1. Introduction
Meat and meat products derived from Iberian pigs (autochthonous breed pig reared in the Southwest of Spain) are highly valued and accepted products at national and international markets [
1], mainly due to their high quality and organoleptic characteristics.
It is well-known that both the production system and breeding have an influence on the quality of meat and meat products. The traditional Iberian pigs finishing system is linked to the
dehesas (
Quercus sp. open woodlands that are found in the Southwest of the Iberian Peninsula), in which the animals are fed exclusively on acorns and grass. Animals finished in
dehesas result in high-quality products. However, the geographical limitations of
dehesas, as well as the natural resources seasonality (November–March), have led to a diversification in the way Iberian pigs are produced in order to face the growing demand of Iberian products [
2]. Thus, the intensification of the management system and a feeding program based on commercial feedstuffs has become an alternative to the traditional
Montanera production system. In addition, the requirements to keep production costs low have led to the spread of Iberian × Duroc (IB × D) pig breeding programmes across farms [
3], given their better productive parameters compared to purebred animals [
4]. This results in crossbred animals being able to reach weights similar to those of purebred Iberian pigs at a younger age and, therefore, reduces the production cycle. Nevertheless, animal age at slaughter is regulated by the current legislative framework—the Spanish Iberian Quality Standard (SIQS) [
5]—that is linked to the production system, despite the growth rate of the animals used (Iberian purebred or crossed with Duroc). Specifically, for animals being reared in the
Montanera system, the minimum age at which they must be slaughtered is 14 months, while, for animals in intensive systems, this is 10 months. Therefore, animal age at slaughter may generate disagreement between the requirements of the current regulatory framework and the interests of farmers, who typically use this breed to increase productivity and reduce costs [
3].
On the other hand, the shortening of the production period may bring about alterations in the quality of the meat [
6,
7], which could be associated with
post-mortem changes in the conversion of muscle to meat and fibre-type composition [
8,
9,
10]. Previous studies have concluded age-related changes on approximate compositions. More in detail, Lorenzo, Fernández, Iglesias, Carril, Rodríguez and Franco [
11] observed higher values of dry matter with animal age in meat from Celtic breed pigs, whilst Bosch, Tor, Reixach and Estany [
12] observed that intramuscular fat increased with animal age in fresh loins belonging to Duroc breed pigs. Additionally, the myoglobin content of the muscle (and, in consequence, the meat instrumental colour), the insoluble collagen (affecting the textural properties of the meat) and the water losses might be related to the lifetimes of animals, as reported by Mayoral, Dorado, Guillén, Robina, Vivo, Vázquez and Ruiz [
13] for purebred Iberian pigs. It is therefore important to raise awareness of the changes that animal age at slaughter may cause on muscle fibre characteristics and their repercussions on the meat quality of products.
The fibre type of classification is mainly based on the contractile and metabolic performance characteristics. Four major types of muscle fibres are found in the skeletal muscle of adult animals: (1) slow-twitch oxidative or type I, (2) fast-twitch oxido-glycolytic or type IIA, (3) fast-twitch glycolytic (IIB) and (4) fast-twitch oxidative (IIX) [
14]. Type I fibres have a greater oxidative capacity to support sustained muscle contractions, whereas type IIB fibres are predominantly glycolytic fibres that utilise the rapid conversion of glycogen for short bursts of energy. The IIA and IIX types of fibres are between type I and IIB fibres. The sizes and proportions of the different muscle fibres can vary according to various production factors, such as breed [
15], growth performance [
16], physical exercise associated to free-range rearing [
17] or diet [
18], and so, their impact on the quality parameters must be taken into account. The scientific literature has addressed the influence of the genetic background (Iberian purebred vs. Iberian × Duroc crossbred), management and feeding conditions on the muscle fibres of various muscles that are present in ham [
19]. However, to our knowledge, there are no age-related changes in the Iberian × Duroc crossbreed, despite this breed catching the interest of all the players of the Iberian pig industry.
There is a need to generate knowledge about the extent to which fibre types and sizes might be impacted by animal age at slaughter in the various production systems, considering the age ranges that are currently used and are suitable in each production system. The variations in fibre compositions may help understand the variations of some of the properties relating to meat quality and technological aptitude, such as colour, water loss or texture, and their importance in the improvement and control of meat quality without reducing the advantages that the fast growth performance of the Iberian × Duroc crossed animals provide.
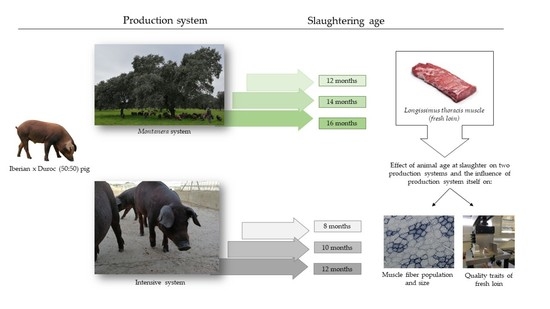
Within this framework, the purpose of the current study was to evaluate the impact of animal age at time of slaughter of two production systems and the effects of the production systems themselves on the muscle fibre characteristics and approximate composition, water losses, instrumental colour and texture properties of the meat.
3. Results
No differences were found between the fibre proportions of the LT muscle on account of the animal age at slaughter in animals reared in either the
Montanera (
Table 2) or intensive system (
Table 3) (
p > 0.05).
As far as fibre diameters are concerned, differences were identified on account of the age at slaughter on the LT muscles of animals reared in the
Montanera system (
Table 2) (
p ≤ 0.05) but not on those of animals reared in the intensive system (
p > 0.05) (
Table 3).
All the fibres of LT muscles from animals slaughtered at the oldest age, i.e., 16 months old, reached a greater diameter in comparison to the muscles from the animals that were slaughtered at 12 and 14 months of age (p ≤ 0.05). In the specific case of type I fibres, an intermediate diameter was identified in muscles from animals slaughtered at 14 months of age with respect to those from animals slaughtered at 12 and 16 months of age (p = 0.017). Thus, the diameters of type I muscle fibres from animals reared in Montanera increased with the age at slaughter (p = 0.017). This diameter was the largest at 16 months, intermediate at 14 months and the smallest at 12 months of age at slaughter (98.38, 89.65 and 82.28 µm, respectively). The highest value of the muscle fibre minor diameter was identified in animals that were slaughtered at 16 months of age: type IIA (96.31 µm; p = 0.001), type IIB (100.20 µm; p = 0.002) and type IIX (96.17 µm; p = 0.000).
The results of the muscle fibre type population and size according to the production system are shown in
Table 4. Type I muscle fibre was predominant in muscles from animals reared in the
Montanera system (
p = 0.026), with 24.16% vs. 18.85%, respectively. Animals being reared in an intensive system did not have their muscles prepared for time-sustained exercising, and it was expected for them to have less type I muscle fibres. On the other hand, the production system only affected the size of type IIB glycolytic fibres, yielding higher values in the muscles from animals reared indoors than those from animals reared in
Montanera (103.42 vs. 88.91 µm of the minor diameter, respectively) (
p = 0.010).
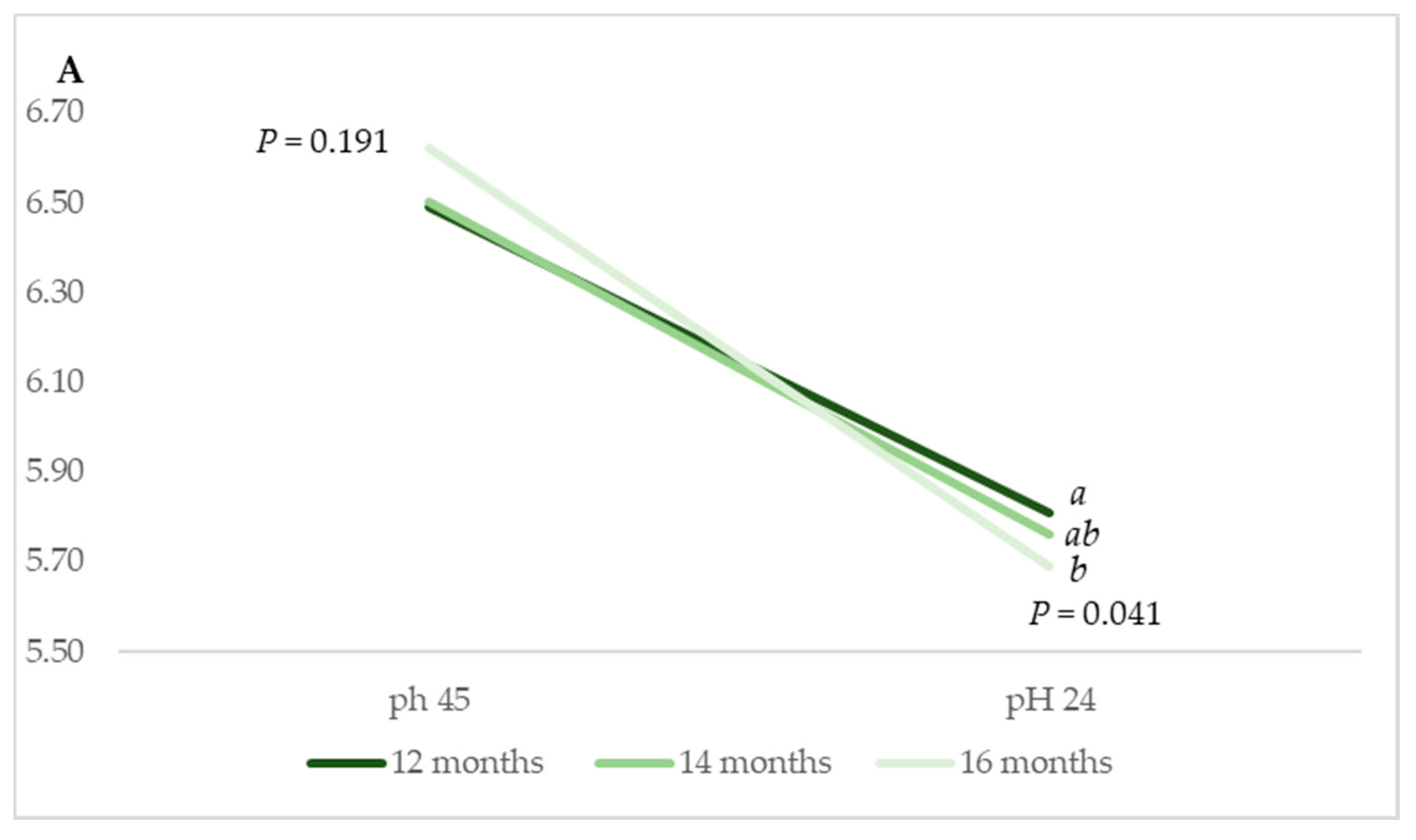
Some differences were identified in terms of pH
24 (
p = 0.041) between muscles from animals reared in
Montanera on account of the animal age at slaughter (
Figure 2A). Muscles from animals that were slaughtered at 12 months of age yielded the highest value, i.e., 5.81, in comparison to 5.76 and 5.69 for LT muscles from animals slaughtered at 14 and 16 months of age, respectively. Additionally, the declining pH rate was more pronounced in LT muscles from animals slaughtered at 16 months of age compared to those of muscles from animals slaughtered at 12 and 14 months old.
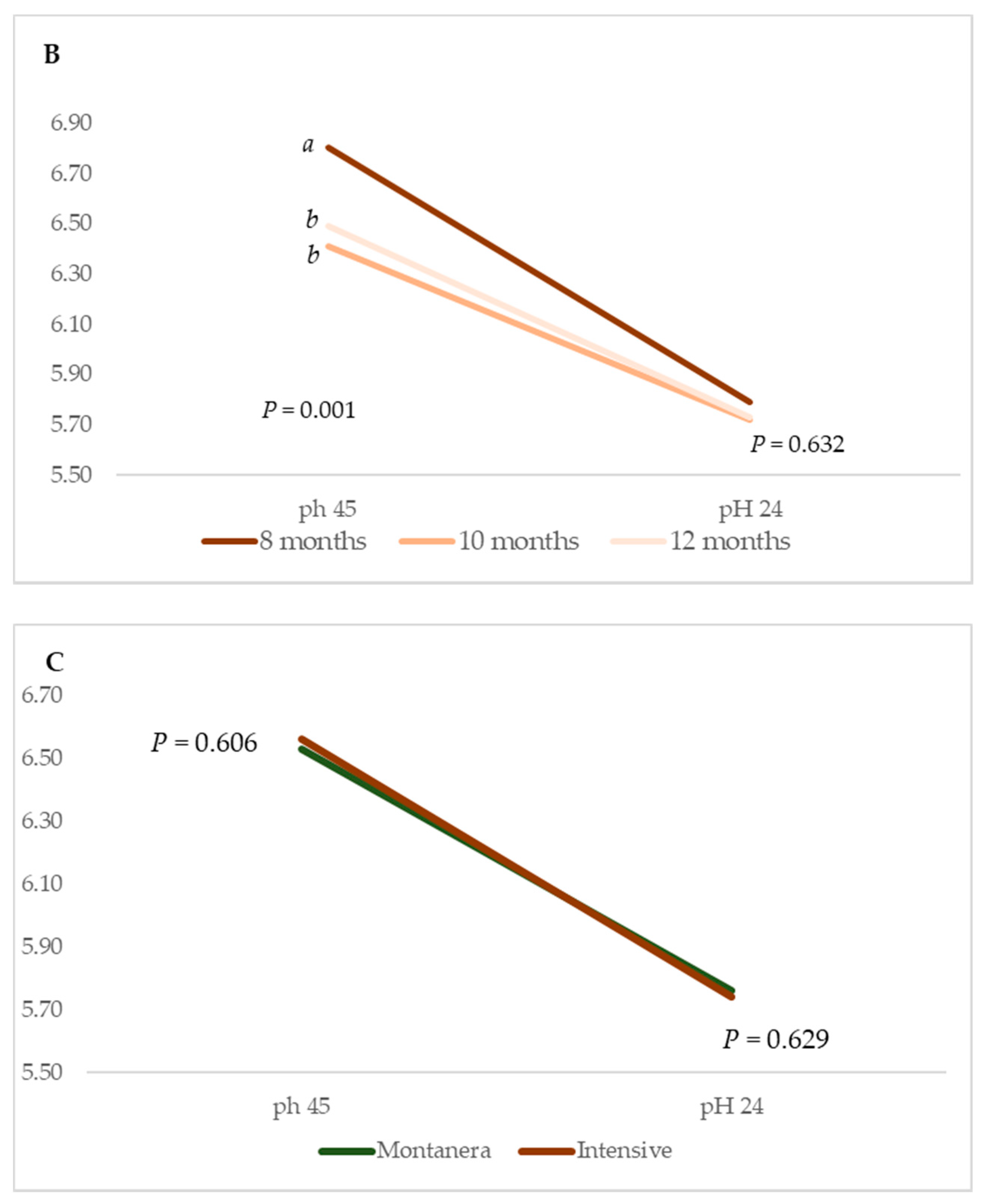
With regards to the animals being reared in intensive systems, differences in the pH were identified 45 min after slaughter. The youngest animals yielded the highest pH value at this stage, i.e., 6.80 vs. 6.41 and 6.49, respectively (
p = 0.001) (
Figure 2B). In this case, the declining pH rate was more pronounced in LT muscles from the youngest animals (8 months).
In contrast, no differences were identified in the pH measurements at 45 min or 24 h later on account of the production system (
p > 0.05) (
Figure 2C).
Table 5,
Table 6 and
Table 7, respectively, represent the way meat quality traits are affected by animal age at slaughter both in the
Montanera and the intensive production systems, as well as the production system itself.
With regards to the
Montanera production system (
Table 5), the data revealed differences found in the DM and myoglobin contents of fresh loins on account of animal age at slaughter (
p = 0.000). Thus, loins from animals that were slaughtered at 16 months of age yielded higher DM and myoglobin contents than loins from animals slaughtered at 12 and 14 months of age. A similar pattern was identified in terms of redness, with loins from animals that were slaughtered at 14 and 16 months of age yielding higher values than those slaughtered at 12 months of age (
p = 0.025). No differences on account of animal age at slaughter were identified for water loss and the textural properties (
p > 0.05).
Some of the meat quality traits evaluated also varied on account of animal age at slaughter when the animals were reared indoors (
Table 6). The IMF content of the fresh loins from animals slaughtered at 12 months of age showed the lowest value (4.65%) with respect to 6.87% and 6.65% yielded by the fresh loins from animals slaughtered at younger ages (8 and 10 months, respectively) (
p = 0.048). The myoglobin content followed a trend similar to that of fresh loins from animals reared in the
Montanera system, with the values increasing as the animals were older at slaughter (
p = 0.033). In contrast, no differences were identified in DM on account of the age at slaughter (
p = 0.436).
Water loss only showed significant differences in terms of cooking loss, and it increased with the age at slaughter, with the highest value being identified in fresh loins from animals slaughtered at 12 months of age (p = 0.002). As far as instrumental colour is concerned, the animal age at slaughter had a significant effect on meat redness. Therefore, higher a* values were identified in loins from animals slaughtered at the oldest age (12 months old) (p = 0.028). In terms of textural properties, differences in WBSF were identified, with fresh loins from animals slaughtered at 12 months of age yielding the highest values in comparison to animals slaughtered at younger ages (p = 0.014).
The meat quality traits of fresh loins were also affected by the production system in which the animals were reared (
Table 7). With regards to the approximate composition, fresh loins from animals reared in the
Montanera system yielded lower IMFs (
p = 0.000) and higher myoglobin contents (
p = 0.014) than animals reared in the intensive system. As far as instrumental colour was concerned, the production system had a significant effect on the lightness, redness and yellowness. Thus, the L* value was higher in fresh loins from animals reared in the intensive system (51.05) (
p = 0.000), whilst the a* (
p = 0.000) and b* (
p = 0.043) coordinates were higher in fresh loins from animals reared in the
Montanera system.
With regard to the textural properties, fresh loins from animals reared in the intensive system yielded lower values of maximum shear force (WBSF) (p = 0.035) but higher hardness (p = 0.000) and chewiness (p = 0.000) values than those from animals under the Montanera system.
4. Discussion
A lower proportion of type I fibres in the LT muscles of animals reared in intensive conditions compared to animals reared in the
Montanera system (
Table 4) was expected. Previous scientific literature agrees on the increase of the oxidative capacity of the muscles as the animal exercises [
9,
10]. Animals reared in the
Montanera system were subjected to sustained exercise for longer periods of time, thus developing more resistance to fatigue and requiring more muscle energy. Thus, the higher type I fibres (slow-twitch oxidation) would be associated with a large oxidative ability to support sustained muscle contractions [
9]. This supports the previous literature on the influence of the muscle function on the fibre-type population [
10,
36].
The lack of differences in the fibre-type populations on account of animal age at slaughter in any of the systems under study (
Table 2 and
Table 3) might be explained, because the most relevant changes occurring during the growth phase of the animals in terms of the fibrillary structure are associated with hypertrophy, this is an increase in size of the fibres [
37]. In fact, significant differences were identified in the minor diameter measurements in all types of fibres of the LT muscle on account of age at slaughter of the animals reared in the
Montanera system (
Table 2). On the other hand, we must take into account that the animals were subjected to feed restrictions during their growing phase in order to be slaughtered at different ages with similar weights. This feed restriction could have impacted the size of the fibres [
38]. Thus, Lefaucher and Ecolan [
38] reported an increase in the fibrillary sizes of the
Longissimus thoracis and
lumborum and
Tibialis cranialis muscles due to feed restrictions in Large White breed pigs with 100 Kg of LW. However, similar to the results obtained in this study, Lefaucher and Ecolan [
38] did not identify variations in the fibre-type populations based on feed restrictions.
The differences in size of the type IIB fibres on account of the production system (
Table 4) could relate to feed type. Specifically, the highest average value identified in LT muscles from animals reared indoors would be explained by the higher intake of proteins that are contained in commercial fodders compared to the proteins provided by acorns and pasture in animals reared in
Montanera (
Table 1), as has been previously reported in other pig breeds [
39]. In this respect, Andrés, Ruiz, Ventanas, Tejeda and Mayoral [
10] also found type I fibres with larger cross-sectional diameters in the
Biceps femoris muscle from animals reared in the intensive system compared to those reared from
Montanera.
With regards to the association between meat quality traits and muscle fibre characteristics, although the muscle pH and the rate of
post-mortem (p.m.) pH decline have been previously reported to depend on the fibre-type composition, the findings of our study did not prove this. Some previous studies agree on the increase of the rate and extent of the p.m. pH decline as the proportion of fast-twitch glycolytic fibres increase in porcine
Longissimus thoracis and
lumborum muscle [
16,
40,
41]. However, despite that this study proved there were pH
24 (
Figure 2A) differences in the meat from animals reared in the
Montanera system and pH
45 (
Figure 2B) differences in the meat from animals reared in the intensive system on account of their age at slaughter, such differences could not be related to the fibre-type populations, as there were no significant differences between them on account of age (
Table 2 and
Table 3). Such discrepancies could relate to the Iberian breed itself—that is, the fact that Iberian is a rustic breed with different pH behaviours in comparison to other improved pig breeds. In any case, the pH values and the pH rate decline identified in the muscles of animals reared in the intensive system and in the
Montanera system can be deemed normal [
41].
On the matter of the approximate composition, the high IMF values in fresh loins from younger animals reared in the intensive system (
Table 6), as well as the higher IMF values in fresh loins from animals under the intensive system compared to those from
Montanera animals (
Table 7), might be associated with the animal growth rate [
42]. Few studies associated a greater IMF content to a higher type I fibre content [
16,
43], but this association was not observed in the current study. More in detail, the decrease of the IMF with the age of the animal in the intensive system could compromise the marbling after the curing process [
44]—dry-cured loin—considered a quality index by consumers [
45].
The fact that there were no differences in water losses on account of animal age at slaughter in animals reared in the
Montanera system (
Table 5) could be explained by the significant impact of IMF, which remained the same as the animals grew up on them [
46]. This association would also explain the smaller cooking losses that were identified in the fresh loins of animals reared in the intensive systems and slaughtered at 8 and 10 months of age in comparison to the animals slaughtered at 12 months of age (
Table 6). The higher cooking losses of fresh Iberian loins from the oldest animals could have an impact on the texture. In fact, an increase in both the shear force measured by the WBSF test and hardness was observed in the latter with respect to the loins from the younger animals, although these differences were only significant for the former. This may condition the acceptance of these for fresh consumption by consumers. The literature has previously reported that fibre-type populations and sizes have an impact on water losses, specifically fast-twitch glycolytic (IIB) [
16], and that hypertrophy [
47] would act in detriment of the WHC. However, this association was not observed in the current study.
The higher a* values in the meat from animals slaughtered at older ages in both production systems (
Table 5 and
Table 6), as well as in the meat from animals reared in the
Montanera in comparison to those reared in the intensive (
Table 7) system, could be explained by the similar behaviour of myoglobin, given the close association between both parameters [
48], which has been specifically identified for the Iberian pig [
7]. At the same time, the increase in redness and in the myoglobin content cannot be associated in our study to the type of muscle fibres, since no differences were found in them on account of age (
Table 2 and
Table 3). On the other hand, as the type I fibre size increased with animal age, although differences were only significant for animals reared in the
Montanera system (
Table 2), the myoglobin content increased (
Table 5), as reported in previous studies [
49,
50,
51]. Age-related colour differences are important, as one of the characteristics of meat and meat products that influences their overall quality is the deep, dark, reddish colour, which is achieved by the myoglobin content [
52].
On the matter of textural properties, these are closely related to muscle fibres. Thus, Ryu and Kim [
53] suggested that, as the fibre size increases, the tenderness decreases, and Renand, Picard, Touraille, Berge and Lepetit [
54] indicated a positive correlation between the tenderness and type I fibre population. This behaviour could explain, at least partially, the differences identified in the meat on account of the production system used for such animals. Specifically, a lower fibre type I population and larger size of type II fibres in LT muscles from animals reared in intensive systems (
Table 4) would give rise to a higher hardness of their meats (
Table 7). However, the findings of this study suggest the impact of other factors on top of the muscle fibre characteristics on the texture of meat. In fact, although the size of all types of fibres in the LT muscles proved to increase with animal age for the animals reared in the
Montanera system, the texture characteristics remained unchanged. This could be explained by other production factors, such as exercise and IMF [
55], the values being the same for the three animal batches. On the other hand, a lower IMF content in the fresh loins from animals reared in the intensive system and slaughtered at 12 months old could be responsible for a higher WBSF [
56].
5. Conclusions
This study is an attempt to shedd light on Iberian fresh meat quality traits and muscle fibre characteristics as affected by animal age at slaughter and production system. Our findings suggest that the fibre-type population of Longissimus thoracis muscles from Iberian × Duroc crossed pigs would not be affected by animal age at slaughter. However, differences in the size of the fibres of the Longissimus thoracis muscles were observed on account of the animal age at slaughter reared under the Montanera system.
In terms of meat quality traits, variations in some quality parameters in the fresh loins from animals under both the Montanera and intensive conditions due to animal slaughter age were found, thus proving the latter to be a factor of variability of the Iberian fresh meat quality. A redder meat, due to a higher content of the pigment myoglobin, could be obtained with the age of the animal regardless of the production system. On the other hand, the increase in age would lead to less fat infiltration, greater losses during cooking and, also, to greater toughness in the case of meats from animals under the intensive system, and therefore, the most tender meat was obtained from animals slaughtered at a younger age (8 and 10 months).
Future studies should evaluate whether the differences observed in the fibrillar structures and quality parameters due to the age at slaughter and production system of the animals are perceived at the sensory level, as well as their impact on the technological curing process.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}