
Diseases Caused by Amoebae in Fish: An Overview


Departament de Biologia Animal, de Biologia Vegetal i d’Ecologia and Servei de Diagnòstic Patològic en Peixos, Universitat Autònoma de Barcelona, 08193 Barcelona, Spain
*
Authors to whom correspondence should be addressed.
†
Both authors contributed equally to this work.
Animals 2021, 11(4), 991; https://doi.org/10.3390/ani11040991
Submission received: 2 March 2021
/
Revised: 28 March 2021
/
Accepted: 29 March 2021
/
Published: 1 April 2021
(This article belongs to the Special Issue Fish Pathology)
{kind=link}
{kind=link}
Abstract
:Simple Summary
Amoebae can be found in many different aquatic environments and are also an emerging risk for fish health. Amoebae can display different types of relationships with fish, some of them (amoeba acting as commensals) do not harm fish. However, in many cases they can act as parasites and can be the cause of severe diseases affecting mainly the gills and also causing relevant systemic infections.
Abstract
Parasitic and amphizoic amoebae are ubiquitous and can affect a huge variety of hosts, from invertebrates to humans, and fish are not an exception. Most of the relationships between amoebae and fish are based on four different types: ectocommensals, ectoparasites, endocommensals and endoparasites, although the lines between them are not always clear. As ectocommensals, they are located specially on the gills and particularly the amphizoic Neoparamoeba perurans is the most relevant species, being a real pathogenic parasite in farmed salmon. It causes amoebic gill disease, which causes a progressive hyperplasia of epithelial cells in the gill filaments and lamellae. Nodular gill disease is its analogue in freshwater fish but the causative agent is still not clear, although several amoebae have been identified associated to the lesions. Other species have been described in different fish species, affecting not only gills but also other organs, even internal ones. In some cases, species of the genera Naegleria or Acanthamoeba, which also contain pathogenic species affecting humans, are usually described affecting freshwater fish species. As endocommensals, Entamoebae species have been described in the digestive tract of freshwater and marine fish species, but Endolimax nana can reach other organs and cause systemic infections in farmed Solea senegalensis. Other systemic infections caused by amoebae are usually described in wild fish, although in most cases these are isolated cases without clinical signs or significance.
1. Introduction
Generally speaking, the term amoeba describes a particular type of unicellular organism characterised by the ability to modify its shape mainly by the development of pseudopods. In the past, most amoebae were placed in the single Rhizopoda clade of the Sarcodina supergroup. However, recent studies based on molecular biology clearly indicate that amoebae are no longer a single taxonomic group but represent a clear example of a polyphyletic group, including members of different Supergroups, including Amoebozoa, Rhizaria, Excavata, Heterokonta, Alveolata and Opistokonta.
Amoebae, due to their characteristics, live in aquatic environments or can be present in aqueous fluids and humid environments and they can be also associated with external surfaces and the internal environments of animals. For this reason, most of the amoebae can be found mainly as free-living organisms (FLA) in aquatic environments [1], including natural environments but also man-made water storage and networks, such as drinking water or industrial cooling towers, and are also in soils. In addition, some species can also live in association with other organisms as ecto and endocommensals and also as ecto and endoparasites, acting in some of these cases as parasites and in many cases can alternate FLA phases with phases with a different level of association with organisms.
2. Diseases Caused by Amoebae in Human and Veterinary Medicine
As pathogens, amoebae are well-known organisms in human and veterinary medicine. In human medicine, several species have been described as pathogens or opportunistic pathogens. Amongst these species, probably the most well-known species and frequently reported in clinical cases in humans are the free-living amphyzoic amoeba Acanthamoeba spp., Naegleria fowleri, Balamuthia mandrillaris and Sappinia sp. [2]; and the Entamoeba complex [3,4] (particularly E. histolytica but also other opportunistic species such as E. coli), but also Endolimax nana or Iodamoeba buetschlii as the most pure form of endocommensals.
Amphyzoic amoebae, unlike true parasites, are not well adapted to parasitism, so they tend to be very aggressive within the host, causing their death in most cases. For instance, Acanthamoeba spp. is one of the most commonly isolated amoebae in environmental samples and they can cause granulomatous amoebic encephalitis (GAE), cutaneous acanthamoebiasis and amoebic keratitis (AK) in humans; or the percolozoan Naegleria fowleri causes primary amoebic meningoencephalitis (PAM) in humans, which is an acute, fulminant and fatal disease [5]. Water supplies, swimming pools, freshwater ponds, lakes and thermally polluted waters have been recognised as sources of human infections.
As endocommensal or endoparasite species of the human intestine Entamoeba histolytica is the only species that is considered pathogenic. However, Entamoeba dispar, E. moshkovskii, E. hartmanni, E. coli, E. polecki, Endolimax nana or Iodamoeba buetschlii are also very frequent and they are usually considered non-pathogenic. Infections by Entamoeba histolytica usually include an intestinal phase that ranges from asymptomatic colonisation to severe invasive infections (dysentery, colitis); and an extraintestinal phase, generally affecting the liver (amoebic liver abscess), with eventual progression to other organs (lung, brain, heart) through blood dissemination [6]. Apart from E. histolytica, other archamoebae species are able to cause diseases in their hosts. For example, gastrointestinal disorders have been reported associated with diverse species of Entamoeba [7,8], but also by Endolimax nana [9]. Furthermore, E. dispar has also been reported causing hepatic lesions [10] and E. nana has been described to be involved in skin processes and rheumatoid arthritis (see [11] and references herein). Iodamoeba buetschlii has also been reported to be able to cause brain granuloma [12].
In veterinary medicine, amphyzoic amoebae have also been reported in different vertebrates, such as non-human primates, dogs, bulls, horses, sheep, kangaroos… [13] and their pathogenesis varies depending on the species involved. They generally cause similar lesions to those in humans; granulomatous inflammatory lesions in nervous tissue, but also lesions in other organs or even systemic infections. Similarly, endoparasitic amoeba are also described in other vertebrates with different degrees of pathogenicity. Entamoeba nuttalli or E. invadens are highly related species to E. histolytica with similar pathogenicity, that can affect non-human primates and reptilians (see [14] and reference herein), or E. bovis and E. ovis infecting ruminants [15,16]. Iodamoeba and Endolimax are also endocommensal or facultative parasitic organisms in the intestinal tract of other vertebrates. Species of Endolimax have been reported for a variety of vertebrate hosts [17,18,19,20,21,22,23]. Although most of them are considered endocommensals, some of them are pathogenic.
In addition to the effect of primary pathogens in human and animal diseases, it is also very important to remark the recently described role of amoeba as reservoirs for human and animal pathogenic bacteria [24] and giant viruses [25]. Concerning these peculiar association of amoebae with internalised bacteria, it is particularly interesting to compare this relationship with the association of macrophagic cells, a cell type also characterised by the ability to develop pseudopods, with several intracellular pathogenic bacteria such as Chlamydia and Rickettsia and pathogenic bacteria that develop autophagy mechanisms [26].
3. Amoebae and Fish Diseases
As fish are aquatic organisms, it is understandable that fish and amoebae can interact in different ways. The presence of amoeba or amoeboid organisms in association with fish has been described since the beginning of the 20th century. In some cases, these descriptions were limited to the observation or isolation of amoebae from fish, without a clear description of the specific relationship with the fish or even the specific fish tissue. As FLA are ubiquitous organisms in aquatic environments, it is not unlikely that the amoebae ‘isolated’ from fish simply correspond to sampling contamination during the capture or manipulation of the fish. This situation is particularly probable for the identification of amoebae isolated from the fish skin or gills. However, in many descriptions, the relationships between amoeba and the fish are more clear and solid. In this scenario, most of these relationships are based on four different types: ectocommensals, ectoparasites, endocommensals and endoparasites.
As ectocommensals, it is important to stress that particularly gills and also skin constitute a favourable ground for the development of amoebae. Gill and skin mucous can represent a source of nutrients for the amoebae and moreover, the gill and skin microbiome can also be a valuable source of bacterial cells for the feeding activities of the amoebae, in the same way as bacterial biofilms in the environment [27] and even other water storage places [28,29]. All these environments represent a relevant source of nutrients for grazing amoebae. In recent years, the fish intestinal, gill and skin microbiomes are attracting increasing attention in its role in the different epithelial mucosa as the first defence barrier against pathogens. It seems that the mucous surface acts like some kind of ‘no man’s land’ with a delicate and very complex balance between the potentially enhancing substances (mucins, glucose, other carbohydrates, nucleosides, free aminoacids, small peptides) and limiting factors (mostly enzymes such as lysozyme, antibodies, complement factors, antimicrobial peptides (AMPs) such as piscidin, lectins and IFN) present in the mucus composition [30,31]. A similar explanation can be given for endocommensals, as most of the endocommensal amoebae described in fish are found in the digestive tract and only in a few cases are associated with the disease [32]. The mutualistic relationship and relevance of the intestinal microbiota for the digestion and other physiological functions in animals is widely known, so the intestinal tract is also a suitable aqueous microenvironment for the development of endocommensal amoebae.
Amphyzoic species have demonstrated their ability for an endozoic way of life also in aquatic environments. Dyková et al. [33] underlined the pathogenic potential of Naegleria sp. for fish since, although most of the Naegleria isolates in different freshwater fish species came from clinically healthy fish (mainly from gills), in some cases they were present in internal organs and even in systemic infections [33,34]. Acanthamoeba spp. are also able to colonise the organs of freshwater fishes [35,36], and the strains isolated in some cases were closely related to those commonly isolated from cases of human infections, especially Acanthamoeba keratitis [36]. Amphyzoic characteristics increase the capacity to survive and increases the complexity of their control, as reinfections are much more likely due to their resilience in the environment and therefore, with limited possibilities for the control. Although these amphyzoic amoebae can cause granulomas in different organs of the host, they are typically isolated from normal organs, like other amoebae genera such as Protacanthamoeba, Vexillifera or Vannella [37,38,39]. Protacanthamoeba bohemica, a closely related species of the genera Acanthamoeba and Balamuthia, was isolated from the liver of tench (Tinca tinca) [38]. Vannella spp. are very common in isolates of various organs from freshwater and marine fish, particularly from the gills [39,40,41,42]. Although in some studies, species of Vannella were isolated from Atlantic salmon and rainbow trout with lesions in gills [43,44,45], their role in these diseases is unknown and, in general, they are considered non-pathogenic species. Vexillifera expectata was isolated from the liver of perch (Perca fluviatilis) [37] and more recently three new Vexillifera species (V. bacillipedes, V. multispinosa, V. fluvialis, V. tasmaniana) have been isolated from the gills and internal organs of freshwater fish [46], without causing apparent lesions. Thecamoeba has been isolated from the healthy gills of turbot (Scophthalmus maximus) [40] and years ago Sawyer et al. [47] pointed to T. hoffmani as the causative agent of gill disease and mortalities in freshwater salmonid fingerlings. Cochliopodium spp. are a scale-bearing amoeba that inhabit both marine and freshwater environments [48,49]. C. minus was also described from different internal organs and the gills of asymptomatic perch (Perca fluviatilis) [50] and the catfish (Heteropneustes fossilis), the latter after experimental infection. A similar species to C. minus was also isolated from the spleen of gudgeon Gobio gobio and the gills of wels catfish (Silurus glanis) [40]. Saccamoeba sp. was isolated from dead aquarium fish [51] and amoeba isolated from the gills of Atlantic salmon Salmo salar was phylogenetically related to S. limax, although the ultrastructure did not allow its identity confirmation [52]. Nolandella has been isolated from the healthy gills of Atlantic salmon (Salmo salar) [43], turbot (Scophthalmus maximus) and rainbow trout (Oncorhynchus mykiss) [53,54]. Therefore, it does not seem to be a pathogenic genus, however English et al. [41] have also isolated it from symptomatic gills, along with other amoeba species. Nuclearia pattersoni was described from the gills of roach (Rutilus rutilus) [55].
Among all these different genera of amoeba, probably the most studied species are those that have been isolated or detected on the gills of Atlantic salmon (Salmo salar), and that may accompany some type of gill lesions, such as Acanthamoeba, Flabellula, Heteroamoeba, Vannella, Vexillifera, Mayorella or Nolandella [41,43,56], but their pathogenic role for the moment remains unknown.
Concerning the more specific relationships of pathogenic amoebae, some species have been described as truly pathogenic for fish in the same way as in other vertebrates. However, due to the special characteristics of the fish as 100% aquatic vertebrates, diseases and pathology associated with amoeba can be clearly differentiated in amoebic gill diseases and internal/systemic amoebic diseases. Both present remarkable peculiarities and differences and for this reason are presented in two different sections.
3.1. Gill Diseases Caused by Amoebae
Gill diseases associated with amoebae are mainly produced by Amoebozoa, most of them lobosan amoebae, in other words, amoeba with lobosan-shaped (broad and round) pseudopodia. Amoebic gill diseases can be present in marine fish and freshwater fish and although the causative agents can be detected in fish in the wild, disease is only described in fish under aquaculture or captivity conditions. In marine fish, the most widely known disease associated with amoebic organisms is AGD (amoebic gill disease) associated with Neoparamoeba perurans, and in freshwater the disease is usually described as NGD (nodular gill disease) with different species of amoebae involved in this complex.
3.1.1. Amoebic Gill Disease (AGD)
AGD is nowadays one of the most relevant diseases in Atlantic salmon (Salmo salar) farming in different geographical locations (Norway, Scotland, Ireland, Australia, Chile), with a substantial disease burden including high mortality (sometimes up to 85% in a total production cycle) and a substantial reduction in growth. It is also one of the most relevant gill diseases in this species [57]. The disease and particularly the fish response, as it plays a major role in the pathogenesis and effects of the disease, has recently been extensively reviewed [58]. Although the main impact of the disease is in Atlantic salmon, other salmonids such as coho salmon (Oncorhynchus kisutch), chinook salmon (Oncorhynchus tshawytscha), rainbow trout (Oncorhynchus mykiss) and brown trout (Salmo trutta) can be affected. Particularly, rainbow trout can also be affected in freshwater by other amoebic species, but this is usually referred to as nodular gill disease and is presented in the following section. Flatfish species can also be affected and farmed turbot (Scophthalmus maximus) is the species where AGD has had a more relevant impact, but the disease and/or the pathogen have been described in other species such as halibut (Hippoglossus hippoglossus) [59] and Senegalese sole (Solea senegalensis) [60]. Other species where AGD is described are sparidae species [54,61,62], such as sea bream (Sparus aurata), sharpsnout seabream (Diplodus puntazzo) and black seabream (Acanthopagrus schlegelii) and also species such as sea bass (Dicentrarchus labrax) [63], ayu (Plecoglossus altivelis) [64], rock bream (Oplegnathus fasciatus) and grey mullet (Mugil cephalus) [62]. However, in these species, the available information about the disease is much more limited than in salmon. A particular case is the infection described in species such as ballan wrasse (Labrus bergylta) [65], corkwing wrasse (Symphodus melops) and lumpfish (Cyclopterus lumpus) [66]. In these species, the development of AGD is probably due to their close contact with potentially affected Atlantic salmon in cages, due to the frequent use of these species as cleaner fish as a system for the biological control of sealice (Lepeophtheirus salmonis) in Atlantic salmon farms. All these affected species are also widely reviewed in Oldham et al. [67] and Marcos and Rodgers [58].
The causative agent of AGD is the amphyzoic amoebae Neoparamoeba perurans (formerly known as Paramoeba perurans). Some years ago, when the disease was first described and studied in the USA and Tasmania in different salmonids reared in marine water, the disease was initially associated with the species Paramoeba pemaquidensis [68,69]. In the following years, more detailed studies based on morphological and molecular characterisation of Neoparamoeba strains isolated from the gills of sea-caged Atlantic salmon but also from marine sediments and the net material of sea-cages were developed [70], stressing the differences between the fish-pathogenic Paramoeba species and other non-pathogenic Paramoeba free-living species isolated in the gills of affected fish such as Paramoeba branchiphila and P. aestuarina. A few years later, further studies using molecular biology including culture-isolated P. pemaquidensis and P. branchiphila and also non-cultured, gill-derived (NCGD) amoebae from AGD-affected Atlantic salmon [71] allowed the identification of these NCGD amoebae as the organism responsible for AGD, and the naming of this species as Neoparamoeba perurans n. sp. The species name was selected after the Latin verb ‘peruro’ that means ‘burn up, consume, or inflaming’, referring the reaction displayed in AGD, although inflammation is not the main pathological component observed in AGD. The new molecular tools developed from this study were applied in archived and new samples in order to clarify the implication of this new species in the development of AGD lesions [71] and reinforcing its role as the aetiological agent for AGD. Some years later, the genus Neoparamoeba was reviewed as a consequence of a study on mortalities on sea urchins associated with amoebae [72] and the name of the genus was moved to Paramoeba. However, in 2019 the name of the genus was reverted again to Neoparamoeba, and the former name is considered as a synonym.
The disease. AGD is typically a chronic disease, but with a relatively fast development [72], affecting not only gills but also causing a progressive systemic involvement of other organs due to their effects on respiratory, osmoregulatory and circulatory functions. Affected fish become lethargic, with reduced swimming speed, feeding rates and growth and display signs of respiratory distress (increased rate of ventilation).
It is characterised by the development of a progressive hyperplasia of epithelial gill cells in the primary and secondary (lamellae) filaments that leads to an increase of the thickness of the gill epithelia, progressive formation of synechia and lacunae between filaments and finally total obliteration of the interlamellar space [73]. These alterations in gill structure lead to an impairment in gas exchange through the gills and acid-base regulation and consequently in the respiratory function and general metabolism. These changes in the respiratory capacity and in the acid-base homeostasis in the affected fish can also affect the metabolism of other organs such as liver or heart, although there are still discrepancies about the real effects in the gas exchange, oxygen transport and aerobic metabolism in the fish [72].
As AGD also affects the whole structure of the gill epithelium including chloride cells [74], physiological disturbances in the osmoregulation are also involved in the physiopathogenesis of the disease. The gill mucus is also affected and in addition to the increase in mucus production, relevant changes have been detected in the gill mucus in AGD affected fish [75].
Factors. High water temperatures (the optimal range for AGD development is 10–18 °C) and also high salinity are considered as two of the main risk factors for the development of the disease and in fact, in some places the outbreaks display a substantial connection with these factors. When seawater salinities drop under 28 per thousand the disease usually diminishes in its intensity. After a first infection, salmon can develop a certain resistance against the disease [76]
Since N. perurans is considered an amphyzoic free-living amoeba, the role of the gill microbiota in the ecology of amoebae in this specific microenvironment has recently received attention [77].
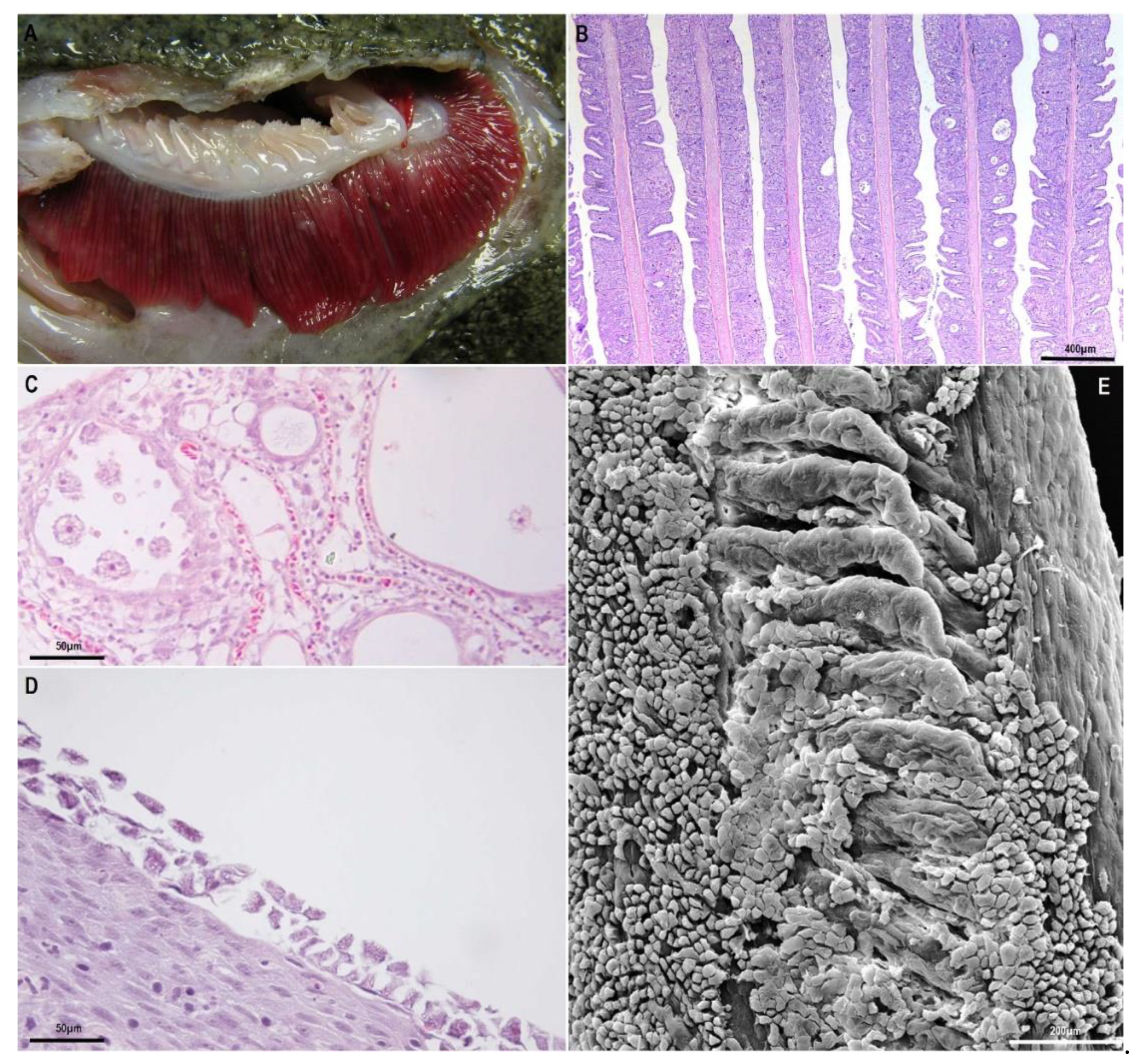
Diagnosis. AGD diagnosis is normally done through the identification of macroscopic and microscopic lesions in the gills. Macroscopically, AGD is usually characterised by the development of white patches that can develop in the respiratory surface of all four gill arches (Figure 1A). A scoring system based on macroscopic lesions [78] has been recently developed to assess the evolution of the disease. Amoebae can also be seen in gill wet mounts, although sometimes cannot be easily seen as they can remain attached to the gill epithelium or entangled in the proliferative epithelial tissue and lacunae formed in the gills.
Histopathology is usually the most frequently used diagnostic method to complement macroscopic gill assessment, as AGD lesions are quite characteristic (Figure 1B), and allows a more accurate assessment of the gill condition and stage of the disease. The lesions are those previously indicated in the description of the disease, and amoebae can be more easily recognised inside the lacunae (Figure 1C). However, AGD-related lesions are not pathognomonic, as some other chronic and proliferative gill diseases may display a similar histopathologic pattern. Particularly CGD (complex gill disease) is the current term used to describe chronic proliferative and inflammatory problems in the gills of Atlantic salmon [79] and although in many cases AGD and Paramoeba perurans can be diagnosed as a single and specific pathological entity, many times AGD can be involved in multifactorial cases of CGD.
As many other non-pathogenic amphyzoic amoebae can be found in gills with lesions, a complementary identification of the presence of N. perurans in the gills is recommended to have a robust diagnostic of AGD. This can be done by routinely sampling the gills using cotton swabs and PCR analysis for the detection of N. perurans [80].
Disease management. AGD management and treatment is usually difficult and complex. Although some oral treatments such as bithionol were tested in the past [81], freshwater and hydrogen peroxide baths are currently the two most frequently used treatments and recently, peracetic acid has also been included in the potential weaponry against this disease [82]. Freshwater baths have been the traditional control system used in Tasmania, but this treatment is highly dependent on the availability of large volumes of freshwater. As good quality freshwater from drinking water networks, rivers, lakes or reservoirs is not always available everywhere, hydrogen peroxide has been considered an acceptable option. Baths are usually performed with the use of tarpaulins in the cages, specific treatment cages already provided with tarpaulins, barges or more recently, well boats, some of them equipped with reverse osmosis systems to supply the ship with freshwater. However, the efficacy of the treatments and the safety of the treated fish are also highly dependent on the environmental conditions, the water quality used in these treatments [83] and also on the fish health condition prior to treatment.
For example, hydrogen peroxide treatment is most efficient at low temperatures [84] and also toxicity increases at high temperatures. This is the reason why AGD treatments should be done as preventive baths during cold seasons and before the onset of the clinical problems at warmer temperatures. This is the most efficient way to keep AGD at bay as it is really difficult to treat and manage once the process is triggered. Regular macroscopic assessments of the aspect of the gills in the different cages or batches using a scoring system is highly recommended and a normal practice in the farms under AGD risk.
Other AGD management strategies include functional feeds, genetic selection and vaccination.
Commercial functional feeds are used to control and reduce the impact of the disease. Most of these feeds are physiology and immunosupportive-based diets formulated to reduce the impact of the disease by increasing immune protection and enhancing the fish recovery.
After some years of research in this field, roe and fry from specific breeds with increased resistance to AGD are nowadays commercially available.
There is a relevant interest in the study of the development of the immune response in AGD mainly to find an efficient protective vaccine, but also to understand the full host response of the salmon immune system against the parasite [58], as the proliferative response triggered by the parasite is itself, one of the problems associated with the disease. Some experimental vaccines were tested in the past and nowadays there are several projects with different vaccine development strategies. However, at this moment there are no available commercial vaccines against AGD.
3.1.2. Nodular Gill Disease (NGD)
A similar pathological condition related to amoebic infection in the gills has also been described in freshwater fish species. This disease is frequently described as nodular gill disease (NGD) but occasionally it has been referred to as proliferative gill disease (PGD) [85]. The use of the name PGD to describe a disease is not recommended as PGD refers to a general epithelial proliferative condition described in a wide number of gill diseases in several fish species associated with different pathogens (e.g., bacterial gill disease (BGD) in salmonids associated with Flavobacterium, hamburger gill disease or PGD from Henneguya in catfish [86] or complex gill disease (CGD) as a multifactorial syndrome in salmon [79]).
In some ways, NGD can be described as the counterpart of AGD in freshwater, although there are some substantial differences between AGD and NGD that need to be highlighted.
The causative agent(s). NGD was described in North America as a specific gill disease affecting rainbow trout [87] usually associated with bacterial gill diseases and in this paper a large number of protozoa were described associated with these lesions. Sometime later, the disease was associated with the presence of so-called ‘A cells’ suggesting similarity of these cells to amoebae [88]. These ‘A-cells’ were lately recognised as amoebae causing similar lesions in rainbow trout and also in arctic charr (Salvelinus alpinus) and tentatively identified as Cochliopodium sp. [89]. Since then, several reports of NGD mainly in farmed rainbow trout and also brown trout (Salmo trutta) have been described in North America and Europe [90] and also in farmed chinook salmon (Oncorhynchus tshawytscha) [91].
The first description of amoebae in farmed salmonid gills was from Sawyer et al. [47]. However, there are some other previous descriptions of amoebic organisms in salmonid fingerling gills, but not associated to NGD or disease. These amoeboid organisms were initially characterised as Thecamoeba hoffmani and Schyzamoeba salmonis. However, the accurate identification of the species was based on general microscopic features and not on more suitable methodologies. Some years later, Dyková et al. [44] published a study based on amoebae isolated in rainbow trout (Oncorhynchus mykiss) farms in Germany, identifying different strains of Acanthamoeba, Hartmannella, Naegleria, Protacanthamoeba and Vannella from the gills with clear NGD lesions. However, in this study it was not possible to identify which of them were the main organisms responsible for the lesions observed. The authors suggested Acanthamoeba, Hartmannella and Protacanthamoeba as the species with highest pathogenic potential. Again, as in marine amoebae, the main challenge was the identification of the potentially pathogenic amoebae from the relatively high diversity of free-living amoebae that can be present in the gills. Later studies [45] also identified another species, Roghostoma minus highly related with NGD infections, highlighting the heterogeneous diversity of amoebic species potentially involved in the NGD development in freshwater, in contrast to the scenario in seawater, where AGD is mainly related with one species.
NGD was and still is a very common disease in rainbow trout farms in Europe [85], although its relevance and burden has not been properly addressed until recently. Recently, some studies on NGD outbreaks in Italy [47,90,91,92] highlighted the re-emergence of NGD as a relevant disease in trout farming.
The disease. NGD symptomatology in rainbow and brown trout is similar to the symptomatology in AGD-affected salmons: fish are less active, display respiratory distress, changes in swimming behaviour and reduced growth. Mortality is usually noticed some weeks after the first symptoms appear. The external aspect of the lesions in the gills is also characterised by the presence of whitish areas in the gill filaments and increased presence of mucus. Microscopic histopathological lesions are characterised by a particularly intense epithelial hyperplasia leading to partial or complete lamellar fusion and even, fusion between adjacent filaments. As a point of difference with AGD, it is important to highlight that the presence of inflammatory infiltrate has been described associated with the lesions [47,90,91,92] although in some cases, very weak or no inflammation is also seen (personal observations). In NGD, the distal region of the filaments is usually the most affected part of the gills (in contrast to AGD) and sometimes filaments display a club or maze aspect. Here, the epithelial hyperplasia is particularly intense and sometimes displays spongiosis. In the most advanced cases amoebae can be seen as flattened organisms attached to the surface of these proliferative areas or in the middle of the lacunae formed between the fused lamellae (Figure 1D,E).
Factors. The epidemiology and factors affecting NGD development in freshwater salmonids is much less understood than in seawater AGD so the role of water characteristics is not known. NGD outbreaks are usually associated with other mixed pathological conditions and infections (other protozoan parasites, monogeneans, BGD by Flavobacterium, Saprolegnia) that makes the diagnosis and the prognosis more complex and difficult.
Diagnosis. The diagnosis is usually done by combined macroscopic and histopathological lesions. The identification of amoebae present in this case has a scarce clinical relevance as many different species of amoeba can be found [44,45]. However, from the scientific point of view it is necessary to have a major insight in the amoebae involved in the NGD cases and particularly in the specific role of the species in the pathogenesis of the disease.
Disease management. Very little information is available about treatments for NGD. General treatments for ectoparasites (formalin) can be effective but no specific data on that is available. No effect was found in 10 ppm chloramine T baths (1 h), although this treatment was mainly directed to treat a BGD co-infection [91]. Surprisingly, no data on the use of salt baths are described in the literature. As freshwater is one of the most efficient treatments to control marine amoebae in AGD, the potential effect of salt baths cannot be ruled out simply using an osmotic shock to kill and remove the amoeba. In fact, salt (sodium chloride) is one of the best, easiest, cheapest and sometimes forgotten options used to treat certain ectoparasites and reduce osmotic stress in freshwater fish. Repeated short treatments (10–20 min) with 10–30 g/L salt are usually recommended to control ectoparasites (ciliates, monogeneans) in aquaculture, so its use to control freshwater NGD is highly promising.
3.2. Systemic Diseases Caused by Amoebae
Systemic amoebic infections in fish have been known for a long time, but they have been reported causing significant lesions in internal organs only occasionally. Systemic granulomatous infection due to an amoebae-like organism was described 40 years ago in goldfish (Carassius auratus) [93,94,95,96]. Systemic infection by amoebae-like organisms were also occasionally described in other fish species such as dwarf (Colisa lalia) [97], European wels catfish (Silurus glanis) [98] or pompano (Trachinotus falcatus) [61]. More recently, a similar pathological condition was described in Senegalese sole (Solea senegalensis) [99] and tench (Tinca tinca) [100]. In both studies, due to the absence of mitochondria in the amoebae-like organisms detected, Archamoebae were suggested as the aetiological agent of the infection. Amoebae belonging to the class Archamoebae, contrary to free-living amphizoic amoebae, are characterised by the lack of mitochondria [15,101,102,103], probably related to their adaptation to parasitism. Among these parasitic amoebae species, the genus Entamoeba, as stressed before, is the most important one because it parasitises several vertebrate species (e.g., [15,100,101,102]), including fish [94]. Entamoeba salpae [104], E. gadi [105], and E. molae [106] were described from marine fish species, whereas E. pimelodi [107], E. ctenopharyngodoni [108] and E. chiangraiensis [109] have been described from freshwater fish. Excluding Entamoeba, most members of Archamoeba are not well studied, probably due to the relatively minor clinical importance and the difficulties associated with their laboratory cultivation [110]. Fish are not an exception, and the majority of mentions of these amoebae refer to specific findings or isolation in a specific fish species. However, the systemic granulomatosis in Senegalese sole (Solea senegalensis), has sparked considerable interest due to the economic losses it causes in the fish farming industry and has been investigated in more detail.
Amoebic Granulomatous Disease in Cultured Senegalese Sole
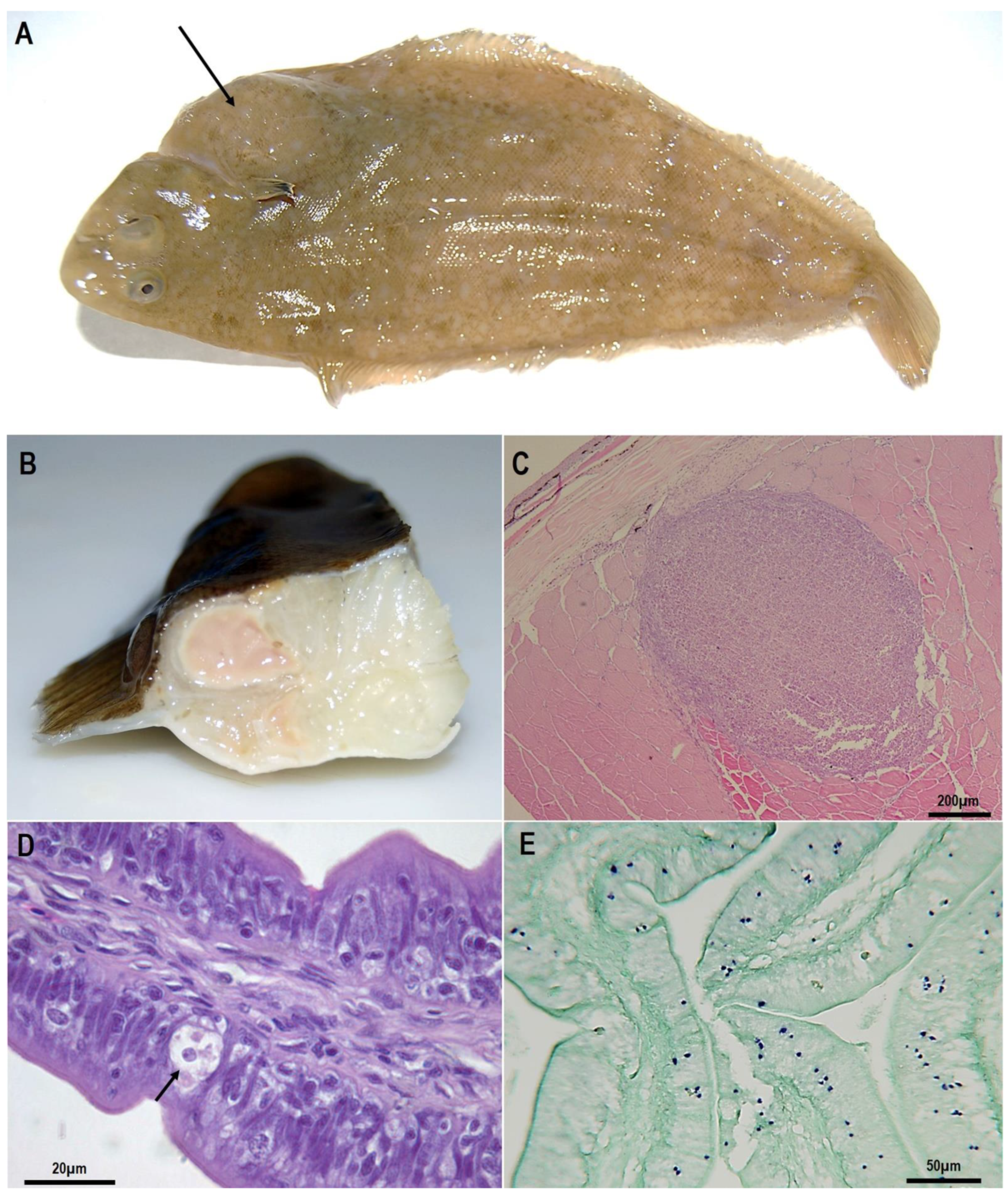
The amoebic granulomatous disease of farmed Senegalese sole (Solea senegalensis) in the Spanish Atlantic coast was first described in 2010 [99]. Affected fish presented protuberances or lumps in the muscle, often noticeable at the skin surface (Figure 2A).
The disease does not cause a high mortality but the prevalence in the farms can be very high [111], causing severe economic problems as the affected fish are unmarketable due to their aspect.
Since then, this pathological condition has discontinuously affected different farms on the European Atlantic coast.
Unicellular organisms were initially found in histopathological studies closely associated with the granulomatous lesions, strongly suggesting a parasitic origin of the disease. The differential diagnosis from other parasitic problems included different groups of protozoans such as flagellates, apicomplexa, microspora, mesomycetozoa or amoebae [99]. Finally, ultrastructural features of the organisms clearly pointed to amoeba and molecular studies allowed the identification of Endolimax piscium as the causative agent [112].
The causative agent. The archamoebic species Endolimax piscium is a species phylogenetically related to the entamoebids E. nana and Iodamoeba spp. [112], which are enteric commensals and parasitic species in humans and other animals. Entamoebids usually have two stages in their life cycle [3,110]: a trophozoite stage (vegetative and infectious form, which is the responsible for the disease) and a cyst (a form of resistance, which is the responsible for transmission). Up to now, the only stage characterised at the moment for E. piscium is the trophozoite. The trophozoite of E. piscium is 2 to 5 μm in diameter and contains one vesicular nucleus with a large and central round nucleolus. Endoplasmatic reticulum, dictiosomes and a variety of structures such as some single-membrane bound vesicles, putative digestive vacuoles containing particulate material, myelinic figures or products of lysosomal action are also described in the cytoplasm [99,112]. Like other members of Archamoebae, they are amitochondriates, but they contain double-membrane-bounded, electron-dense organelles with no apparent cristae resembling mitosomes usually being observed.
The disease. Macroscopic lesions in symptomatic fish consist of nodules with an abscess-like aspect located in different tissues, especially in muscular tissue but also in the liver, digestive tract, gonads, heart and kidney. These nodules are clearly distinguishable from the rest of the tissues and usually present a soft and liquefied consistency (Figure 2B).
Histologically, the nodules correspond to chronic granulomatous inflammatory lesions with a large core of homogeneous necrotic tissue surrounded by fibroblasts and macrophages (Figure 2C). Amoebae appear as ring-like layers surrounding the periphery of the lesions, between the external inflammatory reaction and the necrotic tissue, free or within the cytoplasm of macrophages [99]. In some cases, extensive necrosis and diffuse inflammatory areas are also found, especially in muscle and the liver.
In addition to the lesions, E. piscium is also frequent within the intestinal epithelium of fish without apparent lesions (Figure 2D). One of the most important issues of this disease is that E. piscium can be present in the intestine of both symptomatic (i.e., displaying granulomatous lesions) and asymptomatic fish [99,113,114], which points to an endocommensal behaviour of this species and it is considered the initial stage in the development of the disease [113]. In moderate to high parasite intensity, amoebae can reach submucosa [113], which indicates a proliferation of the parasite and the progression to other organs, with a similar behaviour to E. histolytica. An haematogenous dissemination is the most accepted route of liver invasion and other organs in the case of E. histolytica trophozoites [115,116]. Although in the case of E. piscium this mechanism is still unknown, the connective tissue (probably facilitated by host phagocytic cells) may play a significant role and an haematic route should not be disregarded, especially in advanced infections with high levels of parasitemia [113].
The amoebic granulomatous disease does not cause important mortalities in Senegalese sole (Solea senegalensis); actually it is a long-term chronic disease. The affected fish can survive for a long time with lesions. The detrimental effects in fish, such as reduced growth performance and higher susceptibility to other diseases, are evident, and the extensive lesions make the fish unmarketable after harvesting.
Factors. The triggering factors or specific conditions for the development of overt clinical infections, that facilitate the transition from an endocommensal to an aggressive and systemic parasite, are still unknown.
Temperature has been suggested as an important factor [99,114], but the appearance of symptomatic fish at any time of the year, even in the cold months of January and February, seems to rule it out.
Many amoeba outbreaks can be favoured by immunodepression or suboptimal environmental conditions [98,117]. Brain lesions in humans caused by Acanthamoeba species, Balamuthia or even Entamoeba, among others, are usually seen in individuals suffering from other chronic diseases, having compromised immune systems, or during outbreaks of bacterial infections [118]. In this sense, Constenla et al. [114] pointed to Tenacibaculum infection as one of the important factors that led to positive cases of E. piscium in fish muscle during an experimental infection.
Water recirculation conditions, as in the case of the pre-fattening and fattening systems of sole, could be favouring the survival and permanence of these stages in the rearing system, external to the host, facilitating a proliferation of the parasite among the fish particularly in intensive farming condition facilities.
Diagnosis. Histopathological observation combined with molecular methods (ISH, PCR and qPCR) are the diagnostic methods currently available for the diagnosis of this disease. The histopathological observation is limited by the minute size and cryptic nature of E. piscium, but useful if parasites are associated with the typical granulomatous responses [113]. Diagnosis of subclinical infections by histopathological observation usually is not enough for the confirmation of the diagnostic and only achievable by an in-depth analysis by an experienced pathologist. However, confirmation of the diagnosis can be obtained with high predictive values and high accuracy (>99%) if combined with molecular hybridisation techniques [113]. The ISH efficiently detects E. piscium trophozoites present in different tissues, allowing the identification of the parasite even when found in small numbers or non-associated to lesions (Figure 1E) and therefore, is the reference confirmatory method, particularly in asymptomatic fish.
Probes of PCR and qPCR are also available and offer a rapid diagnostic for a high number of samples, but the sampling for these methods is very important so as not to reduce their sensitivity. For instance, the sensitivity of PCR methods decreases in intestine samples with low parasite intensities, probably due to the patchy distribution of E. piscium within this tissue, but this sensitivity clearly increases if homogenates of all or a large part of the intestine are used instead of small pieces [113].
Disease management. To date, no treatments have been described to control the disease, and therefore prophylaxis appears as the only option for its management. In this context, it is essential to detect not only the parasite in carrier fish [113], but also stages outside the host that may be retained at the bottom of the tanks particularly in recirculation systems. It is important to keep in mind that Endolimax nana and entamoebid species commonly include parasites with two stages in their life cycle: a trophozoite and a cyst [110]. The latter is the resistant stage, which allows the parasite to remain in the environment after leaving the host and until reaching new hosts to continue their life cycle. Although no cyst stages have ever been identified in E. piscium, positive results by qPCR were obtained from water samples after filtration during an experimental infection [114]. The potential encysting capacity of this amoeba is an important handicap for fish farm prophylaxis because amoeba cysts can be very resistant to temperature, desiccation and disinfection treatments [119]. The same experimental infection demonstrated that E. piscium can be horizontally transmitted to healthy fish by cohabitation with no direct contact between fish, only contaminated water [114]. In addition, in intensive culture conditions, where bottom-dwelling flatfish are reared at high density, the faecal−oral contagion route is also likely to occur readily in intestinal parasites.
Early detection of the parasite in the farm should be considered a priority for the management of this disease, as the muscular lesions developed can prevent the commercialisation of the fish after an important investment in their feeding and maintenance.
In most cases, only after a complete and thorough sanitary break and a careful assessment of the absence of the parasite in the new fry or juvenile batches was it possible to eliminate the disease from the facilities. The use of flow-through systems and the reinforcement of hygienic and disinfection technology and husbandry procedures within the recirculation system such as implementation of powerful UV systems, post-filtration disinfection with ozone or similar systems and periodical cleaning of tank bottoms and surfaces can also reduce the risk of the disease in the systems.
Author Contributions
Both authors contributed equally to this work. Both authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We are grateful to Carlos Zarza for his comments and suggestions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Samba-Louaka, A.; Delafont, V.; Rodier, M.H.; Cateau, E.; Héchard, Y. Free-living amoebae and squatters in the wild: Ecological and molecular features. FEMS Microbiol. Rev. 2019, 43, 415–434. [Google Scholar] [CrossRef] [PubMed]
- Król-Turmińska, K.; Olender, A. Human infections caused by free-living amoebae. Ann. Agric. Environ. Med. 2017, 24, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Gomila Sard, B.; Toledo Navarro, R.; Esteban Sanchis, G. Amebas intestinales no patógenas: Una visión clinicoanalítica. Enferm. Infecc. Microbiol. Clin. 2011, 29, 20–28. [Google Scholar] [CrossRef]
- Shirley, D.A.T.; Farr, L.; Watanabe, K.; Moonah, S. A review of the global burden, new diagnostics, and current Therapeutics for amebiasis. Open Forum Infect. Dis. 2018, 5, ofy161. [Google Scholar] [CrossRef] [Green Version]
- da Rocha-Azevedo, B.; Tanowitz, H.B.; Marciano-Cabral, F. Diagnosis of Infections Caused by Pathogenic Free-Living Amoebae. Interdiscip. Perspect. Infect. Dis. 2009, 2009, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Espinosa-Cantellano, M.; Martínez-Palomo, A. Pathogenesis of intestinal amebiasis: From molecules to disease. Clin. Microbiol. Rev. 2000, 13, 318–331. [Google Scholar] [CrossRef]
- Jetter, A.; Walderich, B.; Britten, D.; Mete, O.; Göral, V.; Burchard, G.; Ackers, J. An epidemiological study of Entamoeba histolytica and E. dispar infection in eastern Turkey using a colorimetric polymerase chain reaction. Arch. Med. Res. 1997, 28, 319–321. [Google Scholar]
- Heredia, R.D.; Fonseca, J.A.; López, M.C. Entamoeba moshkovskii perspectives of a new agent to be considered in the diagnosis of amebiasis. Acta Trop. 2012, 123, 139–145. [Google Scholar] [CrossRef]
- Stauffer, J.Q.; Levine, W.L. Chronic diarrhea related to Endolimax nana. Am. J. Dig. Dis. 1974, 19, 59–63. [Google Scholar] [CrossRef]
- Costa, C.A.X.; Brito, K.N.O.; Gomes, M.A.; Caliari, M.V. Histopathological and immunohistochemical study of the hepatic lesions experimentally induced by Entamoeba dispar. Eur. J. Histochem. 2010, 54, 170–174. [Google Scholar] [CrossRef] [Green Version]
- Poulsen, C.S.; Stensvold, C.R. Systematic review on Endolimax nana: A less well studied intestinal ameba. Trop. Parasitol. 2016, 6, 8–29. [Google Scholar] [PubMed] [Green Version]
- Arava, S.; Sharma, A.; Iyer, V.K.; Mathur, S.R. Iodamoeba butschlii in a routine cervical smear: Correspondence. Cytopathology 2010, 21, 342–343. [Google Scholar] [CrossRef] [PubMed]
- Schuster, F.L.; Visvesvara, G.S. Free-living amoebae as opportunistic and non-opportunistic pathogens of humans and animals. Int. J. Parasitol. 2004, 34, 1001–1027. [Google Scholar] [CrossRef]
- Reavill, D.R.; Schmidt, R.E. Urinary Tract Diseases of Reptiles. J. Exot. Pet Med. 2010, 19, 280–289. [Google Scholar] [CrossRef]
- Noble, G.A.; Noble, E.R. Entamoebae in farm mammals. J. Parasitol. 1952, 38, 571–595. [Google Scholar] [CrossRef] [PubMed]
- Stensvold, C.R.; Lebbad, M.; Victory, E.L.; Verweij, J.J.; Tannich, E.; Alfellani, M.; Legarraga, P.; Clark, C.G. Increased sampling reveals novel lineages of Entamoeba: Consequences of genetic diversity and host specificity for taxonomy and molecular detection. Protist 2011, 162, 525–541. [Google Scholar] [CrossRef] [Green Version]
- Weynon, C.; O’Connor, F. Human Intestinal Protozoa in the Near East; The Wellcome Bureau of Scientific Research: London, UK, 1917. [Google Scholar]
- Brug, S.L. Endolimax kueneni n.sp., Parasitic in the intestinal tract of the monkey Macacus cynomolgus. Parasitology 1920, 12, 378–379. [Google Scholar] [CrossRef]
- Chiang, S.F. The Rat as a Possible Carrier of the Dysentery Amoeba. Proc. Natl. Acad. Sci. USA 1925, 11, 239–246. [Google Scholar] [CrossRef] [Green Version]
- McFall, C.M. Endolimax reynoldsi nov. sp. from the Intestine of the Common Swift, Sceloporus undulatus. J. Parasitol. 1926, 12, 191. [Google Scholar] [CrossRef]
- Hegner, R.W. Endolimax caviae n. sp. from the Guinea-Pig and Endolimax janisae n. sp. from the Domestic Fowl. J. Parasitol. 1926, 12, 146. [Google Scholar] [CrossRef]
- Lucas, C.L.T. Two new species of amoeba found in cockroaches: With notes on the cysts of nyctotherus ovalis leidy. Parasitology 1927, 19, 223–235. [Google Scholar] [CrossRef]
- Gutierrez-Ballesteros, E.; Wenrich, D.H. Endolimax clevelandi, n. sp., from turtles. J. Parasitol. 1950, 36, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Strassmann, J.E.; Shu, L. Ancient bacteria–amoeba relationships and pathogenic animal bacteria. PLoS Biol. 2017, 15, e2002460. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, G.; La Scola, B.; Abrahão, J. Giant virus vs amoeba: Fight for supremacy. Virol. J. 2019, 16, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bah, A.; Vergne, I. Macrophage autophagy and bacterial infections. Front. Immunol. 2017, 8, 1483. [Google Scholar] [CrossRef] [PubMed]
- Huws, S.A.; McBain, A.J.; Gilbert, P. Protozoan grazing and its impact upon population dynamics in biofilm communities. J. Appl. Microbiol. 2005, 98, 238–244. [Google Scholar] [CrossRef]
- Barbeau, J.; Buhler, T. Biofilms augment the number of free-living amoebae in dental unit waterlines. Res. Microbiol. 2001, 152, 753–760. [Google Scholar] [CrossRef]
- Bigot, R.; Bertaux, J.; Frere, J.; Berjeaud, J.M. Intra-Amoeba Multiplication Induces Chemotaxis and Biofilm Colonization and Formation for Legionella. PLoS ONE 2013, 8, e77875. [Google Scholar] [CrossRef]
- Rakers, S.; Niklasson, L.; Steinhagen, D.; Kruse, C.; Schauber, J.; Sundell, K.; Paus, R. Antimicrobial peptides (AMPs) from fish epidermis: Perspectives for investigative dermatology. J. Investig. Dermatol. 2013, 133, 1140–1149. [Google Scholar] [CrossRef] [Green Version]
- Reverter, M.; Tapissier-Bontemps, N.; Lecchini, D.; Banaigs, B.; Sasal, P. Biological and Ecological Roles of External Fish Mucus: A Review. Fishes 2018, 3, 41. [Google Scholar] [CrossRef] [Green Version]
- Guz, L.; Szczepaniak, K. Intestinal amoebiasis in Heckel discus Symphysodon discus—A case report. Bull. Eur. Assoc. Fish Pathol. 2009, 29, 28–33. [Google Scholar]
- Dyková, I.; Kyselová, I.; Pecková, H.; Oborník, M.; Lukeš, J. Identity of Naegleria strains isolated from organs of freshwater fishes. Dis. Aquat. Organ. 2001, 46, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Dyková, I.; Pecková, H.; Fiala, I.; Dvořáková, H. Fish-isolated Naegleria strains and their phylogeny inferred from ITS and SSU rDNA sequences. Folia Parasitol. 2006, 53, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Taylor, P.W. Isolation and experimental infection of free living amebae in freshwater fishes. J. Parasitol. 1977, 63, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Dyková, I.; Lom, J.; Schroeder-Diedrich, J.M.; Booton, G.C.; Byers, T.J. Acanthamoeba strains isolated from organs of freshwater fishes. J. Parasitol. 1999, 85, 1106–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyková, I.; Lom, J.; Macháčková, B.; Pecková, H. Vexillifera expectata sp. n. and other non-encysting amoebae isolated from organs of freshwater fish. Folia Parasitol. 1998, 45, 17–26. [Google Scholar]
- Dyková, I.; Veverková-Fialová, M.; Fiala, I.; Dvořáková, H. Protacanthamoeba bohemica sp. n., Isolated from the Liver of Tench, Tinca tinca (Linnaeus, 1758). Acta Protozool. 2005, 44, 369–376. [Google Scholar]
- Dyková, I.; Boháčová, L.; Fiala, I.; Macháčková, B.; Pecková, H.; Dvořáková, H. Amoebae of the genera Vannella Bovee, 1965 and Platyamoeba Page, 1969 isolated from fish and their phylogeny inferred from SSU rRNA gene and ITS sequences. Eur. J. Protistol. 2005, 41, 219–230. [Google Scholar] [CrossRef]
- Dyková, I.; Lom, J. Advances in the knowledge of amphizoic amoebae infecting fish. Folia Parasitol. 2004, 51, 81–97. [Google Scholar] [CrossRef] [Green Version]
- English, C.J.; Tyml, T.; Botwright, N.A.; Barnes, A.C.; Wynne, J.W.; Lima, P.C.; Cook, M.T. A diversity of amoebae colonise the gills of farmed Atlantic salmon (Salmo salar) with amoebic gill disease (AGD). Eur. J. Protistol. 2019, 67, 27–45. [Google Scholar] [CrossRef] [Green Version]
- Jensen, H.M.; Karami, A.M.; Mathiessen, H.; Al-Jubury, A.; Kania, P.W.; Buchmann, K. Gill amoebae from freshwater rainbow trout (Oncorhynchus mykiss): In vitro evaluation of antiparasitic compounds against Vannella sp. J. Fish Dis. 2020, 43, 665–672. [Google Scholar] [CrossRef]
- Bermingham, M.L.; Mulcahy, M.F. Neoparamoeba sp. and other protozoans on the gills of Atlantic salmon Salmo salar smolts in seawater. Dis. Aquat. Organ. 2007, 76, 234–240. [Google Scholar] [CrossRef] [Green Version]
- Dyková, I.; Kostka, M.; Wortberg, F.; Nardy, E.; Pecková, H. New data on aetiology of nodular gill disease in rainbow trout, Oncorhynchus mykiss. Folia Parasitol. 2010, 57, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Dyková, I.; Tyml, T. Testate amoeba Rhogostoma minus Belar, 1921, associated with nodular gill disease of rainbow trout, Oncorhynchus mykiss (Walbaum). J. Fish Dis. 2016, 39, 539–546. [Google Scholar] [CrossRef]
- Dyková, I.; Kostka, M.; Pecková, H. Three new species of the amoebozoan genus Vexillifera Schaeffer, 1926. Acta Protozool. 2011, 50, 55–63. [Google Scholar] [CrossRef]
- Sawyer, T.K.; Hnath, J.G.; Conrad, J.F. Thecamoeba hoffmani sp. n. (Amoebida: Thecamoebidae) from gills of fingerling salmonid fish. J. Parasitol. 1974, 60, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Bark, A. A study of the genus Cochliopodium Hertwig and Lesser 1874. Protistologica 1973, 9, 119–138. [Google Scholar]
- Page, F. A New Key to Freshwater and Soil Gymnamoebae with Instructions for Culture; Freshwater Biological Association: Ambleside, UK, 1988. [Google Scholar]
- Dyková, I.; Lom, J.; Macháčková, B. Cochliopodium minus, a scale-bearing amoeba isolated from organs of perch Perca fluviatilis. Dis. Aquat. Organ. 1998, 34, 205–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyková, I.; Kostka, M.; Pecková, H. Morphology and SSU rDNA-based phylogeny of a new strain of Saccamoeba sp. [Saccamoeba Frenzel, 1892, Amoebozoa]. Acta Protozool. 2008, 47, 397–405. [Google Scholar]
- Dyková, I.; Veverková, M.; Fiala, I.; Macháèková, B. A Free-Living Amoeba with Unusual Pattern of Mitochondrial Structure Isolated from Atlantic Salmon, Salmo salar L. Acta Protozool. 2002, 41, 415–419. [Google Scholar]
- Dyková, I.; Kostka, M. Illustrated Guide to Culture Collection of Free-Living Amoebae; Academia: Praha, Czech Republic, 2013. [Google Scholar]
- Dyková, I.; Novoa, B. Comments on diagnosis of amoebic gill disease (AGD) in turbot, Scophthalmus maximus. Bull. Eur. Ass. Fish Pathol. 2001, 21, 40–44. [Google Scholar]
- Dyková, I.; Veverková, M.; Fiala, I.; Macháčková, B.; Pecková, H. Nuclearia pattersoni sp. n. (Filosea), a new species of amphizoic amoeba isolated from gills of roach (Rutilus rutilus), and its rickettsial endosymbiont. Folia Parasitol. 2003, 50, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Howard, T. Paramoebiasis of Sea-Farmed Salmonids in Tasmania: A Study of Its Aetiology, Pathogenicity, and Control. Ph.D. Thesis, University of Tasmania, Hobart, Australia, 2001. [Google Scholar]
- Boerlage, A.S.; Ashby, A.; Herrero, A.; Reeves, A.; Gunn, G.J.; Rodger, H.D. Epidemiology of marine gill diseases in Atlantic salmon (Salmo salar) aquaculture: A review. Rev. Aquac. 2020, 12, 2140–2159. [Google Scholar] [CrossRef]
- Marcos-López, M.; Rodger, H.D. Amoebic gill disease and host response in Atlantic salmon (Salmo salar L.): A review. Parasite Immunol. 2020, 42, e12766. [Google Scholar] [CrossRef]
- Rodger, H.D. Amoebic gill disease in farmed halibut (Hippoglossus hippoglossus) in the United Kingdom. Vet. Rec. Case Rep. 2019, 7, e000797. [Google Scholar] [CrossRef]
- Padrós, F.; Constenla, M.; Zarza, C. Pathology and Diseases Control. In The Biology of Sole; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2019; pp. 326–353. [Google Scholar]
- Athanassopoulou, F.; Cawthorn, R.; Lytra, K. Amoeba-like Infections in Cultured Marine Fishes: Systemic Infection in Pompano Trachinotus falcatus L. from Singapore and Gill Disease Associated with Paramoeba sp. in Sea Bream Sparus aurata L. from Greece. J. Vet. Med. Ser. B 2002, 49, 411–412. [Google Scholar] [CrossRef]
- Kim, W.S.; Kong, K.H.; Kim, J.O.; Jung, S.J.; Kim, J.H.; Oh, M.J. Amoebic gill disease outbreak in marine fish cultured in Korea. J. Vet. Diagn. Investig. 2017, 29, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.J.; Cavaleiro, F.; Campos, P.; Sousa, A.; Teixeira, F.; Martins, M. Impact of amoeba and scuticociliatidia infections on the aquaculture European sea bass (Dicentrarchus labrax L.) in Portugal. Vet. Parasitol. 2010, 171, 15–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crosbie, P.B.B.; Ogawa, K.; Nakano, D.; Nowak, B.F. Amoebic gill disease in hatchery-reared ayu, Plecoglossus altivelis (Temminck & Schlegel), in Japan is caused by Neoparamoeba perurans. J. Fish Dis. 2010, 33, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Karlsbakk, E.; Olsen, A.B.; Einen, A.C.B.; Mo, T.A.; Fiksdal, I.U.; Aase, H.; Kalgraff, C.; Skår, S.Å.; Hansen, H. Amoebic gill disease due to Paramoeba perurans in ballan wrasse (Labrus bergylta). Aquaculture 2013, 412–413, 41–44. [Google Scholar] [CrossRef] [Green Version]
- Haugland, G.T.; Olsen, A.B.; Rønneseth, A.; Andersen, L. Lumpfish (Cyclopterus lumpus L.) develop amoebic gill disease (AGD) after experimental challenge with Paramoeba perurans and can transfer amoebae to Atlantic salmon (Salmo salar L.). Aquaculture 2017, 478, 48–55. [Google Scholar] [CrossRef]
- Oldham, T.; Rodger, H.; Nowak, B.F. Incidence and distribution of amoebic gill disease (AGD)—An epidemiological review. Aquaculture 2016, 457, 35–42. [Google Scholar] [CrossRef]
- Kent, M.; Sawyer, T.; Hedrick, R. Paramoeba pemaquidensis (Sarcomastigophora: Paramoebidae) infestation of the gills of coho salmon Oncorhynchus kisutch reared in sea water. Dis. Aquat. Organ. 1988, 5, 163–169. [Google Scholar] [CrossRef]
- Munday, B.L.; Foster, C.K.; Roubal, F.R.; Lester, R.J.G. Paramoebic gill infection and associated pathology of Atlantic salmon, Salmo salar, and rainbow trout, Salmo gairdneri, in Tasmania. In Pathology in Marine Science; Academic Press Inc.: San Diego, CA, USA, 1990; pp. 215–222. [Google Scholar]
- Dyková, I.; Nowak, B.F.; Crosbie, P.B.B.; Fiala, I.; Pecková, H.; Adams, M.B.; Macháčková, B.; Dvořáková, H. Neoparamoeba branchiphila n. sp., and related species of the genus Neoparamoeba Page, 1987: Morphological and molecular characterization of selected strains. J. Fish Dis. 2005, 28, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Young, N.; Dyková, I.; Snekvik, K.; Nowak, B.; Morrison, R. Neoparamoeba perurans is a cosmopolitan aetiological agent of amoebic gill disease. Dis. Aquat. Organ. 2008, 78, 217–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leef, M.J.; Harris, J.O.; Powell, M.D. Respiratory pathogenesis of amoebic gill disease (AGD) in experimentally infected Atlantic salmon Salmo salar. Dis. Aquat. Organ. 2005, 66, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Adams, M.B.; Nowak, B.F. Sequential pathology after initial freshwater bath treatment for amoebic gill disease in cultured Atlantic salmon, Salmo salar L. J. Fish Dis. 2004, 27, 163–173. [Google Scholar] [CrossRef]
- Chang, Y.C.; Hamlin-Wright, H.; Monaghan, S.; Herath, T.; Baily, J.; del Pozo, J.; Downes, J.; Preston, A.; Chalmers, L.; Jayasuriya, N.; et al. Changes in distribution, morphology and ultrastructure of chloride cell in Atlantic salmon during an AGD infection. J. Fish Dis. 2019, 42, 1433–1446. [Google Scholar] [CrossRef]
- Valdenegro-Vega, V.A.; Crosbie, P.; Bridle, A.; Leef, M.; Wilson, R.; Nowak, B.F. Differentially expressed proteins in gill and skin mucus of Atlantic salmon (Salmo salar) affected by amoebic gill disease. Fish Shellfish Immunol. 2014, 40, 69–77. [Google Scholar] [CrossRef]
- Wynne, J.W.; O’Sullivan, M.G.; Stone, G.; Cook, M.T.; Nowak, B.F.; Lovell, D.R.; Taylor, R.S.; Elliott, N.G. Resistance to amoebic gill disease (AGD) is characterised by the transcriptional dysregulation of immune and cell cycle pathways. Dev. Comp. Immunol. 2008, 32, 1539–1560. [Google Scholar] [CrossRef]
- MacPhail, D.; Koppenstein, R.; Longshaw, M.; Henriquez, F. The bacterial microbiome of in vitro cultures of Paramoeba perurans. Access Microbiol. 2019, 1, 686. [Google Scholar] [CrossRef]
- Taylor, R.; Huynh, C.; Cameron, D.; Evans, B.; Cook, M.; Ritchie, G. Gill Score Guide—Amoebic Gill Disease (AGD) Management Training Document; Tassal Operations Pty: Hobart, Australia, 2016. [Google Scholar]
- Herrero, A.; Thompson, K.D.; Ashby, A.; Rodger, H.D.; Dagleish, M.P. Complex Gill Disease: An Emerging Syndrome in Farmed Atlantic Salmon (Salmo salar L.). J. Comp. Pathol. 2018, 163, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Downes, J.K.; Rigby, M.L.; Taylor, R.S.; Maynard, B.T.; MacCarthy, E.; O’Connor, I.; Marcos-Lopez, M.; Rodger, H.D.; Collins, E.; Ruane, N.M.; et al. Evaluation of Non-destructive Molecular Diagnostics for the Detection of Neoparamoeba perurans. Front. Mar. Sci. 2017, 4, 61. [Google Scholar] [CrossRef] [Green Version]
- Florent, R.L.; Becker, J.A.; Powell, M.D. Efficacy of bithionol as an oral treatment for amoebic gill disease in Atlantic salmon Salmo salar (L.). Aquaculture 2007, 270, 15–22. [Google Scholar] [CrossRef]
- Lazado, C.; Timmerhaus, G.; Pedersen, L.-F.; Pittman, K.; Soleng, M.; Haddeland, S.; Johansen, L.-H.; Breiland, M.; Rørmark, L.; Mohammed, S.; et al. Peracetic Acid as a Potential Treatment for Amoebic Gill Disease (AGD) in Atlantic Salmon—Stage 1; Nofima Marked. Rapport No. 21/2019; Nofima: Tromsø, Norway, 2019. [Google Scholar]
- Powell, M.D.; Reynolds, P.; Kristensen, T. Freshwater treatment of amoebic gill disease and sea-lice in seawater salmon production: Considerations of water chemistry and fish welfare in Norway. Aquaculture 2015, 448, 18–28. [Google Scholar] [CrossRef] [Green Version]
- Martinsen, K.H.; Thorisdottir, A.; Lillehammer, M. Effect of hydrogen peroxide as treatment for amoebic gill disease in Atlantic salmon (Salmo salar L.) in different temperatures. Aquac. Res. 2018, 49, 1733–1739. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, R.W.; El-Matbouli, M.; Fischer-Scherl, T. A proliferative gill disease (PGD) in rainbow trout (Oncorhynchus mykiss). Bull. Eur. Assoc. Fish Pathol. 1992, 12, 139–141. [Google Scholar]
- Lovy, J.; Goodwin, A.E.; Speare, D.J.; Wadowska, D.W.; Wright, G.M. Histochemical and ultrastructural analysis of pathology and cell responses in gills of channel catfish affected with proliferative gill disease. Dis. Aquat. Organ. 2011, 94, 125–134. [Google Scholar] [CrossRef]
- Daoust, P.-Y.; Ferguson, H. Potential for recovery in nodular gill disease of rainbow trout, Salmo gairdneri Richardson. J. Fish Dis. 1986, 9, 313–318. [Google Scholar] [CrossRef]
- Speare, D.J.; Ferguson, H.W. Clinical and pathological features of common gill diseases of cultured salmonids in Ontario. Can. Vet. J. Rev. Vet. Can. 1989, 30, 882–887. [Google Scholar]
- Speare, D.J. Nodular gill disease (amoebic gill infestation) in Arctic char, Salvelinus alpinus. J. Comp. Pathol. 1999, 121, 277–282. [Google Scholar] [CrossRef]
- Perolo, A.; Gustinelli, A.; Fioravanti, M.L.; Manfrin, A.; Dalla Pozza, M.; Lunelli, F.; Accini, A.; Quaglio, F. Occurrence of nodular gill disease in farmed brown trout (Salmo trutta L.). J. Fish Dis. 2019, 42, 1315–1320. [Google Scholar] [CrossRef]
- Tubbs, L.; Wybourne, B.A.; Lumsden, J.S.; Lumsden, J.S. Nodular gill disease causing proliferative branchitis and mortality in chinook salmon (Oncorhynchus tshawytscha). N. Z. Vet. J. 2010, 58, 59–61. [Google Scholar] [CrossRef]
- Quaglio, F.; Perolo, A.; Bronzatti, P.; Gustinelli, A.; Menconi, V.; Cavazza, G.; Caffara, M.; Manfrin, A.; Gallo, E.; Fioravanti, M.L. Nodular gill disease in farmed rainbow trout (Oncorhynchus mykiss) in Italy. J. Fish Dis. 2016, 39, 1139–1142. [Google Scholar] [CrossRef] [PubMed]
- Voelker, F.A.; Anver, M.R.; McKee, A.E.; Casey, H.W.; Brenniman, G.R. Amebiasis in Goldfish. Vet. Pathol. 1977, 14, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Lom, J.; Dyková, I. Protozoan Parasites of Fishes; Elsevier Science Publishers: Amsterdam, The Netherlands, 1992. [Google Scholar]
- Steinhagen, D.; Jendrysek, S.; Körting, W. Amöbiasis bei Goldfishen. Kleintierpraxis 1993, 38, 469–474. [Google Scholar]
- Dyková, I.; Lom, J.; Machácková, B.; Sawyer, T.K. Amoebic infections in goldfishes and granulomatous lesions. Folia Parasitol. 1996, 43, 80–91. [Google Scholar]
- Anderson, I.; Prior, H.; Rodwell, B.; Harris, G. Iridovirus-like virions in imported dwarf gourami (Colisa lalia) with systemic amoebiasis. Aust. Vet. J. 1993, 70, 66–67. [Google Scholar] [CrossRef]
- Nash, G.; Nash, M.; Schlotfeldt, H.-J. Systemic amoebiasis in cultured European catfish, Silurus glanis L. J. Fish Dis. 1988, 11, 57–71. [Google Scholar] [CrossRef]
- Constenla, M.; Padrós, F. Histopathological and ultrastructural studies on a novel pathological condition in Solea senegalensis. Dis. Aquat. Organ. 2010, 90, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Palíková, M.; Navrátil, S.; Dyková, I.; Pavlík, I.; Slaný, M.; Tichý, F.; Novotný, L.; Zendulková, D.; Laichmanová, M.; Mareš, J. Archamoeba infection manifested by in European tench, Tinca tinca (L.). Bull. Eur. Assoc. Fish Pathol. 2012, 32, 174–180. [Google Scholar]
- Cavalier-Smith, T. A revised six-kingdom system of life. Biol. Rev. Camb. Philos. Soc. 1998, 73, 203–266. [Google Scholar] [CrossRef] [PubMed]
- Simitch, T. Sur une amibe buccale du chien Entamœba canibuccalis n. sp. Ann. Parasitol. Hum. Comp. 1938, 16, 251–253. [Google Scholar] [CrossRef]
- Rodhain, J.; Van Hoof, M. Entamoeba knowlesi n.sp. parasite de deux tortues: Terrapina cinosternoïdes et Platysternum megacephalum. Ann. Parasitol. Hum. Comp. 1947, 22, 129–137. [Google Scholar] [CrossRef]
- Alexeieff, A. Sur les charactères cytologiques et la systématique des amibes du groupe limax (Naegleria nov. gen. et Hartmannia nov. gen.) et des amibes parasites des vertebrates (Protamoeba nov. gen.). Bull. Soc. Zool. Fr. 1912, 37, 55–74. [Google Scholar] [CrossRef]
- Bullock, W.L. Entamoeba gadi sp. n. from the rectum of the pollock, Pollachius virens (L., 1758), with some observations on its cytochemistry. J. Parasitol. 1966, 52, 679–684. [Google Scholar] [CrossRef] [PubMed]
- Noble, E.; Noble, G. Amebic Parasites of Fishes. J. Protozool. 1966, 13, 478–480. [Google Scholar] [CrossRef]
- Da Cunha, A.; Pendio, J. Entamoeba pimelodi n.sp., parasite d’un poisson d’eau douce. C. R. Biol. 1926, 95, 1010–1011. [Google Scholar]
- Chen, C. The protozoan parasites from four species of chinese pond fishes: Ctenopharyngodon idellus, Mylopharingodon piceus, Aristichthys nobilis and Hypophthalmichthys molithrix III.The Protozoan Parasites of Aristichthys nobillis and Hypophthalmichthys moli. Acta Hydrobiol. Sin. 1956, 1, 123–164. [Google Scholar]
- Jinatham, V.; Popluechai, S.; Clark, C.G.; Gentekaki, E. Entamoeba chiangraiensis n. sp. (Amoebozoa: Entamoebidae) isolated from the gut of Asian swamp eel (Monopterus albus) in northern Thailand. Parasitology 2019, 146, 1719–1724. [Google Scholar] [CrossRef] [Green Version]
- Silberman, J.D.; Clark, C.G.; Diamond, L.S.; Sogin, M.L. Phylogeny of the genera Entamoeba and Endolimax as deduced from small-subunit ribosomal RNA sequences. Mol. Biol. Evol. 1999, 16, 1740–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constenla, M. Contributions to the Knowledge of a New Disease Caused by an Amoeba in Ongrowing Senegalese Sole, Solea Senegalensis (Kaup 1858). Ph.D. Thesis, Universitat Autònoma de Barcelona, Bellaterra, Spain, 2013. [Google Scholar]
- Constenla, M.; Padrós, F.; Palenzuela, O. Endolimax piscium sp. nov. (Amoebozoa), causative agent of systemic granulomatous disease of cultured sole, Solea senegalensis Kaup. J. Fish Dis. 2014, 37, 229–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constenla, M.; Padrós, F.; del Pozo, R.; Palenzuela, O. Development of different diagnostic techniques for Endolimax piscium (archamoebae) and their applicability in Solea senegalensis clinical samples. J. Fish Dis. 2016, 39, 1433–1443. [Google Scholar] [CrossRef] [PubMed]
- Constenla, M.; Padrós, F.; Villanueva-González, A.; Del Pozo, R.; Palenzuela, O. Horizontal transmission of Endolimax piscium, causative agent of systemic amoebiasis in Senegalese sole Solea senegalensis. Dis. Aquat. Organ. 2018, 130, 235–240. [Google Scholar] [CrossRef]
- Pérez-Tamayo, R. Pathology of amebiasis. In Amebiasis; Martinez-Palomo, A., Ed.; Elsevier: New York, NY, USA, 1986. [Google Scholar]
- Costa, C.A.; Nunes, Á.C.; Ferreira, A.J.; Gomes, M.A.; Caliari, M.V. Entamoeba histolytica and E. dispar trophozoites in the liver of hamsters: In vivo binding of antibodies and complement. Parasites Vectors 2010, 3, 23. [Google Scholar] [CrossRef] [Green Version]
- Noble, A.C.; Herman, R.L.; Noga, E.J.; Bullock, G.L. Recurrent Amoebic Gill Infestation in Rainbow Trout Cultured in a Semiclosed Water Recirculation System. J. Aquat. Anim. Health 1997, 9, 64–69. [Google Scholar] [CrossRef]
- Visvesvara, G.S.; Moura, H.; Schuster, F.L. Pathogenic and opportunistic free-living amoebae: Acanthamoeba spp., Balamuthia mandrillaris, Naegleria fowleri, and Sappinia diploidea. FEMS Immunol. Med. Microbiol. 2007, 50, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Coulon, C.; Collignon, A.; McDonnell, G.; Thomas, V. Resistance of Acanthamoeba cysts to disinfection treatments used in health care settings. J. Clin. Microbiol. 2010, 48, 2689–2697. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Gills affected by amoebic gill disease (AGD) and nodular gill disease (NGD): (A), gill filaments of turbot affected by amoebic gill disease. Note the typical white multifocal lesions. (B–D), Paraffin-embedded histological sections: (B), extensive hyperplasia of epithelial gill cells in Atlantic salmon, with synechiae and lacunae between filaments; (C), detail of amoebae within lacunae between gill filaments in AGD in Atlantic salmon; (D), flattened amoebae attached to the surface of proliferative gill lesions of NGD in rainbow trout; (E), transmission electron micrographs of gill with amoebae attached to the epithelium in NGD in rainbow trout.
Figure 1.
Gills affected by amoebic gill disease (AGD) and nodular gill disease (NGD): (A), gill filaments of turbot affected by amoebic gill disease. Note the typical white multifocal lesions. (B–D), Paraffin-embedded histological sections: (B), extensive hyperplasia of epithelial gill cells in Atlantic salmon, with synechiae and lacunae between filaments; (C), detail of amoebae within lacunae between gill filaments in AGD in Atlantic salmon; (D), flattened amoebae attached to the surface of proliferative gill lesions of NGD in rainbow trout; (E), transmission electron micrographs of gill with amoebae attached to the epithelium in NGD in rainbow trout.

Figure 2.
Senegalese soles clinically infected by Endolimax piscium: (A), conspicuous lump from the abdominal cavity, and noticeable at the skin surface (arrow); (B), nodule in muscle with an abscess-like aspect with a soft and liquefied consistency. (C–E), paraffin-embedded histological sections: (C), skeletal muscle showing the typical granulomatous inflammatory reaction to E. piscium; (D), E. piscium cells within the intestinal epithelium (arrow); (E), in situ hybridisation of histological sections where parasite cells can be recognised stained in purple.
Figure 2.
Senegalese soles clinically infected by Endolimax piscium: (A), conspicuous lump from the abdominal cavity, and noticeable at the skin surface (arrow); (B), nodule in muscle with an abscess-like aspect with a soft and liquefied consistency. (C–E), paraffin-embedded histological sections: (C), skeletal muscle showing the typical granulomatous inflammatory reaction to E. piscium; (D), E. piscium cells within the intestinal epithelium (arrow); (E), in situ hybridisation of histological sections where parasite cells can be recognised stained in purple.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Padrós, F.; Constenla, M. Diseases Caused by Amoebae in Fish: An Overview. Animals 2021, 11, 991. https://doi.org/10.3390/ani11040991
AMA Style
Padrós F, Constenla M. Diseases Caused by Amoebae in Fish: An Overview. Animals. 2021; 11(4):991. https://doi.org/10.3390/ani11040991
Chicago/Turabian StylePadrós, Francesc, and Maria Constenla. 2021. "Diseases Caused by Amoebae in Fish: An Overview" Animals 11, no. 4: 991. https://doi.org/10.3390/ani11040991
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.