
Abiotic and Biotic Influences on the Movement of Reintroduced Chinese Giant Salamanders (Andrias davidianus) in Two Montane Rivers


 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
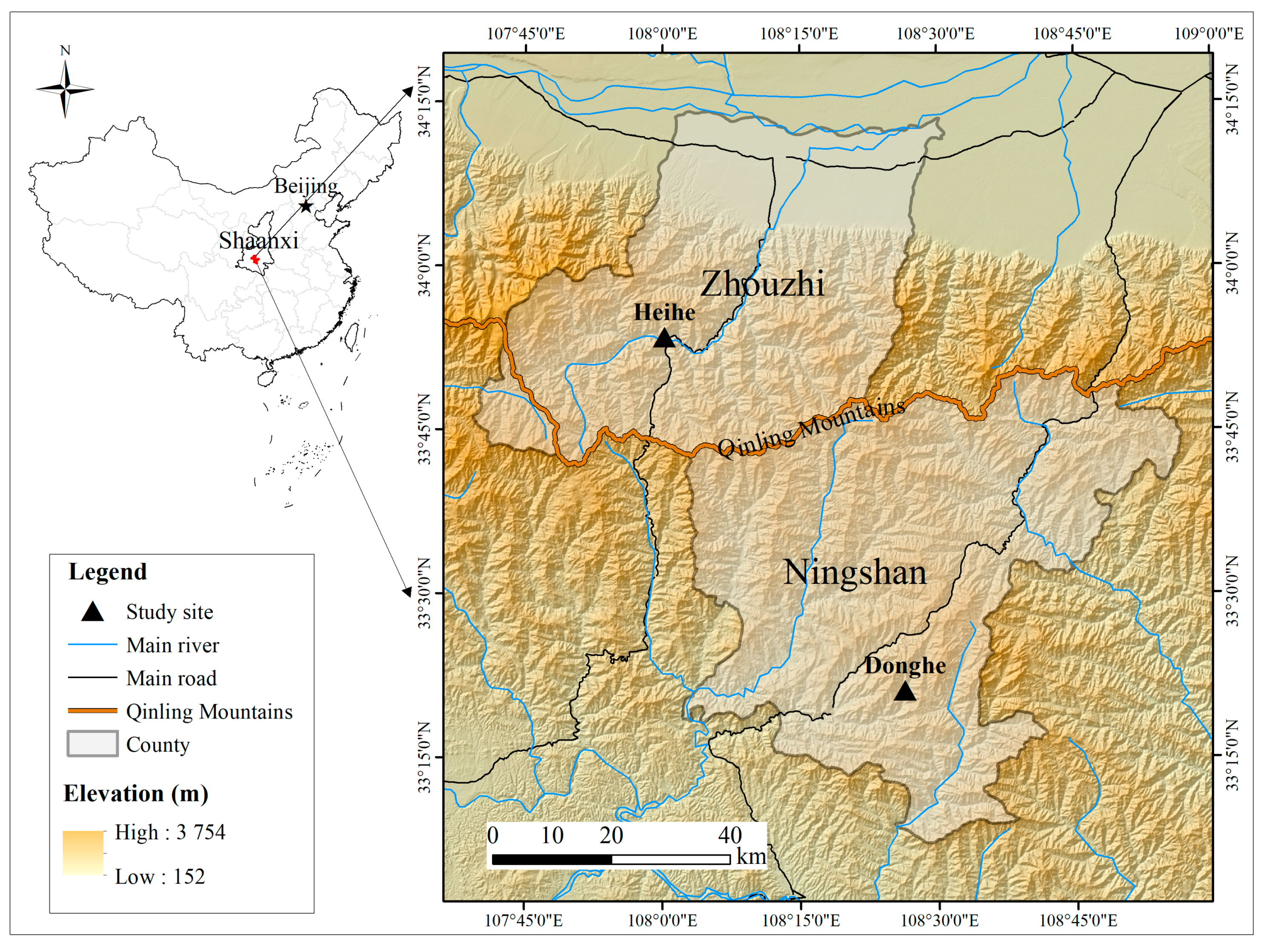
2.1. Study Area and Animals
2.2. Movement Monitoring and Abiotic Data Collection
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nathan, R.; Getz, W.M.; Revilla, E.; Holyoak, M.; Kadmon, R.; Saltz, D.; Smouse, P.E. A movement ecology paradigm for unifying organismal movement research. Proc. Natl. Acad. Sci. USA 2008, 105, 19052–19059. [Google Scholar] [CrossRef] [Green Version]
- Jeltsch, F.; Bonte, D.; Pe’er, G.; Reineking, B.; Leimgruber, P.; Balkenhol, N.; Schroder, B.; Buchmann, C.M.; Mueller, T.; Blaum, N.; et al. Integrating movement ecology with biodiversity research—Exploring new avenues to address spatiotemporal biodiversity dynamics. Mov. Ecol. 2013, 1, 6. [Google Scholar] [CrossRef] [Green Version]
- Kays, R.; Crofoot, M.C.; Jetz, W.; Wikelski, M. Terrestrial animal tracking as an eye on life and planet. Science 2015, 348, aaa2478. [Google Scholar] [CrossRef] [Green Version]
- Richardson, K.M.; Doerr, V.; Ebrahimi, M.; Lovegrove, T.G.; Parker, K.A. Considering dispersal in reintroduction and restoration planning. In Advances in Reintroduction Biology of Australian and New Zealand Fauna; CSIRO Publishing: Clayton South, VIC, Australia, 2015; pp. 59–72. [Google Scholar]
- Armstrong, D.P.; Seddon, P.J. Directions in reintroduction biology. Trends Ecol. Evol. 2008, 23, 20–25. [Google Scholar] [CrossRef]
- Seddon, P.J.; Griffiths, C.J.; Soorae, P.S.; Armstrong, D.P. Reversing defaunation: Restoring species in a changing world. Science 2014, 345, 406–412. [Google Scholar] [CrossRef]
- Knox, C.D.; Monks, J.M. Penning prior to release decreases post-translocation dispersal of jewelled geckos. Anim. Conserv. 2014, 17, 18–26. [Google Scholar] [CrossRef]
- Germano, J.M.; Bishop, P.J. Suitability of amphibians and reptiles for translocation. Conserv. Biol. 2009, 23, 7–15. [Google Scholar] [CrossRef]
- Dumont, C.P.; Himmelman, J.H.; Russell, M.P. Daily movement of the sea urchin Stronavlocentrotus droebachiensis in different subtidal habitats in eastern Canada. Mar. Ecol. Prog. Ser. 2006, 317, 87–99. [Google Scholar] [CrossRef] [Green Version]
- García-Jiménez, R.; Pérez-García, J.M.; Margalida, A. Drivers of daily movement patterns affecting an endangered vulture flight activity. BMC Ecol. 2018, 18, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Sitthipornvorakul, E.; Waongenngarm, P.; Lohsoonthorn, V.; Janwantanakul, P. Is the number of daily walking steps in sedentary workers affected by age, gender, body mass index, education, and overall energy expenditure? Work 2020, 66, 637–644. [Google Scholar] [CrossRef]
- Drabik-Hamshare, M.; Downs, C.T. Movement of leopard tortoises in response to environmental and climatic variables in a semi-arid environment. Mov. Ecol. 2017, 5, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celestino, L.F.; Sanz-Ronda, F.J.; Kashiwaqui, E.A.L.; Celestino, E.F.; Makrakis, M.C.; Makrakis, S. Daily movement behavior of two Neotropical armored catfish species (Ancistrus aff. cirrhosus Valenciennes, 1836 and Hypostomus ancistroides Ihering, 1911) at a road-stream crossing culvert. J. Appl. Ichthyol. 2017, 33, 1092–1099. [Google Scholar] [CrossRef]
- Kay, S.L.; Fischer, J.W.; Monaghan, A.J.; Beasley, J.C.; Boughton, R.; Campbell, T.A.; Cooper, S.M.; Ditchkoff, S.S.; Hartley, S.B.; Kilgo, J.C.; et al. Quantifying drivers of wild pig movement across multiple spatial and temporal scales. Mov. Ecol. 2017, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Perez, A.U.; Schmitter-Soto, J.J.; Adams, A.J.; Herrera-Pavón, R.L. Influence of environmental variables on abundance and movement of bonefish (Albula vulpes) in the Caribbean Sea and a tropical estuary of Belize and Mexico. Environ. Biol. Fish. 2019, 102, 1421–1434. [Google Scholar] [CrossRef]
- Parlin, A.F.; Nardone, J.A.; Kelly Dougherty, J.; Rebein, M.; Safi, K.; Schaeffer, P.J. Activity and movement of free-living box turtles are largely independent of ambient and thermal conditions. Mov. Ecol. 2018, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Zhao, E.M. China Red Data Book of Endangered Animals: Amphibia and Reptilia; Science Press: Beijing, China, 1998. [Google Scholar]
- Liang, G.; Geng, B.R.; Zhao, E.M. Andrias davidianus. In The IUCN Red List of Threatened Species; 2004; Available online: https://dx.doi.org/10.2305/IUCN.UK.2004.RLTS.T1272A3375181.en (accessed on 30 March 2017).
- Wang, X.M.; Zhang, K.J.; Wang, Z.H.; Ding, Y.Z.; Wu, W.; Huang, S. The decline of the Chinese giant salamander Andrias davidianus and implications for its conservation. Oryx 2004, 38, 197–202. [Google Scholar] [CrossRef] [Green Version]
- National Forestry and Grassland Administration; Ministry of Agriculture and Rural Affairs of the PRC. Lists of Wild Animals under Special State Protection in China. Chin. J. Wildl. 2021, 42, 605–640. [Google Scholar] [CrossRef]
- Hu, X.L. An ecological study of megalobatrachus from MT. Dabie in Anhui. J. Anhui Univ. Nat. Sci. Ed. 1987, 1, 69–71. (In Chinese) [Google Scholar]
- Song, M.T. Food habit of great salamander of China. Chin. J. Zool. 1994, 29, 38–42. (In Chinese) [Google Scholar]
- Cunningham, A.A.; Turvey, S.T.; Zhou, F.; Meredith, H.M.R.; Guan, W.; Liu, X.L.; Sun, C.M.; Wang, Z.Q.; Wu, M.Y. Development of the Chinese giant salamander Andrias davidianus farming industry in Shaanxi Province, China: Conservation threats and opportunities. Oryx 2015, 50, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Jiang, W.; Wang, Q.J.; Zhao, H.; Zhang, H.X.; Marcec, R.M.; Willard, S.T.; Kouba, A.J. Reintroduction and post-release survival of a living fossil: The Chinese giant salamander. PLoS ONE 2016, 11, e0156715. [Google Scholar] [CrossRef]
- He, D.; Zhu, W.M.; Zeng, W.; Lin, J.; Ji, Y.; Wang, Y.; Zhang, C.; Lu, Y.; Zhao, D.Q.; Su, N.; et al. Nutritional and medicinal characteristics of Chinese giant salamander (Andrias davidianus) for applications in healthcare industry by artificial cultivation: A review. Food Sci. Hum. Well. 2018, 7, 1–10. [Google Scholar] [CrossRef]
- Nickerson, M.A.; Mays, C.E. A study of the Ozark hellbender Cryptobranchus alleganiensis bishopi. Ecology 1973, 54, 1164–1165. [Google Scholar] [CrossRef]
- Burgmeier, N.G.; Sutton, T.M.; Williams, R.N. Spatial ecology of the Eastern Hellbender (Cryptobranchus alleganiensis alleganiensis) in Indiana. Herpetologica 2011, 67, 135–145. [Google Scholar] [CrossRef]
- Larson, K.A.; Gall, B.G.; Briggler, J.T. The use of gastric transmitters to locate nests and study movement pattern of breeding male Ozark hellbenders. Herpetol. Rev. 2013, 44, 434–439. [Google Scholar]
- Taguchi, Y. Seasonal movements of the Japanese giant salamander (Andrias japonicus): Evidence for possible breeding migration by this stream-dwelling amphibian. Jpn. J. Ecol. Jpn. 2009, 59, 117–128. [Google Scholar]
- Zheng, H.X.; Wang, X.M. Telemetric data reveals ecolgoically adaptive behavior of captive raised Chinese giant salamanders when reintroduced into their native habitat. Asian Herpetol. Res. 2010, 1, 31–35. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, H.; Willard, S.; Wang, Q.J.; Jiang, W.; Zhang, H.X.; Kouba, A. Spatial distribution and seasonal movement patterns of reintroduced Chinese giant salamanders. BMC Zool. 2019, 4, 1–11. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Q.J.; Willard, S.T.; Jiang, W.; Zhang, H.X.; Zhao, H.; Kouba, A.J. Environmental characteristics associated with settlement of reintroduced Chinese giant salamanders. J. Herpetol. 2017, 51, 417–424. [Google Scholar] [CrossRef]
- Vasconcelos, D.; Calhoun, A.J.K. Movement patterns of adult and juvenile Rana sylvatica (LeConte) and Ambystoma maculatum (Shaw) in three restored seasonal pools in Maine. J. Herpetol. 2004, 38, 551–561. [Google Scholar] [CrossRef]
- Bodinof, C.M.; Briggler, J.T.; Junge, R.E.; Beringer, J.; Wanner, M.D.; Schuette, C.D.; Ettling, J.; Gitzen, R.A.; Millspaugh, J.J. Postrelease movements of captive-reared Ozark Hellbenders (Cryptobranchus Alleganiensis Bishopi). Herpetologica 2012, 68, 160–173. [Google Scholar] [CrossRef]
- Walls, S.C.; Barichivich, W.J.; Brown, M.E. Drought, deluge and declines: The impact of precipitation extremes on amphibians in a changing climate. Biology 2013, 2, 399–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trochet, A.; Le Chevalier, H.; Calvez, O.; Barthe, L.; Isselin-Nondedeu, F.; Picard, D.; Debelgarric, M.; Pégourié, N.; Rocher, R.; Ribéron, A. Postbreeding movements in Marbled newts (Caudata, Salamandridae): A comparative radiotracking study in two habitat types. Herpetologica 2017, 73, 1–9. [Google Scholar] [CrossRef]
- Delgado Mdel, M.; Penteriani, V.; Revilla, E.; Nams, V.O. The effect of phenotypic traits and external cues on natal dispersal movements. J. Anim. Ecol. 2010, 79, 620–632. [Google Scholar] [CrossRef]
- Marcec, R.; Kouba, A.; Zhang, L.; Zhang, H.X.; Wang, Q.J.; Zhao, H.; Jiang, W.; Willard, S. Surgical implantation of coelomic radiotransmitters and postoperative survival of Chinese giant salamanders (Andrias davidianus) following reintroduction. J. Zoo Wildl. Med. 2016, 47, 187–195. [Google Scholar] [CrossRef]
- Zhang, L.; Kouba, A.; Wang, Q.J.; Zhao, H.; Jiang, W.; Willard, S.; Zhang, H.X. The effect of water temperature on the growth of captive Chinese giant salamanders (Andrias davidianus) reared for reintroduction: A comparison with wild salamander body condition. Herpetologica 2014, 70, 369–377. [Google Scholar] [CrossRef]
- Xu, W.G.; Wang, Z.Q.; Liang, G. The washing behavior and its significance for male adult Andrias davidianus in the pre-reproductive period. Chin. J. Zool. 2013, 48, 529–533. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop: Release 10; Environmental Systems Research Institute: Redlands, CA, USA, 2011. [Google Scholar]
- Wheeler, B.A.; Prosen, E.; Mathis, A.; Wilkinson, R.F. Population declines of a long-lived salamander: A 20+-year study of hellbenders, Cryptobranchus alleganiensis. Biol. Conserv. 2003, 109, 151–156. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 30 March 2017).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Naimi, B.; Hamm, N.A.S.; Groen, T.A.; Skidmore, A.K.; Toxopeus, A.G. Where is positional uncertainty a problem for species distribution modelling? Ecography 2014, 37, 191–203. [Google Scholar] [CrossRef]
- Zimecki, M. The lunar cycle: Effects on human and animal behavior and physiology. Postepy Hig. Med. Dosw. 2006, 60, 1–7. [Google Scholar]
- Grant, R.; Halliday, T.; Chadwick, E. Amphibians’ response to the lunar synodic cycle—a review of current knowledge, recommendations, and implications for conservation. Behav. Ecol. 2013, 24, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Lima, S.L.; Dill, L.M. Behavioral decisions made under the risk of predation—A review and prospectus. Can. J. Zool. 1990, 68, 619–640. [Google Scholar] [CrossRef]
- Penteriani, V.; Kuparinen, A.; Delgado, M.d.M.; Lourenço, R.; Campioni, L. Individual status, foraging effort and need for conspicuousness shape behavioural responses of a predator to moon phases. Anim. Behav. 2011, 82, 413–420. [Google Scholar] [CrossRef]
- Cozzi, G.; Broekhuis, F.; McNutt, J.W.; Turnbull, L.A.; Macdonald, D.W.; Schmid, B. Fear of the dark or dinner by moonlight? Reduced temporal partitioning among Africa’s large carnivores. Ecology 2012, 93, 2590–2599. [Google Scholar] [CrossRef]
- Elangovan, V.; Marimuthu, G. Effect of moonlight on the foraging behaviour of a megachiropteran bat Cynopterus sphinx. J. Zool. 2001, 253, 347–350. [Google Scholar] [CrossRef]
- Penteriani, V.; Kuparinen, A.; del Mar Delgado, M.; Palomares, F.; Lopez-Bao, J.V.; Fedriani, J.M.; Calzada, J.; Moreno, S.; Villafuerte, R.; Campioni, L.; et al. Responses of a top and a meso predator and their prey to moon phases. Oecologia 2013, 173, 753–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broekhuis, F.; Grünewälder, S.; McNutt, J.W.; Macdonald, D.W. Optimal hunting conditions drive circalunar behavior of a diurnal carnivore. Behav. Ecol. 2014, 25, 1268–1275. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wang, Q.Y.; Yang, L.; Li, F.; Chan, B.P.L.; Xiao, Z.S.; Li, S.; Song, D.Z.; Piao, Z.J.; Fan, P.F. The neglected otters in China: Distribution change in the past 400 years and current conservation status. Biol. Conserv. 2018, 228, 259–267. [Google Scholar] [CrossRef]
- Bond, D.J.; Avery, R.A. Movement patterns of lacertid lizards: Effects of temperature on speed, pauses and gait in Lacerta vivipara. Amphib. Reptil. 1989, 10, 77–84. [Google Scholar]
- Lucas, M.C.; Batley, E. Seasonal movements and behaviour of adult barbel Barbus barbus, a riverine cyprinid fish: Implications for river management. J. Appl. Ecol. 1996, 33, 1345–1358. [Google Scholar] [CrossRef]
- Hurlbert, A.H.; Ballantyne, F.; Powell, S. Shaking a leg and hot to trot: The effects of body size and temperature on running speed in ants. Ecol. Entomol. 2008, 33, 144–154. [Google Scholar] [CrossRef]
- Gibert, J.P.; Chelini, M.C.; Rosenthal, M.F.; DeLong, J.P. Crossing regimes of temperature dependence in animal movement. Glob. Chang. Biol. 2016, 22, 1722–1736. [Google Scholar] [CrossRef]
- Liu, J.Y.; Tan, Y.A.; Tan, Q.S.; He, X.L.; Zhang, Y.C.; Liu, M.G. Research on Chinese giant salamander F2 adaptability and growth advantages. Sichuan J. Zool. 2006, 25, 387–390. (In Chinese) [Google Scholar]
- Cong, W.; Zhou, Y.L.; Zhu, Q.H.; Zhou, Z.K.; Gu, W.B.; Liu, Z.P.; Wang, L.Z.; Shu, M.A. Effects of heat stress on the liver of the Chinese giant salamander Andrias davidianus: Histopathological changes and expression characterization of Nrf2-mediated antioxidant pathway genes. J. Ther. Biol. 2018, 76, 115–125. [Google Scholar]
- Timm, B.C.; McGarigal, K.; Compton, B.W. Timing of large movement events of pond-breeding amphibians in Western Massachusetts, USA. Biol. Conserv. 2007, 136, 442–454. [Google Scholar] [CrossRef]
- Taylor, M.K.; Cooke, S.J. Meta-analyses of the effects of river flow on fish movement and activity. Environ. Rev. 2012, 20, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Lowe, W.H. Linking dispersal to local population dynamics: A case study using a headwater salamander system. Ecology 2003, 84, 2145–2154. [Google Scholar] [CrossRef]
- Cecala, K.K.; Price, S.J.; Dorcas, M.E. Evaluating existing movement hypotheses in linear systems using larval stream salamanders. Can. J. Zool. 2009, 87, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Schulte-Hostedde, A.I.; Zinner, B.; Millar, J.S.; Hickling, G.J. Restitution of Mass–Size Residuals: Validating Body Condition Indices. Ecology 2005, 86, 155–163. [Google Scholar] [CrossRef]
- O’Riain, M.J.; Jarvis, J.; Faulkes, C.G. A dispersive morph in the Naked Mole-rat. Nature 1996, 380, 619–621. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| River | Hypothesis | Model Structure | K | AIC | ∆AIC |
|---|---|---|---|---|---|
| Donghe | Positive effect of moon phase | ~moon + (1|ID) | 3 | 4746.9 | 0 |
| Null | ~(1|ID) | 2 | 4751.8 | 4.9 | |
| Positive effect of body condition | ~condition + (1|ID) | 3 | 4751.9 | 5 | |
| Positive effect of precipitation | ~precipitation + (1|ID) | 3 | 4752.0 | 5.1 | |
| Positive effect of precipitation in the previous day | ~precipitationpre + (1|ID) | 3 | 4752.8 | 5.9 | |
| Positive effect of body mass | ~mass + (1|ID) | 3 | 4753.6 | 6.7 | |
| Positive effect of temperature | ~temperature + (1|ID) | 3 | 4753.7 | 6.8 | |
| Heihe | Null | ~(1|ID) | 2 | 1514.5 | 0 |
| Positive effect of temperature | ~temperature + (1|ID) | 3 | 1515.2 | 0.7 | |
| Positive effect of moon phase | ~moon + (1|ID) | 3 | 1515.4 | 0.9 | |
| Positive effect of precipitation in the previous day | ~precipitationpre + (1|ID) | 3 | 1516.5 | 2 | |
| Positive effect of precipitation | ~precipitation + (1|ID) | 3 | 1516.5 | 2 | |
| Positive effect of body mass | ~mass + (1|ID) | 3 | 1516.5 | 2 | |
| Positive effect of body condition | ~condition + (1|ID) | 3 | 1516.5 | 2 |
| River | Hypothesis | Model Structure | K | AIC | ∆AIC |
|---|---|---|---|---|---|
| Donghe | Positive effect of temperature | ~temperature + (1|ID) | 4 | 22,098 | 0 |
| Positive effect of temperature and precipitation | ~temperature + precipitation + (1|ID) | 5 | 22,099 | 1 | |
| Positive effect of precipitation | ~precipitation + (1|ID) | 4 | 22,127 | 29 | |
| Null | ~+ (1|ID) | 3 | 22,129 | 31 | |
| Positive effect of moon phase | ~moon + (1|ID) | 4 | 22,129 | 31 | |
| Positive effect of precipitation in the previous day | ~precipitationpre + (1|ID) | 4 | 22,129 | 31 | |
| Positive effect of body mass | ~mass + (1|ID) | 4 | 22,131 | 33 | |
| Positive effect of body condition | ~condition + (1|ID) | 4 | 22,131 | 33 | |
| Heihe | Positive effect of temperature | ~temperature + (1|ID) | 4 | 7921.5 | 0 |
| Positive effect of temperature and precipitation in the previous day | ~temperature + precipitationpre + (1|ID) | 5 | 7921.5 | 0 | |
| Positive effect of precipitation in the previous day | ~precipitationpre + (1|ID) | 4 | 7928.6 | 7.1 | |
| Null | ~(1|ID) | 3 | 7928.7 | 7.2 | |
| Positive effect of body condition | ~condition + (1|ID) | 4 | 7928.8 | 7.3 | |
| Positive effect of moon phase | ~moon + (1|ID) | 4 | 7929.2 | 7.7 | |
| Positive effect of precipitation | ~precipitation + (1|ID) | 4 | 7929.7 | 8.2 | |
| Positive effect of body mass | ~mass + (1|ID) | 4 | 7930.6 | 9.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Zhang, L.; Zhao, H.; Zhao, Q.; Deng, J.; Kong, F.; Jiang, W.; Zhang, H.; Liu, H.; Kouba, A. Abiotic and Biotic Influences on the Movement of Reintroduced Chinese Giant Salamanders (Andrias davidianus) in Two Montane Rivers. Animals 2021, 11, 1480. https://doi.org/10.3390/ani11061480
Wang Q, Zhang L, Zhao H, Zhao Q, Deng J, Kong F, Jiang W, Zhang H, Liu H, Kouba A. Abiotic and Biotic Influences on the Movement of Reintroduced Chinese Giant Salamanders (Andrias davidianus) in Two Montane Rivers. Animals. 2021; 11(6):1480. https://doi.org/10.3390/ani11061480
Chicago/Turabian StyleWang, Qijun, Lu Zhang, Hu Zhao, Qing Zhao, Jie Deng, Fei Kong, Wei Jiang, Hongxing Zhang, Hong Liu, and Andrew Kouba. 2021. "Abiotic and Biotic Influences on the Movement of Reintroduced Chinese Giant Salamanders (Andrias davidianus) in Two Montane Rivers" Animals 11, no. 6: 1480. https://doi.org/10.3390/ani11061480