals 2021, 11, 2493. https://doi.org/10.3390/ani11092493
Hu L, Brito LF, Abbas Z, Sammad A, Kang L, Wang D, Wu H, Liu A, Qi G, Zhao M, et al. Investigating the Short-Term Effects of Cold Stress on Metabolite Responses and Metabolic Pathways in Inner-Mongolia Sanhe Cattle. Animals. 2021; 11(9):2493. https://doi.org/10.3390/ani11092493
Chicago/Turabian StyleHu, Lirong, Luiz F. Brito, Zaheer Abbas, Abdul Sammad, Ling Kang, Dongsheng Wang, Hongjun Wu, Airong Liu, Guiqiang Qi, Man Zhao, and et al. 2021. "Investigating the Short-Term Effects of Cold Stress on Metabolite Responses and Metabolic Pathways in Inner-Mongolia Sanhe Cattle" Animals 11, no. 9: 2493. https://doi.org/10.3390/ani11092493