1. Introduction
Weaning can bring enormous stress to piglets, leading to a decline in piglet growth performance [
1]. In recent years, high doses of ZnO have been used to reduce the harm of weaning to piglets [
2]. However, the demand for zinc in piglets is 80–100 mg/kg [
3], and piglets cannot absorb zinc completely, which leads to a large amount of zinc discharged from the body, causing potential harm to the environment [
4]. Announcement No.2625 of the Ministry of Agriculture of China stipulates that the maximum allowable concentration of ZnO in piglets’ diet is 1600 mg/kg. In 2017, the CVMP intends to prohibit the use of high-dose ZnO in piglets within 5 years. Therefore, it is necessary to find a potential alternative to ZnO to maintain the normal growth of weaned piglets.
Some studies have found that tannic acid has good physiological functions: bacteriostasis, anti-diarrhea, anti-oxidation, and anti-inflammation [
5,
6,
7]. However, tannic acid contains a large number of hydroxyl groups [
8], which can easily combine with proteins and digestive enzymes to form insoluble complexes [
9]. Sathe et al. also found that tannic acid can inhibit pepsin activity [
10]. We choose to add stearic acid to hydrolyzed Chinese gallnut tannic acid and to reduce the combination between hydrolyzed Chinese gallnut tannic acid and pepsin. A recent study showed that a dietary supplement of 0.2% tannic acid could effectively alleviate diarrhea in weaned piglets, which may be achieved by improving the integrity and function of intestinal barrier [
11]. In this study, we compared the production performance, diarrhea rate, antioxidant capacity, gut morphology, and intestinal flora of two groups of piglets to explore whether 1899.5 mg/kg GCT (0.2%) could replace 1600 mg/kg ZnO as a new piglet feed.
2. Materials and Methods
2.1. Diets for Piglets
Hydrolytic Chinese gallnut tannic acid was purchased from Ropeedar Biotechnology Co., Ltd. (Shaoxing, Zhejiang, China), which was derived from Chinese gallnut. The hydrolytic tannic acid of Galla chinensis was wrapped by stearic acid, and the effective content of tannic acid was 70.38% (calculated by absolute dry weight).
2.2. Animals and Management
The Animal Experimental Ethics Committee of Guangxi University approved the experiment (Gxu2018-052). Seventy-two weaned piglets (average initial weight 10 ± 0.2 kg) of 31 days old were randomly divided into two groups: (1) A control group, with a basal diet including + 1600 mg/kg ZnO; ZnO and premix were mixed into diet. (2) A treated group, with a basal diet including + 1899.5 mg/kg GCT (0.2%); tannic acid was evenly added to the premix for feeding.
The whole experiment lasted 21 days. Diets were formulated according to nutritional requirements recommended by NRC (2012) and adjusted according to the situation [
3]; detailed information is shown in
Table 1. During the experiment, weaned piglets drank water freely and fed regularly 4 times per day. The weight of every piglet was recorded before fasting at 07:00 a.m. in the morning every week, as was the daily feed intake of each pen. Finally, we calculated an average value to obtain the average daily gain (ADG), average daily feed intake (ADFI), FCR (= average daily feed intake (ADFI)/average daily gain (ADG)), and diarrhea rate. Diarrhea monitoring was assessed by fecal score, recorded daily using the score scheme previously proposed by Girard et al. [
7].
2.3. Sampling and Collection
Blood samples were taken from the jugular vein and put into a heparin anticoagulant tube quickly before feeding (08:00 a.m.) on day 21st. A total of 72 blood samples were collected, 3 of which showed hemolysis due to mis-operation, so only 69 samples were assayed. They were centrifuged at 3000 rpm at 4 °C for 15 min to collect serum and stored in a refrigerator at −80 °C to assay the malondialdehyde (MDA), glutathione (GSH), superoxide dismutase (SOD) and D-lactic acid in serum. MDA, GSH and SOD were assayed with commercial assay kits (Jiangsu Yutong Biological Technology Co., Ltd., Yancheng, China). All blood samples were collected and assayed according to the kit instructions.
2.4. Analysis of Intestinal Morphology and Intestinal Flora
We collected 1–2 cm segments of the middle of duodenum, jejunum, and the end of ileum, and put them into a formalin solution for hematoxylin-eosin staining. The pictures were measured using 40× field of view by a light microscope with a computer-assisted morphometric system. We made the tissue fill the entire field when taking pictures, and ensured that the background light of each photo was consistent. Five intact villi and the crypt near the villus were selected for each slice and measured by Image-Pro Plus 6.0 software.
Pigs were euthanized by the intravenous injection of pentobarbital sodium (lethal dose). The cecal contents were collected by gently squeezing the lumen contents from the tissue into the sterile collection tube, then stored in refrigerator at −80 °C to measure the changes of intestinal flora. Genomic DNA isolation of cecal contents was performed according to the instructions of a DNA Stool Mini Kit (Qiagen, Hilden, Germany). DNA isolation was performed by 2% agarose gel electrophoresis. The bacterial universal V3-V4 region of the 16S rRNA gene was amplified according to polymerase chain reaction barcoded primers 515F (5′-ACTCCTACGGGAGGCAGCAG-3′) and the reverse primer 806R (5′-GGACTACHVGGGTWTCTAAT-3′). PCR was run at 95 °C for three minutes to denaturation, followed by twenty seven cycles of 95 °C for thirty seconds, annealing at 55 °C for thirty seconds, 72 °C for forty-five seconds and a final extension at 72 °C for ten minutes. Briefly, paired-end sequencing was performed on the Illumina MiSeq platform at the Majorbio Bio-Pham Technology (Shanghai, China).
2.5. Statistical Analysis
Analysis was conducted by Excel of Microsoft (Redmond, WA, USA, Office 2019) and analyzed by SPSS 19.0 (SPSS Inc., Chicago, IL, USA, 2009). Firstly, the data were evaluated through the Shapiro–Wilk test to check whether the distribution of the variables exhibited a normal distribution. Then, the variables that showed a normal distribution and a non-normal distribution were analyzed by the independent sample t-test and the Kruskal–Wallis test. Statistical significance was set at p < 0.05 and tendencies at 0.05 ≤ p ≤ 0.10.
4. Discussion
It has always been a controversial issue whether tannic acid can improve the diet utilization rate of piglets. Tannic acid has long been considered an “anti-nutritional” factor. However, Bee et al. have found that hydrolyzed tannins seem to have less impact on growth performance than concentrated tannins [
12]. Kotrotsios et al. have found that high doses of condensed tannin (9.7 g/kg) can reduce ADFI for piglets [
13]. Although high-dose hydrolyzed tannin (4.5 g/kg) fed piglets can also reduce the FCR, it has no effect on ADFI and ADG [
14]. We used stearic acid to wrap hydrolyzed Chinese gallnut tannic acid to reduce its effect on pepsin, but the effect of tannic acid on digestive enzymes in the small intestine is unavoidable [
15]. The results show no influence on ADG, ADFI and FCR between the two diets. It is suggested that some beneficial changes may have taken place in the intestine, but these changes are still unknown.
A previous review explained that tannic acid hydrolysis can reduce the rate and duration of diarrhea in piglets [
16]. Our study also proved this point: compared with the control group, we found that GCT can reduce the diarrhea rate, and the effect is increasingly obvious with the extension of time. Within the first 14 days, the duration of diarrhea in the treatment group was generally 1–2 days, while that in the control group was about 3 days, which is consistent with the previous report [
16]. From 14 to 21 days, the piglets in the control group still had diarrhea, which is different from the results reported by Madec [
17]. We must, however, take into account that Madec’s research was conducted in a pathogen-free environment, not in traditional pig farms.
Piglets are prone to stress in the early stage of weaning; the immune organs in the body are not fully developed and mature, and coupled with the stress of the feeding environment, it is easy to produce free radicals, which reduce the body’s resistance and growth performance [
18]. MDA is a marker of oxidative stress, which reflects the degree of free radical attack on body cells. SOD and GSH are important components of the antioxidant defense system, which can scavenge free radicals or lipid peroxides. Therefore, we also studied the effect of GCT on the antioxidant capacity of piglets. Ye et al. found that tannic acid can improve the antioxidant capacity of piglets [
19]. The data showed that the GCT reduced the MDA and increased the SOD and GSH, improving the antioxidant capacity of piglets and improving weaning stress.
One limitation of our study is that we did not assess the content of butyric acid in the piglets’ intestines. In vitro experiments showed that the decomposition products of tannic acid contain butyric acid [
13]. Butyric acid is the preferred energy substrate for ileal mucosa [
20]. It has been proved that 2.25 mg/kg tannic acid can reduce the depth of ileal crypt and increase the content of butyric acid in intestine [
13]. In agreement with Kotrotsios et al., in our experiment, the depth of ileal crypt decreased in the treated group. The crypts of the small intestine have a secretory function, so reducing its surface may help to reduce the severity of diarrhea in weaned piglets [
21]. Ampting et al. found that tannic acid was beneficial for the intestinal barrier of piglets [
22]. Lactic acid is one of the main products of rapid fermentation of an animal’s stomach, and a large amount of lactic acid can cause subacute rumen acidosis in ruminants [
23]. The production of lactic acid is negatively correlated with animal health; it can pass through the gastrointestinal wall, dissolve in the blood, and finally lead to an increase in the blood [
24]. In agreement with these reports, we found that GCT could significantly reduce D-lactic acid in serum. It is suggested that GCT can improve the intestinal barrier.
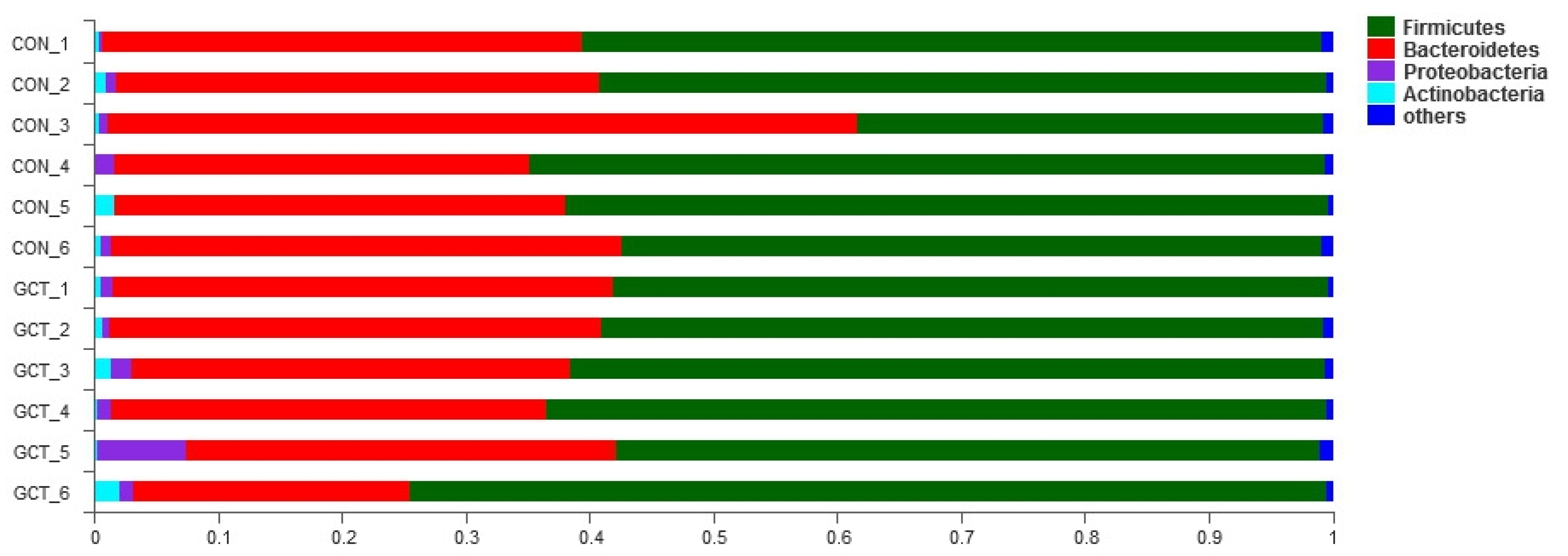
When we analyzed the samples at OTU level, we found no significant influence on the community richness, evenness, or diversity of the two groups. Most of the bacteria in Bacteroidetes and Firmicutes are anaerobic, and the cecum is an absolutely anaerobic environment, so it is predictable that Bacteroidetes and Firmicutes account for about 95% of the cecum bacteria, regardless of the treatment group. This result is similar to that reported by Wang et al. [
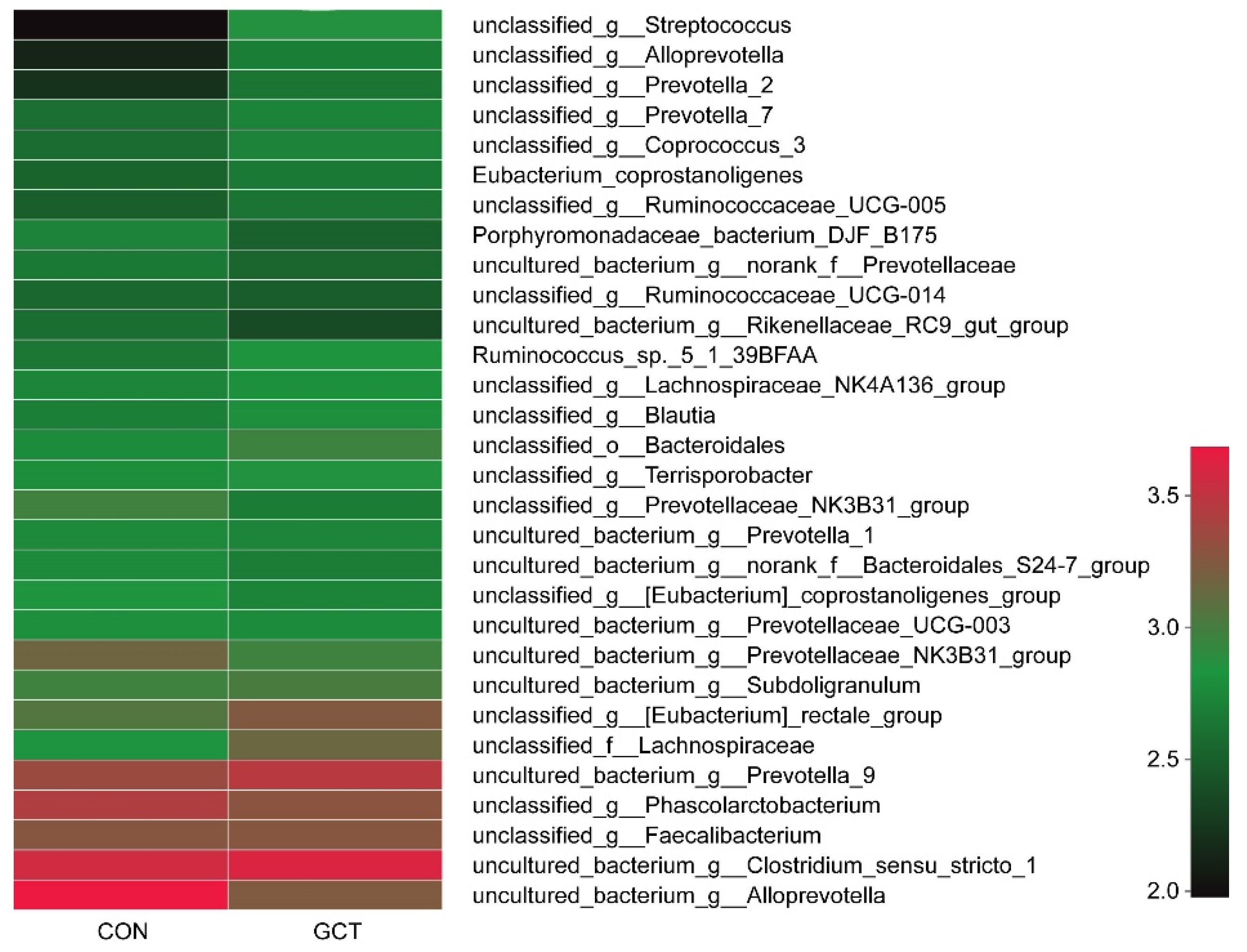
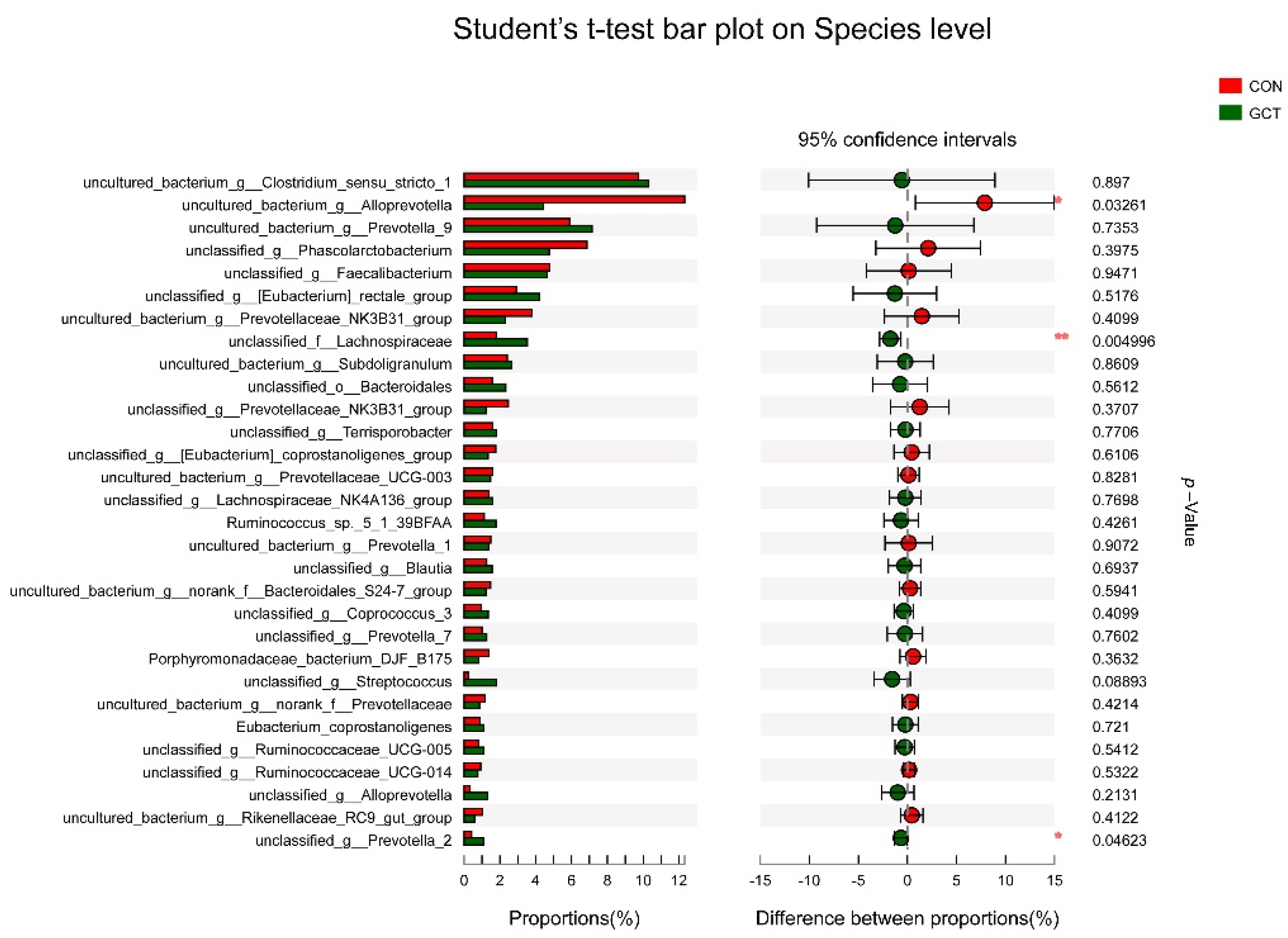
25]. When we analyzed the samples at the species level, we detected that the richness of some bacteria in the GCT diet decreased, such as the opportunistic pathogen
Allprevotella. Therefore, GCT could reduce opportunistic pathogens in the cecum of piglets. When piglets are newly weaned, their food is changed from sow milk to feed containing cellulose and hemicellulose (soybean and corn).
Prevotella is the leader in the degradation of hemicellulose in piglets’ cecum [
25,
26].
Lachnospiraceae also has a strong ability to degrade polysaccharides, even cellulose and hemicellulose [
27]. In our experiment, GCT significantly increased the richness of
Lachnospiraceae and
Prevotella in piglets’ cecum. In addition, some articles have reported that
Lachnospiraceae can reduce the incidence of colitis by reducing the colonization of Clostridium difficile in the intestine [
28,
29].
Lactobacillus amylovorus has a strong ability to hydrolyze starch, and its hydrolysis efficiency is 10 times higher than that of
Lactobacillus plantarum [
30]. In this experiment, GCT increased the richness of
Lactobacillus amylovorus in the cecum of piglets. Overall, GCT may support weaned piglet production performance by promoting the growth of bacteria specialized in the degradation of polysaccharides in the cecum. These results indicate that 1899.5 mg/kg GCT could be an alternative to 1600 mg/kg ZnO in the post-weaning diets of piglets.



 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}