
Association between Anti-Müllerian Hormone Concentration and Inflammation Markers in Serum during the Peripartum Period in Dairy Cows

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Management
2.2. Blood Sample Collection and Analysis
2.3. Evaluation of Ovary and Uterine Condition
2.4. Somatic Cell Count of Milk
2.5. Reproductive Records
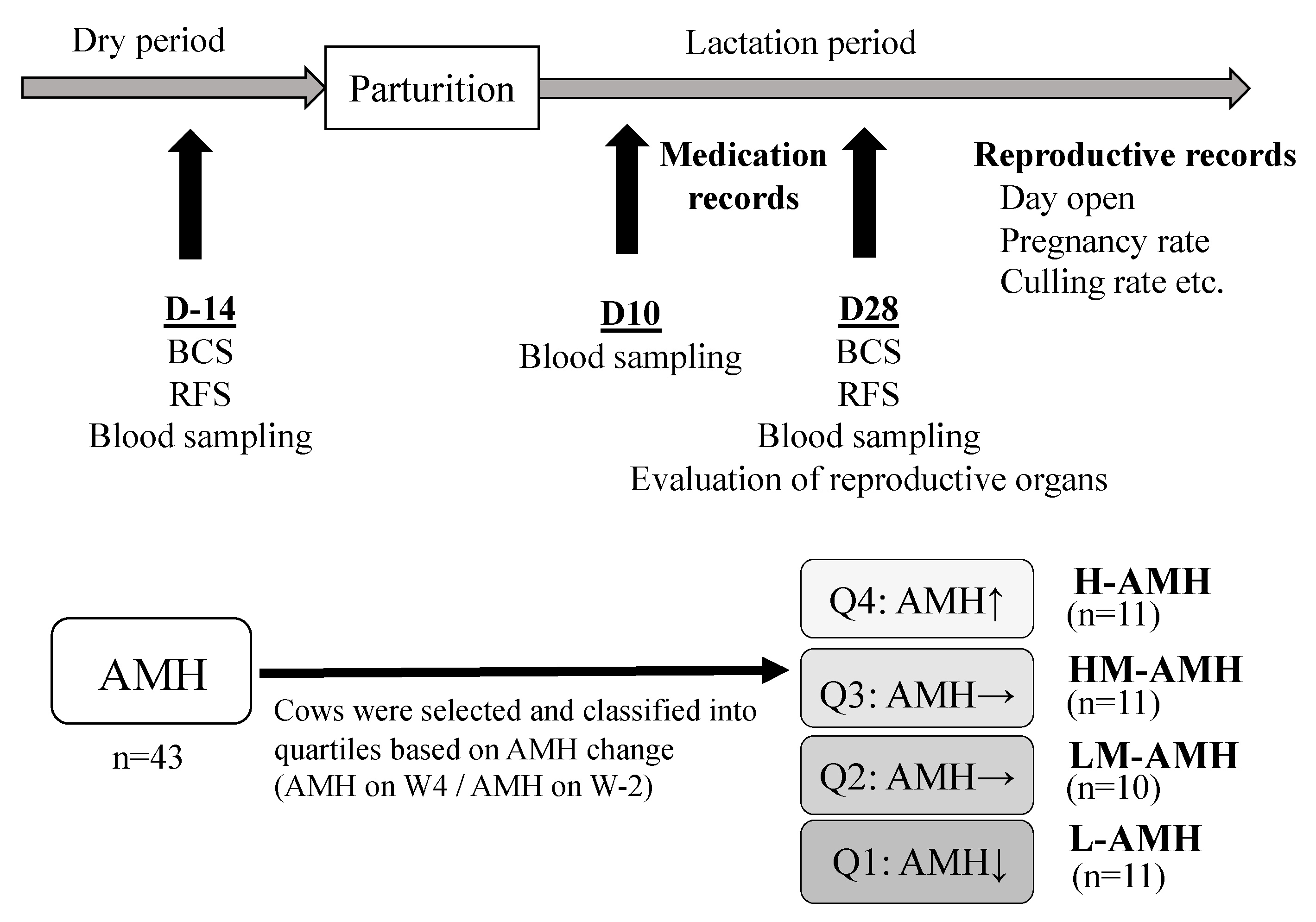
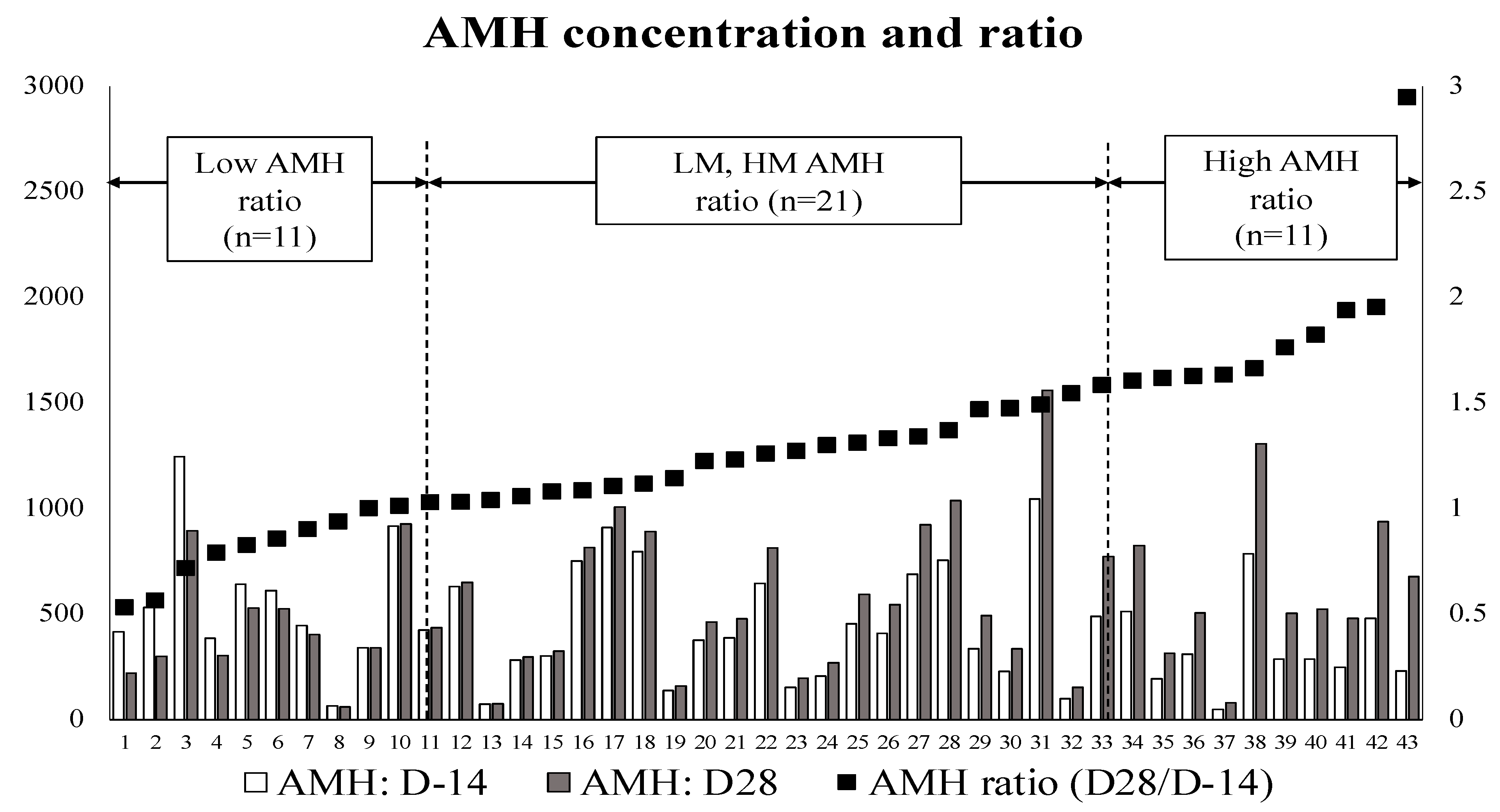
2.6. Data Management and Classification of the Cows Based on Their AMH Concentration
2.7. Statistical Analysis
3. Results
3.1. First Evaluation: Relationship between the AMH Ratio (AMH at D28/AMH at D-14) with Serum Metabolic and Inflammation Parameters
3.2. Second Evaluation: AMH Ratio and Serum Biochemical Markers
3.3. Second Evaluation: AMH Ratio, Clinical Status of Mammary Gland and Reproductive Organs, and Reproductive Records
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gobikrushanth, M.; Purfield, D.C.; Colazo, M.G.; Butler, S.T.; Wang, Z.; Ambrose, D.J. The relationship between serum anti-Mullerian hormone concentrations and fertility, and genome-wide associations for anti-Mullerian hormone in Holstein cows. J. Dairy Sci. 2018, 101, 7563–7574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawaz, M.Y.; Jimenez-Krassel, F.; Steibel, J.P.; Lu, Y.; Baktula, A.; Vukasinovic, N.; Neuder, L.; Ireland, J.L.H.; Ireland, J.J.; Tempelman, R.J. Genomic heritability and genome-wide association analysis of anti-Mullerian hormone in Holstein dairy heifers. J. Dairy Sci. 2018, 101, 1–13. [Google Scholar] [CrossRef]
- Krause, A.R.T.; Pfeifer, L.F.; Montagner, P.; Weschenfelder, M.M.; Schwegler, E.; Lima, M.E.; Xavier, E.G.; Brauner, C.C.; Schmitt, E.; Del Pino, F.A.B.; et al. Associations between resumption of postpartum ovarian activity, uterine health and concentrations of metabolites and acute phase proteins during the transition period in Holstein cows. Anim. Reprod. Sci. 2014, 145, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Meikle, A.; de Brun, V.; Carriquiry, M.; Soca, P.; Sosa, C.; de Lourdes Adrien, M.; Chilibroste, P.; Abecia, J.A. Influences of nutrition and metabolism on reproduction of the female ruminant. Anim. Reprod. 2018, 15, 899–911. [Google Scholar] [CrossRef]
- Okawa, H.; Fujikura, A.; Wijayagunawardane, M.M.P.; Vos, P.L.A.M.; Taniguchi, M.; Takagi, M. Effect of diagnosis and treatment of clinical endometritis based on vaginal discharge score grading system in postpartum Holstein cows. J. Vet. Med. Sci. 2017, 79, 1545–1551. [Google Scholar] [CrossRef] [Green Version]
- Okawa, H.; Goto, A.; Wijayagunawardane, M.M.P.; Vos, P.L.A.M.; Yamato, O.; Taniguchi, M.; Takagi, M. Risk factors associated with reproductive performance in Japanese dairy cows: Vaginal discharge with flecks of pus or calving abnormality extend time to pregnancy. J. Vet. Med. Sci. 2019, 81, 95–99. [Google Scholar] [CrossRef] [Green Version]
- Chastant, S.; Saint-Dizier, M. Inflammation: Friend or foe of bovine reproduction? Anim. Reprod. 2019, 16, 539–547. [Google Scholar] [CrossRef] [Green Version]
- Cheong, S.H.; Sa Filho, O.G.; Absalon-Medina, V.A.; Schneider, A.; Butler, W.R.; Gilbert, R.O. Uterine and systemic inflammation influences ovarian follicular function in postpartum dairy cows. PLoS ONE 2017, 12, e0177356. [Google Scholar] [CrossRef] [Green Version]
- Sheldon, I.M.; Owens, S.E.; Turner, M.L. Innate immunity and the sensing of infection, damage and danger in the female genital tract. J. Reprod. Immunol. 2017, 119, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, E.S.; Carvalho, M.R. Impact and mechanisms of inflammatory diseases on embryonic development and fertility in cattle. Anim. Reprod. 2017, 14, 589–600. [Google Scholar] [CrossRef]
- Monniaux, D.; di Clemente, N.; Touzé, J.L.; Belville, C.; Rico, C.; Bontoux, M.; Picard, J.Y.; Fabre, S. Intrafollicular steroids and anti-mullerian hormone during normal and cystic ovarian follicular development in the cow. Biol. Reprod. 2008, 79, 387–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monniaux, D.; Drouilhet, L.; Rico, C.; Estienne, A.; Jarrier, P.; Touze, J.L.; Sapa, J.; Phocas, F.; Dupont, J.; Dalbies-Tran, R.; et al. Regulation of anti-Mullerian hormone production in domestic animals. Reprod. Fertil. Dev. 2013, 25, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Monniaux, D.; Barbey, S.; Rico, C.; Fabre, S.; Gallard, Y.; Larroque, H. Anti-Müllerian hormone: A predictive marker of embryo production in cattle? Reprod. Fertil. Dev. 2010, 22, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Ireland, J.J.; Smith, G.W.; Scheetz, D.; Jimenez-Krassel, F.; Folger, J.K.; Ireland, J.L.; Mossa, F.; Lonergan, P.; Evans, A.C. Does size matter in females? An overview of the impact of the high variation in the ovarian reserve on ovarian function and fertility, utility of anti-Müllerian hormone as a diagnostic marker for fertility and causes of variation in the ovarian reserve in cattle. Reprod. Fertil. Dev. 2011, 23, 1–14. [Google Scholar] [CrossRef]
- Mossa, F.; Ireland, J.J. Physiology and endocrinology symposium: Anti-Mullerian hormone: A biomarker for the ovarian reserve, ovarian function, and fertility in dairy cows. J. Anim. Sci. 2019, 97, 1446–1455. [Google Scholar] [CrossRef]
- Fushimi, Y.; Monniaux, D.; Takagi, M. Efficacy of a single measurement of plasma anti-Müllerian hormone concentration for ovum pick-up donor selection of Japanese Black heifers in herd breeding programs. J. Reprod. Dev. 2019, 65, 369–374. [Google Scholar] [CrossRef] [Green Version]
- Webb, R.; Garnsworthy, P.C.; Gong, J.-G.; Armstrong, D.G. Control of follicular growth: Local interactions and nutritional influences. J. Anim. Sci. 2004, 82, E63–E74. [Google Scholar] [CrossRef]
- Webb, R.; Garnsworthy, P.C.; Campbell, B.K.; Hunter, M.G. Intra-ovarian regulation of follicular development and oocyte competence in farm animals. Theriogenology 2007, 68, S22–S29. [Google Scholar] [CrossRef] [PubMed]
- Scaramuzzi, R.J.; Brown, H.M.; Dupont, J. Nutritional and metabolic mechanisms in the ovary and their role in mediating the effects of diet on folliculogenesis: A perspective. Reprod. Dom. Anim. 2010, 45, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, I.M.; Cronin, J.G.; Healey, G.D.; Gabler, C.; Heuwieser, W.; Streyl, D.; Bromfield, J.J.; Miyamato, A.; Fergani, C.; Dobson, H. Innate immunity and inflammation of the bovine female reproductive tract in health and disease. Reproduction 2014, 148, R41–R51. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, R.O. Symposium review: Mechanisms of disruption of fertility by infectious diseases of the reproductive tract. J. Dairy Sci. 2019, 102, 3754–3765. [Google Scholar] [CrossRef] [Green Version]
- Berg, L.C.; Thomsen, P.D.; Andersen, P.H.; Jensen, H.E.; Jacobsen, S. Serum amyloid A is expressed in histologically normal tissues from horses and cattle. Vet. Immunol. Immunopathol. 2011, 144, 155–159. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, F.; Oguejiofor, C.F.; Wang, D.; Dong, S.; Yan, Z. Endometrial expression of the acute phase molecule SAA is more significant than HP in reflecting the severity of endometritis. Res. Vet. Sci. 2018, 121, 130–133. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, J.D.; Galligan, D.T.; Thomsen, N. Principal descriptors of body condition score in Holstein cows. J. Dairy Sci. 1994, 77, 2695–2703. [Google Scholar] [CrossRef]
- Kawashima, C.; Karaki, C.; Munakata, M.; Matsui, M.; Shimizu, T.; Miyamoto, A.; Kida, K. Association of rumen fill score and energy status during the close-up dry period with conception at first artificial insemination in dairy cows. Anim. Sci. J. 2016, 87, 1218–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, U.; Okamoto, K.; Miyamoto, A.; Otoi, T.; Yamato, O.; Tshering, C.; Takagi, M. A Japanese Black breeding herd exhibiting low blood urea nitrogen: A metabolic profile study examining the effect on reproductive performance. Anim. Sci. J. 2013, 84, 389–394. [Google Scholar] [CrossRef]
- Kawashima, C.; Sakaguchi, M.; Suzuki, T.; Sasamoto, Y.; Takahashi, Y.; Matsui, M.; Miyamoto, A. Metabolic profile in ovulatory and anovulatory primiparous dairy cows during the first follicular wave postpartum. J. Reprod. Dev. 2007, 53, 113–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takagi, M.; Yamagishi, N.; Lee, I.H.; Oboshi, K.; Tsuno, M.; Wijayagunawardane, M.P.B. Reproductive management with ultrasound scanner-monitoring system for a high-yielding commercial dairy herd reared under stanchion management style. Asian-Aust. J. Anim. Sci. 2005, 18, 949–956. [Google Scholar] [CrossRef]
- Mido, S.; Murata, N.; Rawy, M.S.; Kitahara, G.; Osawa, T. Effects of intrauterine infusion of povidone-iodine on endometrial cytology and bacteriology in dairy cows with clinical endometritis. J. Vet. Med. Sci. 2016, 78, 551–556. [Google Scholar] [CrossRef] [Green Version]
- Lukas, J.M.; Hawkins, D.M.; Kinsel, M.L.; Reneau, J.K. Bulk tank somatic cell cpunts analyzed by statistical process control tools to identified and monitor subclinical mastitis incidence. J. Dairy Sci. 2005, 88, 3944–3952. [Google Scholar] [CrossRef]
- Santos, G.; Bottino, M.P.; Santos, A.P.C.; Simoes, L.M.S.; Souza, J.C.; Ferreira, M.B.D.; da Silveira, J.C.; Avila, A.C.F.C.M.; Bride, A.; Sales, J.N.S. Subclinical mastitis interferes with ovulation, oocyte and granulosa cell quality in dairy cows. Theriogenology 2018, 119, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.E.P.; Rutigliano, H.M.; Sa Filho, M.F. Risk factors for resumption of postpartum cyclicity and embryonic survival in lactating dairy cows. Anim. Reprod. Sci. 2009, 110, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, E.S.; Gomes, G.; Greco, L.F.; Cerri, R.L.A.; Vieira-Neto, A.; Monteiro Jr, P.L.J.; Lima, F.S.; Bisinotto, R.S.; Thatcher, W.W.; Santos, J.E.P. Carryover effect of postpartum inflammatory diseases on developmental biology and fertility in lactating dairy cows. J. Dairy Sci. 2016, 99, 2201–2220. [Google Scholar] [CrossRef] [PubMed]
- Mossa, F.; Jimenez-Krassel, F.; Scheetz, D.; Weber-Nielsen, M.; Evans, A.C.O.; Ireland, J.J. Anti-Mullerian Hormone (AMH) and fertility management in agricultural species. Reproduction 2017, 154, R1–R11. [Google Scholar] [CrossRef] [PubMed]
- Umer, S.; Zhao, S.J.; Sammad, A.; Sahlu, B.W.; Yunwei, P.; Zhu, H. AMH: Could it be used as a biomarker for fertility and superovulation in domestic animals. Genes 2019, 10, 1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rico, C.; Drouilhet, L.; Salvetti, P.; Dalbiès-Tran, R.; Jarrier, P.; Touzé, J.L.; Pillet, E.; Ponsart, C.; Fabre, S.; Monniaux, D. Determination of anti-Müllerian hormone concentrations in blood as a tool to select Holstein donor cows for embryo production: From the laboratory to the farm. Reprod. Fertil. Dev. 2012, 24, 932–944. [Google Scholar] [CrossRef]
- Gamarra, G.; Ponsart, C.; Lacaze, S.; Le Guienne, B.; Humblot, P.; Deloche, M.C.; Monniaux, D.; Ponter, A.A. Dietary propylene glycol and in vitro embryo production after ovum pick-up in heifers with different anti-Müllerian hormone profiles. Reprod. Fertil. Dev. 2014, 27, 1249–1261. [Google Scholar] [CrossRef]
- Puppel, K.; Kuczynska, B. Metabolic profiles of cow’s blood; a review. J. Sci. Food Agric. 2016, 96, 4321–4328. [Google Scholar] [CrossRef]
- Deelen, S.M.; Leslie, K.E.; Steele, M.A.; Eckert, E.; Brown, H.E.; DeVriest, T.J. Validation of a calf-side-β-hydroxybutyrate test and its utility for estimation of starter intake in dairy calves around weaning. J. Dairy Sci. 2016, 99, 7624–7633. [Google Scholar] [CrossRef] [Green Version]
- Raboisson, D.; Mounié, M.; Maigné, E. Diseases, reproductive performance, and changes in milk production associated with subclinical ketosis in dairy cows: A meta-analysis and review. J. Dairy Sci. 2014, 97, 7547–7563. [Google Scholar] [CrossRef]
- Burke, C.R.; Meier, S.; McDougall, S.; Compton, C.; Mitchell, M.; Roche, J.R. Relationships between endometritis and metabolic state during the transition period in pasture-grazed dairy cows. J. Dairy Sci. 2010, 93, 5363–5373. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, J.J.; Harvey, J.W.; Bruss, M.L. Chapter 5- Proteins, Proteomics, and the Dysproteinemias. In Clinical Biochemistry of Domestic Animals, 6th ed.; Academic Press: San Diego, CA, USA, 2008; pp. 117–155. [Google Scholar]
- Murata, H.; Shimada, N.; Yoshioka, M. Current research on acute phase proteins in veterinary diagnosis: An overview. Vet. J. 2004, 168, 28–40. [Google Scholar] [CrossRef]
- Schneider, A.; Correa, M.N.; Butler, W.R. Acute phase proteins in Holstein cows diagnosed with uterine infection. Res. Vet. Sci. 2013, 95, 269–271. [Google Scholar] [CrossRef] [PubMed]
- Johns, J.L.; Heller, M. Chapter 26. Alterations in blood proteins. In Large Animal Internal Medicine, 6th ed.; Smith, B.P., Ed.; Elsevier: St. Louis, MO, USA, 2020; pp. 425–441. [Google Scholar]
- Monniaux, D. Factors influencing establishment of the ovarian reserve and their effects on fertility. Anim. Reprod. 2018, 15, 635–647. [Google Scholar] [CrossRef]
- Nielsen, F.H. Effects of magnesium depletion on inflammation in chronic disease. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 525–530. [Google Scholar] [CrossRef]
- Monniaux, D.; Clément, F.; Dalbiès-Tran, R.; Estienne, A.; Fabre, S.; Mansanet, C.; Monget, P. The ovarian reserve of primordial follicles and the dynamic reserve of antral growing follicles: What is the link? Biol. Reprod. 2014, 90, 85. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | D-14 | D10 | D28 | |||
|---|---|---|---|---|---|---|
| r | p | r | p | r | p | |
| TP | −0.4102 | 0.0063 | −0.0597 | 0.7039 | −0.3305 | 0.0304 |
| A/G ratio | 0.231 | 0.1362 | 0.3877 | 0.0102 | 0.435 | 0.0036 |
| Mg | 0.0872 | 0.5783 | 0.5512 | <0.001 | 0.3958 | 0.0086 |
| SAA | −0.0344 | 0.8265 | −0.3161 | 0.039 | −0.2880 | 0.0611 |
| NEFA | 0.1679 | 0.2819 | 0.414 | 0.0058 | 0.4034 | 0.0073 |
| Prepartum (D-14) | Postpartum (D28) | AMH Ratio | |
|---|---|---|---|
| AMH | AMH | (D28/D-14) | |
| Q1: L-AMH (n = 11) | 547.9 ± 94.5 | 449.6 ± 79.7 | 0.83 ± 0.05 |
| Q2: LM-AMH (n = 10) | 465.1 ± 91.3 | 516.4 ± 99.9 | 1.11 ± 0.02 |
| Q3: HM-AMH (n = 11) | 456.7 ± 89.4 | 629.4 ± 128.3 | 1.38 ± 0.03 |
| Q4: H-AMH (n = 11) | 351.9 ± 60.4 | 629.7 ± 99.1 | 1.83 ± 0.12 |
| Prepartum (D-14) | Postpartum (D28) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H-AMH | L-AMH | H-AMH | L-AMH | |||||||||
| BCS | 3.2 | ± | 0.1 c | 3.4 | ± | 0.1 d | 2.8 | ± | 0.1 | 2.9 | ± | 0.1 |
| RFS | 3.5 | ± | 0.2 | 3.9 | ± | 0.2 | 3.1 | ± | 0.2 | 3.0 | ± | 0.3 |
| AST (U/L) | 59.2 | ± | 3.5 | 52.6 | ± | 3.5 | 73.7 | ± | 5.1 | 64.7 | ± | 4.8 |
| GGT (U/L) | 17.2 | ± | 1.9 | 15.7 | ± | 1.1 | 22.3 | ± | 1.1 | 29.9 | ± | 7.9 |
| BUN (mg/dL) | 8.3 | ± | 0.6 | 8.2 | ± | 0.4 | 10.2 | ± | 0.8 | 10.1 | ± | 1.1 |
| Bil (mg/dL) | 0.18 | ± | 0.02 | 0.13 | ± | 0.01 | 0.19 | ± | 0.01 | 0.15 | ± | 0.02 |
| NEFA (mmol/L) | 169.2 | ± | 29.4 | 120.6 | ± | 11.0 | 325.0 | ± | 41.6 | 241.6 | ± | 31.9 |
| T-Cho (mg/dL) | 69.7 | ± | 4.0 | 76.2 | ± | 4.3 | 172.7 | ± | 10.5 | 155.7 | ± | 8.4 |
| Glu (mg/dL) | 56.4 | ± | 4.1 | 62.9 | ± | 3.2 | 48.2 | ± | 3.3 | 57.6 | ± | 4.7 |
| TG (mg/dL) | 15.7 | ± | 0.8 | 17.2 | ± | 1.4 | 5.5 | ± | 0.3 | 5.9 | ± | 0.3 |
| Ca (mg/dL) | 9.3 | ± | 0.1 | 9.4 | ± | 0.1 | 9.5 | ± | 0.2 | 9.3 | ± | 0.1 |
| iP (mg/dL) | 5.9 | ± | 0.3 | 5.8 | ± | 0.2 | 5.7 | ± | 0.2 | 5.4 | ± | 0.3 |
| Mg (mg/dL) | 2.4 | ± | 0.1 | 2.3 | ± | 0.04 | 2.6 | ± | 0.1 a | 2.3 | ± | 0.1 b |
| Fe (μg/dL) | 170.1 | ± | 9.5 | 151.6 | ± | 9.2 | 125.5 | ± | 6.7 | 101.6 | ± | 12.4 |
| 3-HB (μmol/dL) | 643.7 | ± | 25.3 | 645.5 | ± | 47.5 | 1504.4 | ± | 229.5 a | 844.5 | ± | 184.2 b |
| TP (mg/dL) | 7.0 | ± | 0.2 a | 7.6 | ± | 0.2 b | 7.5 | ± | 0.1 a | 8.1 | ± | 0.2 b |
| Alb (mg/dL) | 3.7 | ± | 0.1 | 3.7 | ± | 0.1 | 4.0 | ± | 0.1 a | 3.6 | ± | 0.1 b |
| A/G ratio | 1.15 | ± | 0.07 | 1.01 | ± | 0.09 | 1.14 | ± | 0.06 a | 0.84 | ± | 0.06 b |
| IGF1 (ng/mL) | 31.8 | ± | 6.4 | 37.9 | ± | 5.9 | 42.3 | ± | 5.4 | 39.3 | ± | 4.9 |
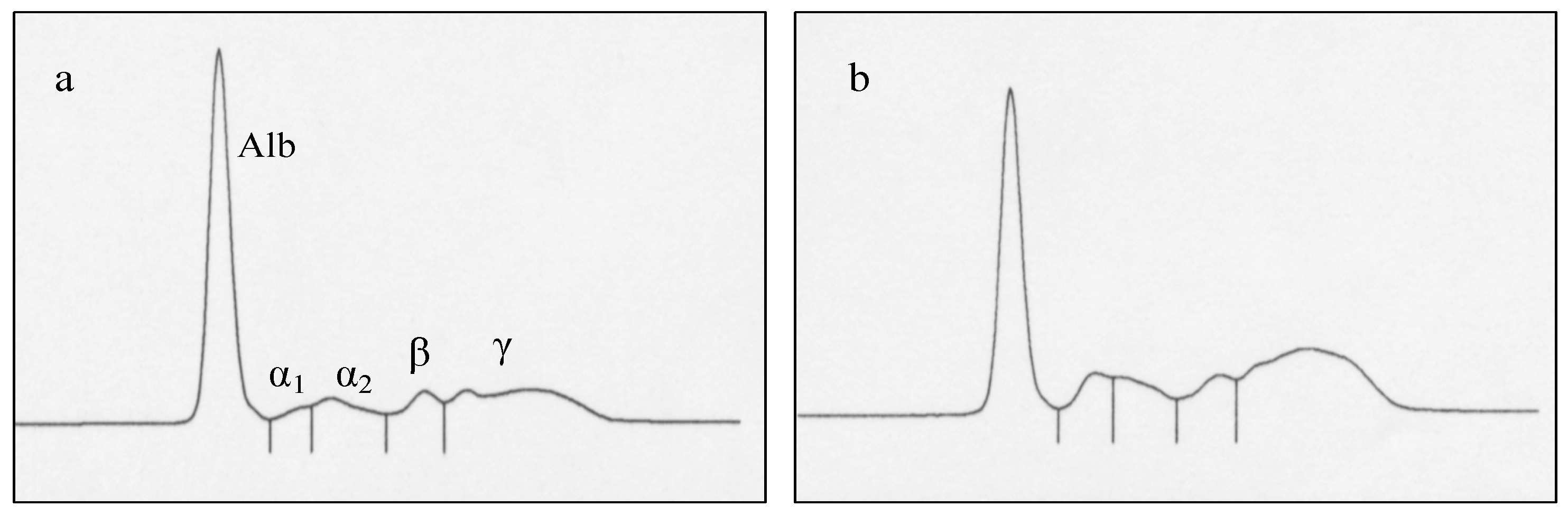
| Prepartum (D-14) | Postpartum (D28) | |||||||
|---|---|---|---|---|---|---|---|---|
| H-AMH Group | L-AMH Group | H-AMH Group | L-AMH Group | |||||
| Total protein (mg/dL) (1) | 7.1 | 6.4–7.9 * | 7.6 | 6.9–8.4 * | 7.6 | 7.2–8.0 # | 8.1 | 7.7–8.5 # |
| Albumin | 3.6 | 3.0–4.2 | 3.6 | 3.1–4.2 | 3.9 | 3.4–4.3 ## | 3.5 | 3.0–4.0 ## |
| α1-globulin | 0.23 | 0.20–0.26 ** | 0.26 | 0.24–0.29 ** | 0.29 | 0.21–0.37 | 0.33 | 0.25–0.41 |
| α2-globulin | 0.60 | 0.52–0.69 | 0.62 | 0.53–0.71 | 0.64 | 0.59–0.69 | 0.66 | 0.61–0.71 |
| β-globulin | 0.67 | 0.54–0.81 | 0.83 | 0.69–0.96 | 0.62 | 0.48–0.76 ### | 0.81 | 0.67–0.94 ### |
| γ-globulin | 2.2 | 1.6–2.8 | 2.2 | 1.6–2.9 | 2.0 | 1.5–2.6 #### | 2.6 | 2.1–3.2 #### |
| SAA (μg/mL) | 10.9 | 0.0–35.0 | 10.9 | 0.0–35.0 | 3.5 | 0.0–17.3 ##### | 22.4 | 9.5–35.3 ##### |
| H-AMH | L-AMH | |||
|---|---|---|---|---|
| SCC (/μL) | 143.1 | [0.0–1069.9] | 728.4 | [0.0–1632.1] |
| (DIM: 35.3 [29.4–41.3]) * | (DIM: 30.5 [24.6–36.5]) * | |||
| PMNL (%) | 15.9 | [2.8–29.0] | 17.8 | [4.7–30.9] |
| VDS | 0.63 | [0.04–1.22] | 0.56 | [0.05–1.06] |
| CL rate (%) | 27.0 [0.0–59.0] (3/11) | 45.8 [13.7–77.8] (5/11) | ||
| No. of Medication Treatment | Calving to FAI Interval (Days) | Pregnancy Rate of FAI | Days Open | No. of AI | No. of Pregnant Cows within 200 DIM | Culling Rate | |
|---|---|---|---|---|---|---|---|
| H-AMH (n = 11) | 0.8 [0.0–3.2] * (n = 3) | 105.7 [84.4–127.0] | 5/11 50.0% [11.8–88.3] | 147.1 [114.3–179.9] | 1.9 [0.3–3.4] | 9/11 ** 78.2% [42.8–100.0] | 1/11 *** 20.7% [0.0–56.8] |
| L-AMH (n = 11) | 3.9 [1.5–6.3] * (n = 9) | 89.8 [64.8–114.7] | 3/8 # 59.9% [5.7–100.0] | 133.3 [86.8–179.7] | 1.1 [0.0–2.8] | 4/11 ** 35.8% [1.8–69.9] | 6/11 *** 58.9% [23.0–94.7] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okawa, H.; Monniaux, D.; Mizokami, C.; Fujikura, A.; Takano, T.; Sato, S.; Shinya, U.; Kawashima, C.; Yamato, O.; Fushimi, Y.; et al. Association between Anti-Müllerian Hormone Concentration and Inflammation Markers in Serum during the Peripartum Period in Dairy Cows. Animals 2021, 11, 1241. https://doi.org/10.3390/ani11051241
Okawa H, Monniaux D, Mizokami C, Fujikura A, Takano T, Sato S, Shinya U, Kawashima C, Yamato O, Fushimi Y, et al. Association between Anti-Müllerian Hormone Concentration and Inflammation Markers in Serum during the Peripartum Period in Dairy Cows. Animals. 2021; 11(5):1241. https://doi.org/10.3390/ani11051241
Chicago/Turabian StyleOkawa, Hiroaki, Danielle Monniaux, Chihiro Mizokami, Atsushi Fujikura, Toshihiro Takano, Satoko Sato, Urara Shinya, Chiho Kawashima, Osamu Yamato, Yasuo Fushimi, and et al. 2021. "Association between Anti-Müllerian Hormone Concentration and Inflammation Markers in Serum during the Peripartum Period in Dairy Cows" Animals 11, no. 5: 1241. https://doi.org/10.3390/ani11051241