
Testicular Melatonin and Its Pathway in Roe Deer Bucks (Capreolus capreolus) during Pre- and Post-Rut Periods: Correlation with Testicular Involution

 ,
,  , , , ,
, , , ,  ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Testicular Tissue Sampling
2.3. Melatonin Quantification in Testicular Parenchyma
2.4. DHEA Quantification in Testicular Parenchyma
2.5. Western Blot for MT1 and MT2
2.6. RNA Extraction and qPCR for AANAT, ASMT, UCP1 and MT1-2
2.7. Statistical Analysis
3. Results
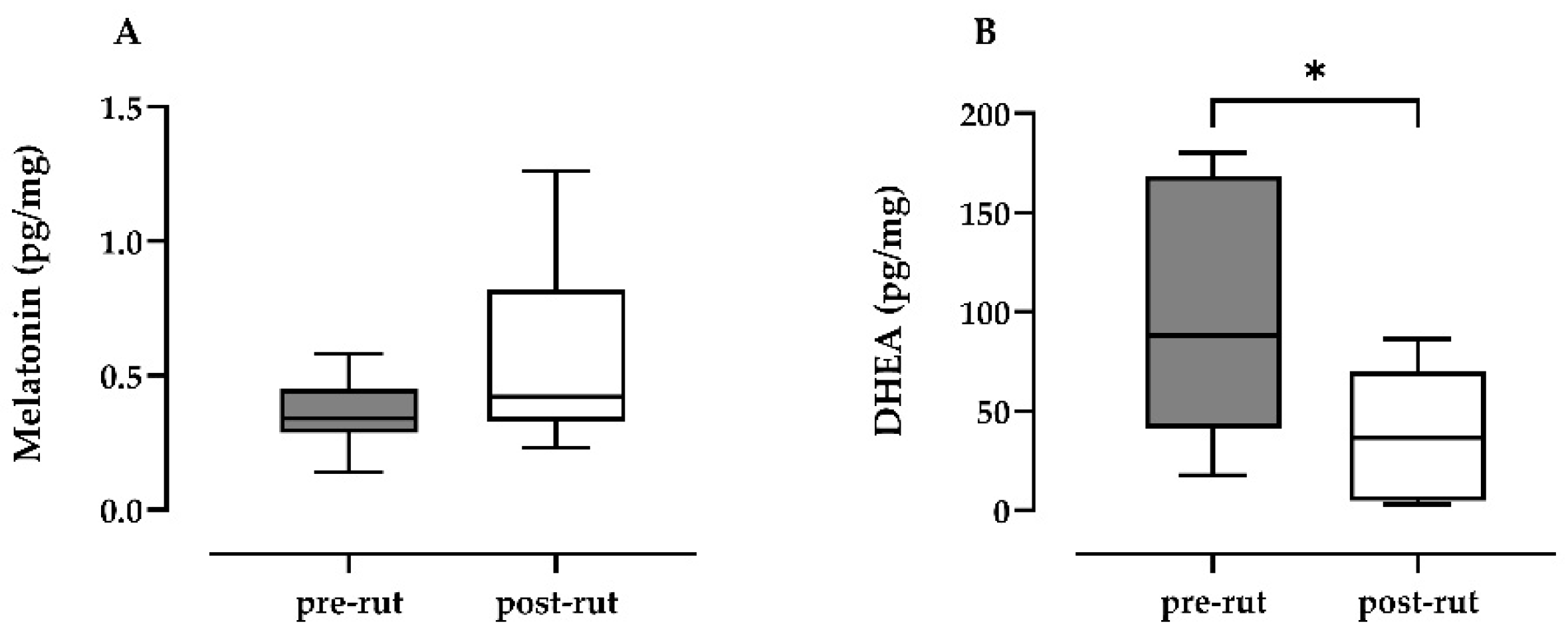
3.1. Melatonin and DHEA Quantification
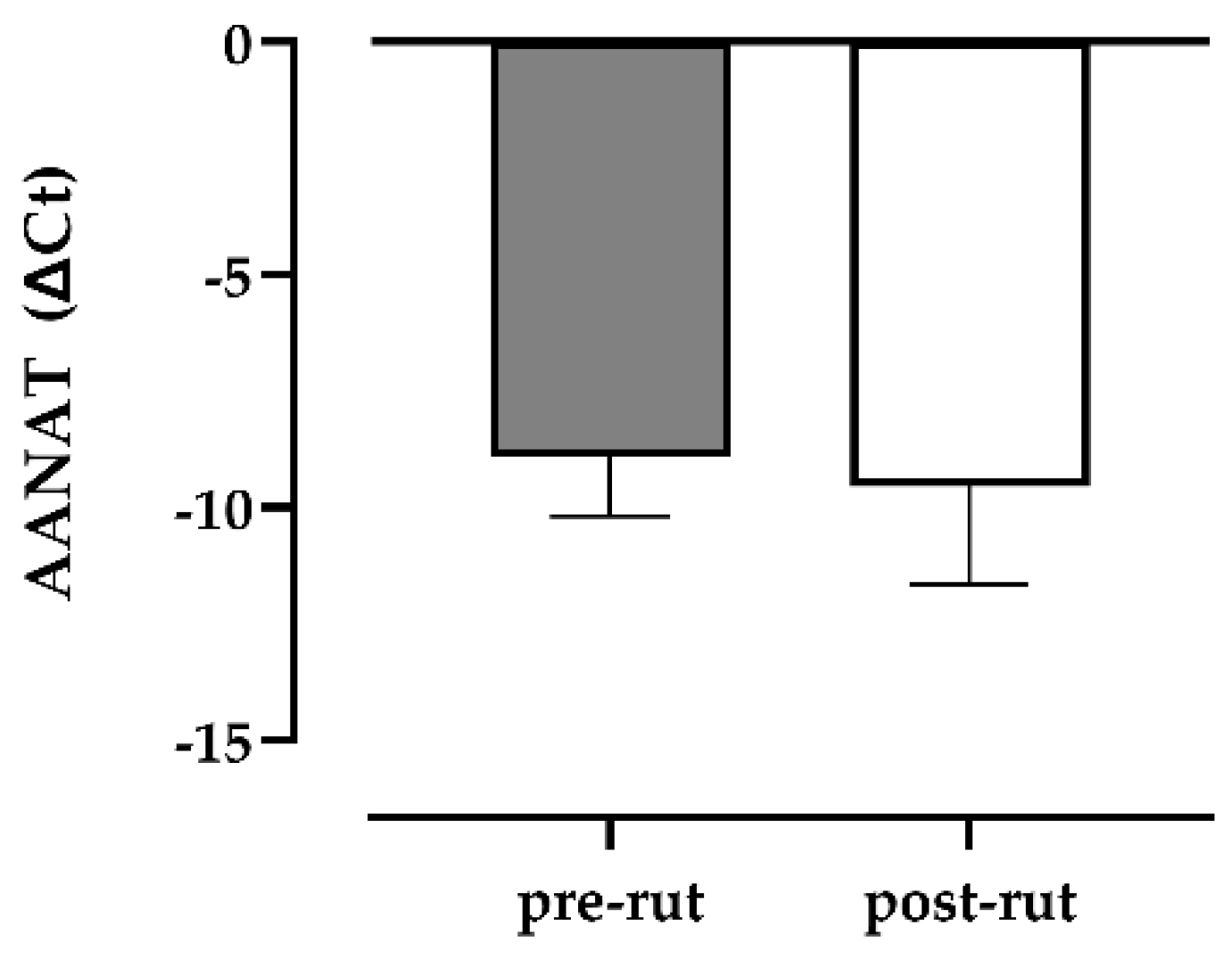
3.2. qPCR for AANAT, ASMT, UCP1, MT1-2
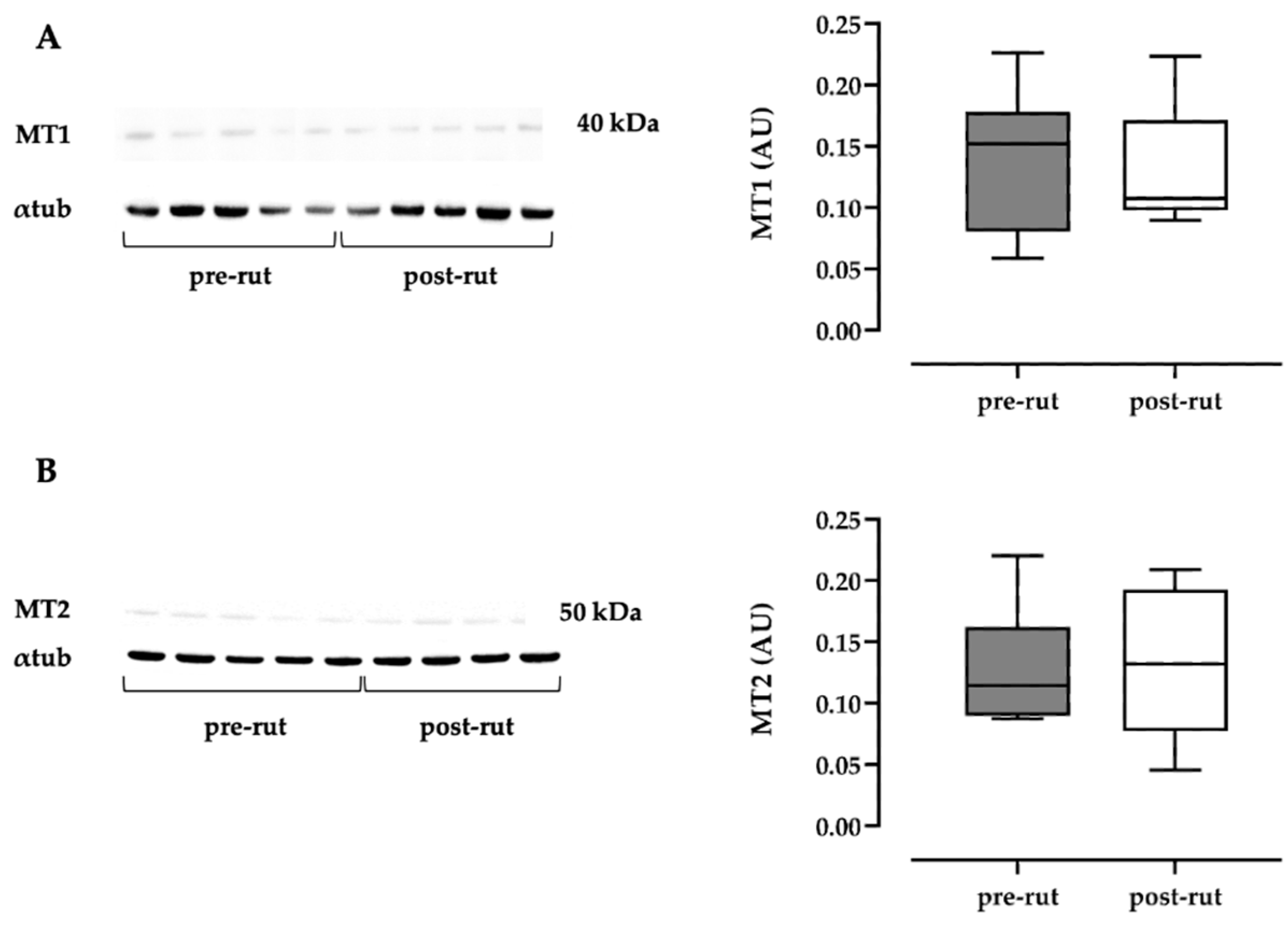
3.3. Western Blot Analysis of MTR1A and MTR1B
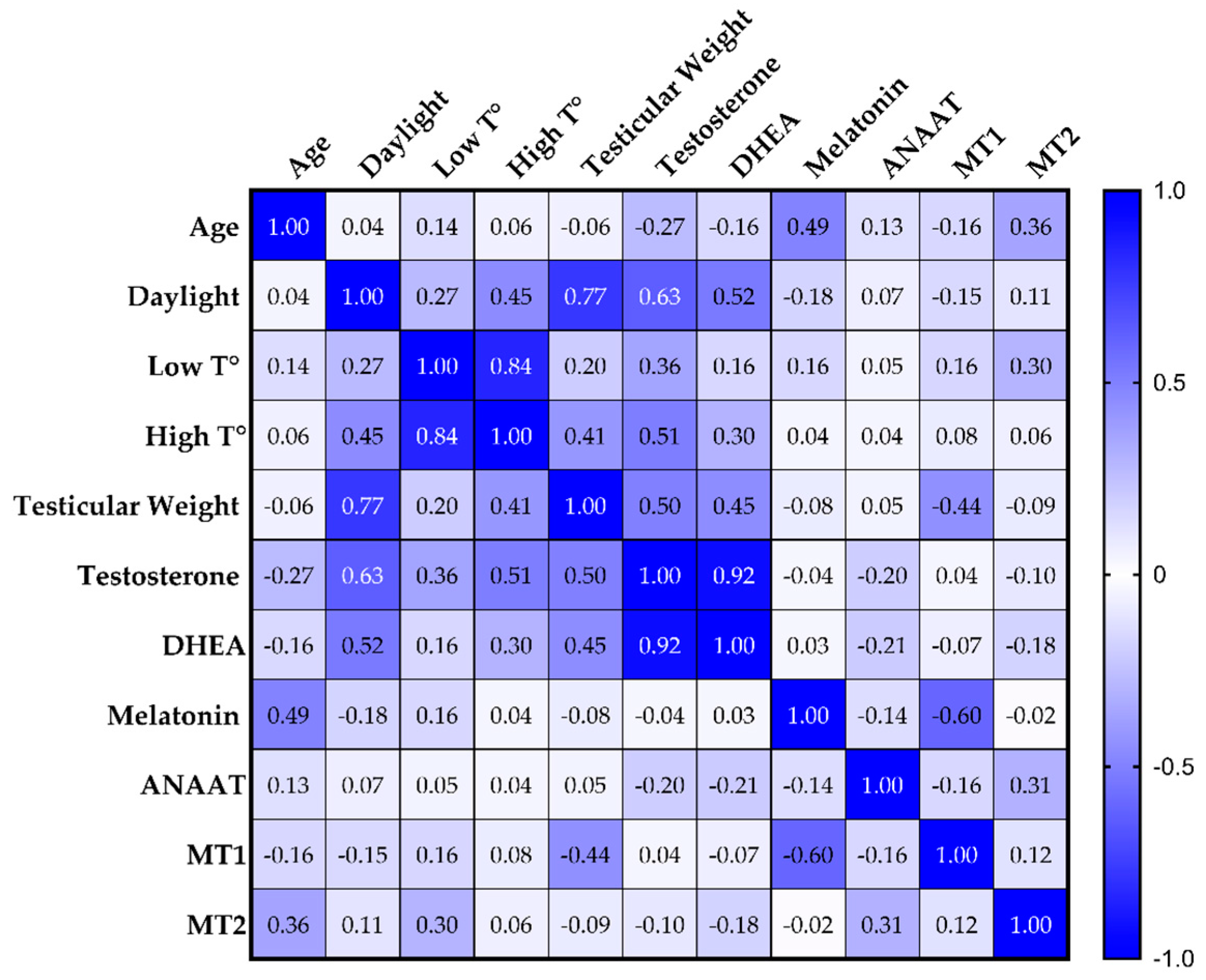
3.4. Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santiago-Moreno, J.; Gómez-Brunet, A.; Toledano-Díaz, A.; Picazo, R.; Gonzalez-Bulnes, A.; López-Sebastián, A. Seasonal Endocrine Changes and Breeding Activity in Mediterranean Wild Ruminants. Reprod. Domest. Anim. 2006, 41, 72–81. [Google Scholar] [CrossRef]
- Borah, B.K.; Renthlei, Z.; Trivedi, A.K. Seasonality in Terai Tree Frog (Polypedates Teraiensis): Role of Light and Temperature in Regulation of Seasonal Breeding. J. Photochem. Photobiol. B 2019, 191, 44–51. [Google Scholar] [CrossRef]
- Mahmoud, I.Y.; Licht, P. Seasonal Changes in Gonadal Activity and the Effects of Stress on Reproductive Hormones in the Common Snapping Turtle, Chelydra Serpentina. Gen. Comp. Endocrinol. 1997, 107, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, A.; Maitra, S.K. The Pineal Gland, but Not Melatonin, Is Associated with the Termination of Seasonal Testicular Activity in an Annual Reproductive Cycle in Roseringed Parakeet Psittacula Krameri. Chronobiol. Int. 2006, 23, 915–933. [Google Scholar] [CrossRef]
- Dawson, A. Annual Gonadal Cycles in Birds: Modeling the Effects of Photoperiod on Seasonal Changes in GnRH-1 Secretion. Front. Neuroendocrinol. 2015, 37, 52–64. [Google Scholar] [CrossRef] [Green Version]
- Pastor, L.M.; Zuasti, A.; Ferrer, C.; Bernal-Mañas, C.M.; Morales, E.; Beltrán-Frutos, E.; Seco-Rovira, V. Proliferation and Apoptosis in Aged and Photoregressed Mammalian Seminiferous Epithelium, with Particular Attention to Rodents and Humans. Reprod. Domest. Anim. 2011, 46, 155–164. [Google Scholar] [CrossRef]
- Dadhich, R.K.; Barrionuevo, F.J.; Real, F.M.; Lupiañez, D.G.; Ortega, E.; Burgos, M.; Jiménez, R. Identification of Live Germ-Cell Desquamation as a Major Mechanism of Seasonal Testis Regression in Mammals: A Study in the Iberian Mole (Talpa Occidentalis). Biol. Reprod. 2013, 88. [Google Scholar] [CrossRef] [PubMed]
- Blottner, S.; Roelants, H.; Waganer, A.; Wenzel, U.D. Testicular Mitosis, Meiosis and Apoptosis in Mink (Mustela Vison) during Breeding and Non-Breeding Seasons. Anim. Reprod. Sci. 1999, 57, 237–249. [Google Scholar] [CrossRef]
- Tabecka-Lonczynska, A.; Mytych, J.; Solek, P.; Kowalewski, M.P.; Koziorowski, M. Seasonal Expression of Insulin-like Growth Factor 1 (IGF-1), Its Receptor IGF-1R and Klotho in Testis and Epididymis of the European Bison (Bison Bonasus, Linnaeus 1758). Theriogenology 2019, 126, 199–205. [Google Scholar] [CrossRef]
- viviD, D.; Bentley, G.E. Seasonal Reproduction in Vertebrates: Melatonin Synthesis, Binding, and Functionality Using Tinbergen’s Four Questions. Molecules 2018, 23, 652. [Google Scholar] [CrossRef] [Green Version]
- Wehr, T.A. Melatonin and Seasonal Rhythms. J. Biol. Rhythms 1997, 12, 518–527. [Google Scholar] [CrossRef]
- Frungieri, M.B.; Calandra, R.S.; Rossi, S.P. Local Actions of Melatonin in Somatic Cells of the Testis. Int. J. Mol. Sci. 2017, 18, 1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiter, R.J.; Tan, D.-X.; Manchester, L.C.; Paredes, S.D.; Mayo, J.C.; Sainz, R.M. Melatonin and Reproduction Revisited. Biol. Reprod. 2009, 81, 445–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiter, R.J. Pineal Melatonin: Cell Biology of Its Synthesis and of Its Physiological Interactions. Endocr. Rev. 1991, 12, 151–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, K.; Deng, S.-L.; Sun, T.-C.; Li, Y.-Y.; Liu, Y.-X. Melatonin Regulates the Synthesis of Steroid Hormones on Male Reproduction: A Review. Molecules 2018, 23, 447. [Google Scholar] [CrossRef] [Green Version]
- Tabecka-Lonczynska, A.; Mytych, J.; Solek, P.; Kulpa, M.; Koziorowski, M. New Insight on the Role of Melatonin Receptors in Reproductive Processes of Seasonal Breeders on the Example of Mature Male European Bison (Bison Bonasus, Linnaeus 1758). J. Photochem. Photobiol. B 2017, 173, 84–91. [Google Scholar] [CrossRef]
- Mukherjee, A.; Haldar, C. Photoperiodic Regulation of Melatonin Membrane Receptor (MT1R) Expression and Steroidogenesis in Testis of Adult Golden Hamster, Mesocricetus Auratus. J. Photochem. Photobiol. B 2014, 140, 374–380. [Google Scholar] [CrossRef]
- Yang, W.-C.; Tang, K.-Q.; Fu, C.-Z.; Riaz, H.; Zhang, Q.; Zan, L.-S. Melatonin Regulates the Development and Function of Bovine Sertoli Cells via Its Receptors MT1 and MT2. Anim. Reprod. Sci. 2014, 147, 10–16. [Google Scholar] [CrossRef]
- González-Arto, M.; Aguilar, D.; Gaspar-Torrubia, E.; Gallego, M.; Carvajal-Serna, M.; Herrera-Marcos, L.V.; Serrano-Blesa, E.; Hamilton, T.R.D.S.; Pérez-Pé, R.; Muiño-Blanco, T.; et al. Melatonin MT₁ and MT₂ Receptors in the Ram Reproductive Tract. Int. J. Mol. Sci. 2017, 18, 662. [Google Scholar] [CrossRef]
- Izzo, G.; Francesco, A.; Ferrara, D.; Campitiello, M.R.; Serino, I.; Minucci, S.; d’Istria, M. Expression of Melatonin (MT1, MT2) and Melatonin-Related Receptors in the Adult Rat Testes and during Development. Zygote Camb. Engl. 2010, 18, 257–264. [Google Scholar] [CrossRef]
- Gonzalez-Arto, M.; Hamilton, T.R.D.S.; Gallego, M.; Gaspar-Torrubia, E.; Aguilar, D.; Serrano-Blesa, E.; Abecia, J.A.; Pérez-Pé, R.; Muiño-Blanco, T.; Cebrián-Pérez, J.A.; et al. Evidence of Melatonin Synthesis in the Ram Reproductive Tract. Andrology 2016, 4, 163–171. [Google Scholar] [CrossRef] [Green Version]
- Tijmes, M.; Pedraza, R.; Valladares, L. Melatonin in the Rat Testis: Evidence for Local Synthesis. Steroids 1996, 61, 65–68. [Google Scholar] [CrossRef]
- Frungieri, M.B.; Gonzalez-Calvar, S.I.; Rubio, M.; Ozu, M.; Lustig, L.; Calandra, R.S. Serotonin in Golden Hamster Testes: Testicular Levels, Immunolocalization and Role during Sexual Development and Photoperiodic Regression-Recrudescence Transition. Neuroendocrinology 1999, 69, 299–308. [Google Scholar] [CrossRef]
- Frungieri, M.B.; Zitta, K.; Pignataro, O.P.; Gonzalez-Calvar, S.I.; Calandra, R.S. Interactions between Testicular Serotoninergic, Catecholaminergic, and Corticotropin-Releasing Hormone Systems Modulating CAMP and Testosterone Production in the Golden Hamster. Neuroendocrinology 2002, 76, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Blottner, S.; Hingst, O.; Meyer, H.H.D. Seasonal Spermatogenesis and Testosterone Production in Roe Deer (Capreolus Capreolus). J. Reprod. Fertil. 1996, 108, 299–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schams, D.; Barth, D. Annual Profiles of Reproductive Hormones in Peripheral Plasma of the Male Roe Deer (Capreolus Capreolus). J. Reprod. Fertil. 1982, 66, 463–468. [Google Scholar] [CrossRef] [Green Version]
- Roelants, H.; Schneider, F.; Göritz, F.; Streich, J.; Blottner, S. Seasonal Changes of Spermatogonial Proliferation in Roe Deer, Demonstrated by Flow Cytometric Analysis of c-Kit Receptor, in Relation to Follicle-Stimulating Hormone, Luteinizing Hormone, and Testosterone. Biol. Reprod. 2002, 66, 305–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blottner, S.; Schön, J.; Roelants, H. Apoptosis Is Not the Cause of Seasonal Testicular Involution in Roe Deer. Cell Tissue Res. 2007, 327, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Schön, J.; Göritz, F.; Streich, J.; Blottner, S. Histological Organization of Roe Deer Testis throughout the Seasonal Cycle: Variable and Constant Components of Tubular and Interstitial Compartment. Anat. Embryol. 2004, 208, 151–159. [Google Scholar] [CrossRef]
- Ventrella, D.; Elmi, A.; Barone, F.; Carnevali, G.; Govoni, N.; Bacci, M.L. Hair Testosterone and Cortisol Concentrations in Pre- and Post-Rut Roe Deer Bucks: Correlations with Blood Levels and Testicular Morphometric Parameters. Animals 2018, 8, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schön, J.; Blottner, S. Estrogens Are Involved in Seasonal Regulation of Spermatogenesis and Sperm Maturation in Roe Deer (Capreolus Capreolus). Gen. Comp. Endocrinol. 2008, 159, 257–263. [Google Scholar] [CrossRef]
- Sempéré, A.J.; Mauget, R.; Bubenik, G.A. Influence of Photoperiod on the Seasonal Pattern of Secretion of Luteinizing Hormone and Testosterone and on the Antler Cycle in Roe Deer (Capreolus Capreolus). J. Reprod. Fertil. 1992, 95, 693–700. [Google Scholar] [CrossRef] [Green Version]
- Kozioł, K.; Koziorowski, M. Steroid Hormones in Peripheral Blood Plasma and Androgen Receptors in Testis and Epididymis of Roe Deer Male (Capreolus Capreolus) during the Reproduction Season. Small Rumin. Res. 2013, 115, 86–93. [Google Scholar] [CrossRef]
- Klonisch, T.; Schön, J.; Hombach-Klonisch, S.; Blottner, S. The Roe Deer as a Model for Studying Seasonal Regulation of Testis Function. Int. J. Androl. 2006, 29, 122–128. [Google Scholar] [CrossRef]
- Elmi, A.; Zannoni, A.; Govoni, N.; Bertocchi, M.; Forni, M.; Ventrella, D.; Bacci, M.L. Uncovering the Physiological Mechanisms Underlying the Roe Deer (Capreolus Capreolus) Testicular Cycle: Analyses of Gelatinases and VEGF Patterns and Correlation with Testes Weight and Testosterone. Animals 2020, 10, 444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koziorowska-Gilun, M.; Fraser, L.; Gilun, P.; Koziorowski, M.; Kordan, W. Activity of Antioxidant Enzymes and Their MRNA Expression in Different Reproductive Tract Tissues of the Male Roe Deer (Capreolus Capreolus) during the Pre-Rut and Rut Seasons. Small Rumin. Res. 2015, 129, 97–103. [Google Scholar] [CrossRef]
- Kozioł, K.; Broda, D.; Romerowicz-Misielak, M.; Nowak, S.; Koziorowski, M. Melatonin Concentration in Peripheral Blood and Melatonin Receptors (MT1 and MT2) in the Testis and Epididymis of Male Roe Deer during Active Spermatogenesis. Theriogenology 2020, 149, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Munley, K.M.; Deyoe, J.E.; Ren, C.C.; Demas, G.E. Melatonin Mediates Seasonal Transitions in Aggressive Behavior and Circulating Androgen Profiles in Male Siberian Hamsters. Horm. Behav. 2020, 117, 104608. [Google Scholar] [CrossRef]
- Elmi, A.; Galligioni, V.; Govoni, N.; Bertocchi, M.; Aniballi, C.; Bacci, M.L.; Sánchez-Morgado, J.M.; Ventrella, D. Quantification of Hair Corticosterone, DHEA and Testosterone as a Potential Tool for Welfare Assessment in Male Laboratory Mice. Anim. Open Access J. 2020, 10, 2408. [Google Scholar] [CrossRef]
- González-Arto, M.; Vicente-Carrillo, A.; Martínez-Pastor, F.; Fernández-Alegre, E.; Roca, J.; Miró, J.; Rigau, T.; Rodríguez-Gil, J.E.; Pérez-Pé, R.; Muiño-Blanco, T.; et al. Melatonin Receptors MT1 and MT2 Are Expressed in Spermatozoa from Several Seasonal and Nonseasonal Breeder Species. Theriogenology 2016, 86, 1958–1968. [Google Scholar] [CrossRef]
- Barajas-López, C.; Peres, A.L.; Espinosa-Luna, R.; Reyes-Vázquez, C.; Prieto-Gómez, B. Melatonin Modulates Cholinergic Transmission by Blocking Nicotinic Channels in the Guinea-Pig Submucous Plexus. Eur. J. Pharmacol. 1996, 312, 319–325. [Google Scholar] [CrossRef]
- Xiao, L.; Hu, J.; Zhao, X.; Song, L.; Zhang, Y.; Dong, W.; Zhang, Q.; Ma, Y.; Li, F. Expression of Melatonin and Its Related Synthase and Membrane Receptors in the Oestrous Corpus Luteum and Corpus Luteum Verum of Sheep. Reprod. Domest. Anim. Zuchthyg. 2018, 53, 1142–1148. [Google Scholar] [CrossRef]
- Stefulj, J.; Hörtner, M.; Ghosh, M.; Schauenstein, K.; Rinner, I.; Wölfler, A.; Semmler, J.; Liebmann, P.M. Gene Expression of the Key Enzymes of Melatonin Synthesis in Extrapineal Tissues of the Rat. J. Pineal Res. 2001, 30, 243–247. [Google Scholar] [CrossRef]
- Frungieri, M.B.; Mayerhofer, A.; Zitta, K.; Pignataro, O.P.; Calandra, R.S.; Gonzalez-Calvar, S.I. Direct Effect of Melatonin on Syrian Hamster Testes: Melatonin Subtype 1a Receptors, Inhibition of Androgen Production, and Interaction with the Local Corticotropin-Releasing Hormone System. Endocrinology 2005, 146, 1541–1552. [Google Scholar] [CrossRef]
- Lan, M.; Zhang, Y.; Wan, X.; Pan, M.-H.; Xu, Y.; Sun, S.-C. Melatonin Ameliorates Ochratoxin A-Induced Oxidative Stress and Apoptosis in Porcine Oocytes. Environ. Pollut. 2020, 256, 113374. [Google Scholar] [CrossRef] [PubMed]
- Acuña-Castroviejo, D.; Escames, G.; Venegas, C.; Díaz-Casado, M.E.; Lima-Cabello, E.; López, L.C.; Rosales-Corral, S.; Tan, D.-X.; Reiter, R.J. Extrapineal Melatonin: Sources, Regulation, and Potential Functions. Cell. Mol. Life Sci. CMLS 2014, 71, 2997–3025. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pre-Rut | Post-Rut | |||
|---|---|---|---|---|
| Mean (SD) | Min; Max | Mean (SD) | Min; Max | |
| Animals | ||||
| Age (months) § | 39 (15) | 24; 72 | 32 (10) | 15; 48 |
| Testis | ||||
| Weight (g) § * | 20.47 (5.26) | 10.24; 26.61 | 11.11 (5.26) | 7.20; 23.42 |
| Testosterone (pg/mg) § * | 917.39 (676.30) | 248.87; 2419.75 | 281.18 (261.18) | 48.22; 848.00 |
| Melatonin (pg/mg) | 0.36 (0.13) | 0.14; 0.58 | 0.59 (0.34) | 0.23; 1.26 |
| DHEA (pg/mg) * | 135.15 (130.00) | 17.72; 443.6 | 36.20 (34.16) | 3.1; 86.25 |
| Environmental data | ||||
| T min (°C) | 15.00 (1.32) | 13; 17 | 12.56 (4.85) | 5; 17 |
| T max (°C) | 28.89 (2.85) | 23; 32 | 26.44 (4.50) | 19; 33 |
| Daylight | 13 h 52 min (14 min) | 13 h 28 min; 14 h 12 min | 10 h 41 min (52 min) | 10 h 30 min; 12 h 11 min |
| Gene expression | ||||
| AANAT (ΔCT) | −8.92 (1.27) | −9.91; −5.99 | −9.55 (2.11) | −13.05; −7.42 |
| Western blot | ||||
| MT1 (AU) | 0.136 (0.056) | 0.059; 0.226 | 0.133 (0.047) | 0.090; 0.224 |
| MT2 (AU) | 0.127 (0.047) | 0.087; 0.220 | 0.129 (0.058) | 0.045; 0.209 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elmi, A.; Govoni, N.; Zannoni, A.; Bertocchi, M.; Bernardini, C.; Forni, M.; Ventrella, D.; Bacci, M.L. Testicular Melatonin and Its Pathway in Roe Deer Bucks (Capreolus capreolus) during Pre- and Post-Rut Periods: Correlation with Testicular Involution. Animals 2021, 11, 1874. https://doi.org/10.3390/ani11071874
Elmi A, Govoni N, Zannoni A, Bertocchi M, Bernardini C, Forni M, Ventrella D, Bacci ML. Testicular Melatonin and Its Pathway in Roe Deer Bucks (Capreolus capreolus) during Pre- and Post-Rut Periods: Correlation with Testicular Involution. Animals. 2021; 11(7):1874. https://doi.org/10.3390/ani11071874
Chicago/Turabian StyleElmi, Alberto, Nadia Govoni, Augusta Zannoni, Martina Bertocchi, Chiara Bernardini, Monica Forni, Domenico Ventrella, and Maria Laura Bacci. 2021. "Testicular Melatonin and Its Pathway in Roe Deer Bucks (Capreolus capreolus) during Pre- and Post-Rut Periods: Correlation with Testicular Involution" Animals 11, no. 7: 1874. https://doi.org/10.3390/ani11071874