
Differences in Intestinal Barrier Development between Intrauterine Growth Restricted and Normal Birth Weight Piglets

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Tissue Collection
2.2. Histological Staining
2.3. Optical Microscopy Analysis
2.4. Scanning Electron Microscopy
2.5. Statistical Analysis
3. Results
3.1. Pig Body Weight
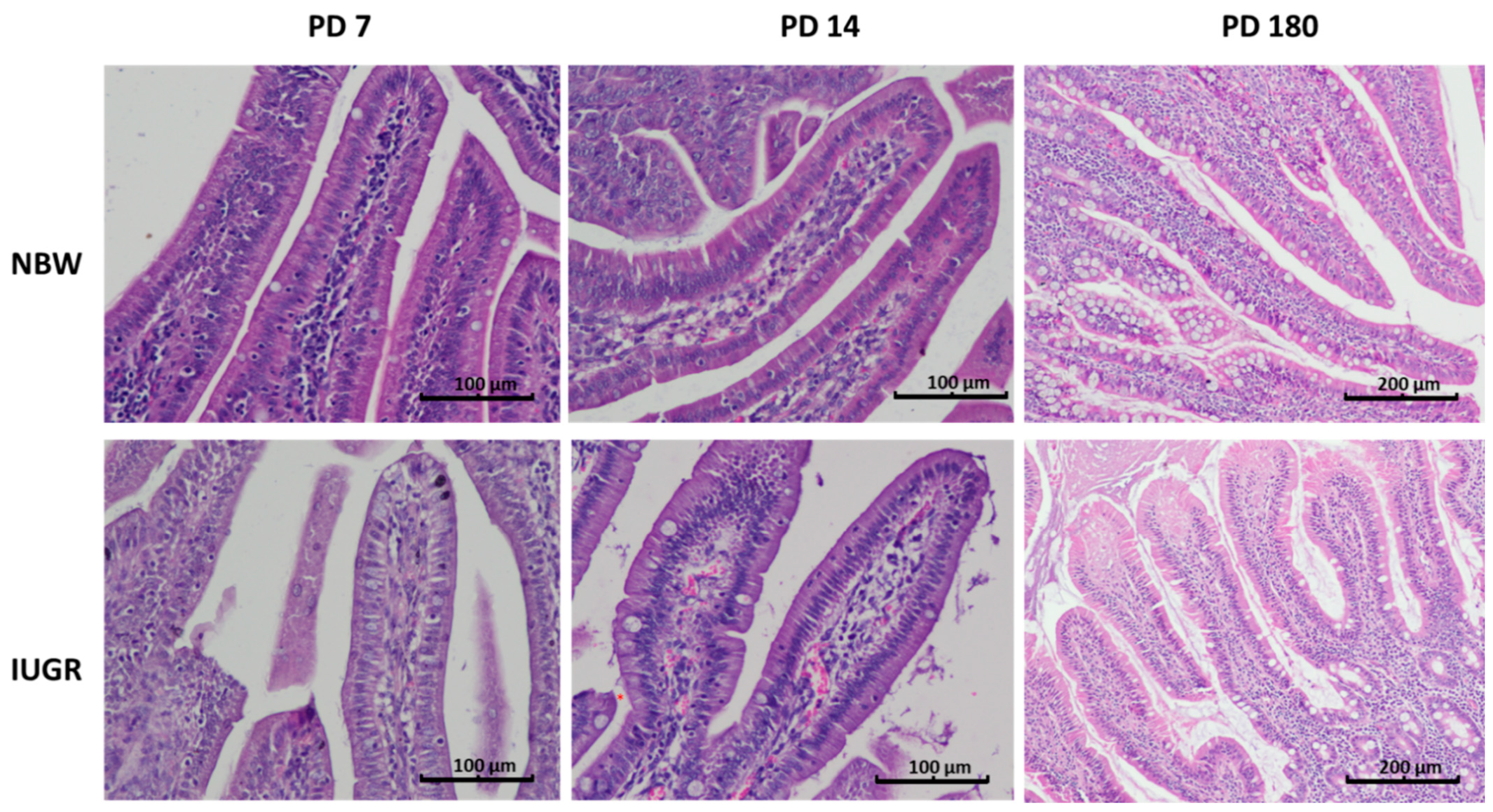
3.2. Disappearance of Foetal-Type Type Enterocytes as a Marker of Intestinal Mucosa Development
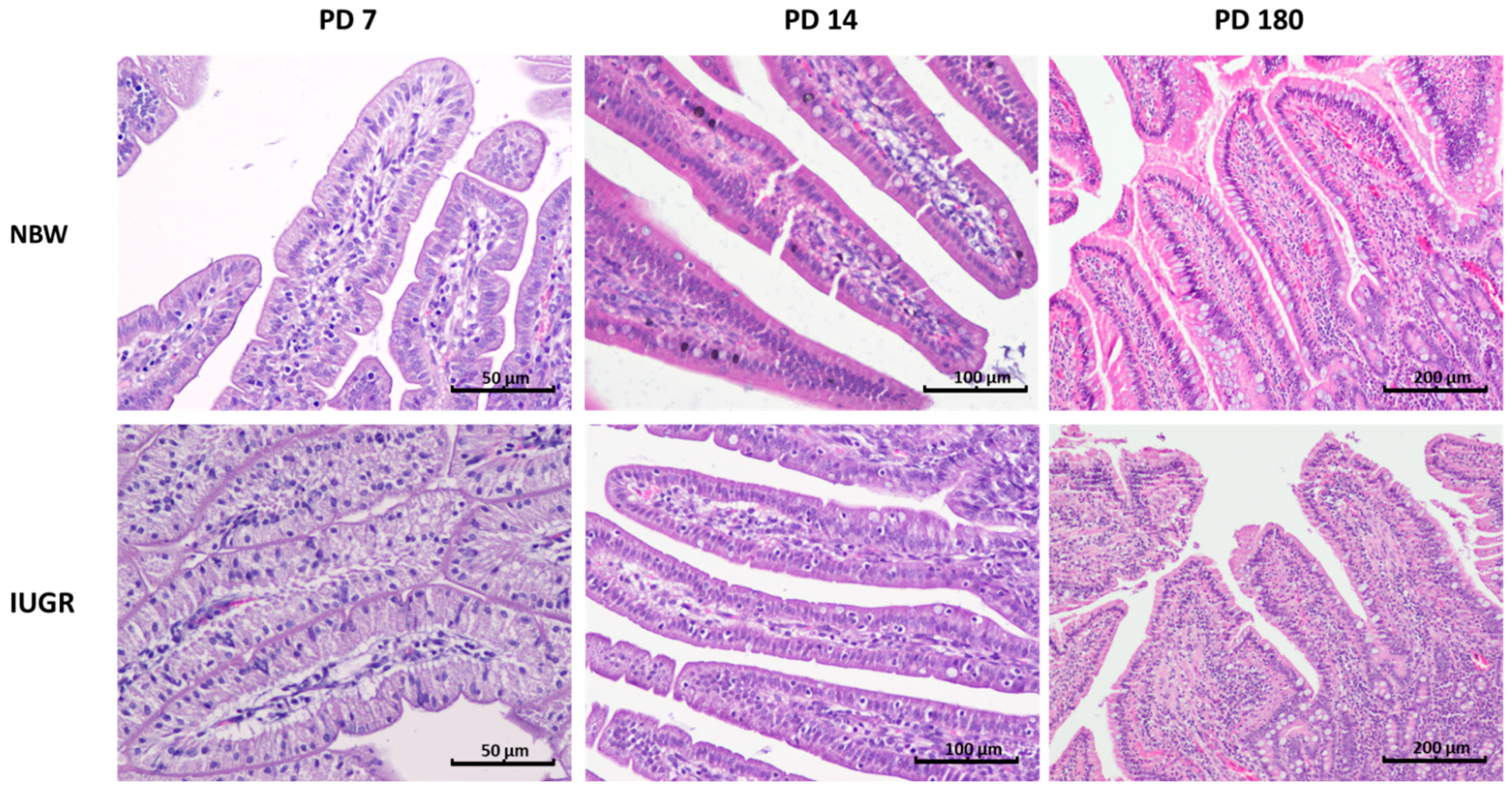
3.3. Intraepithelial Leukocytes
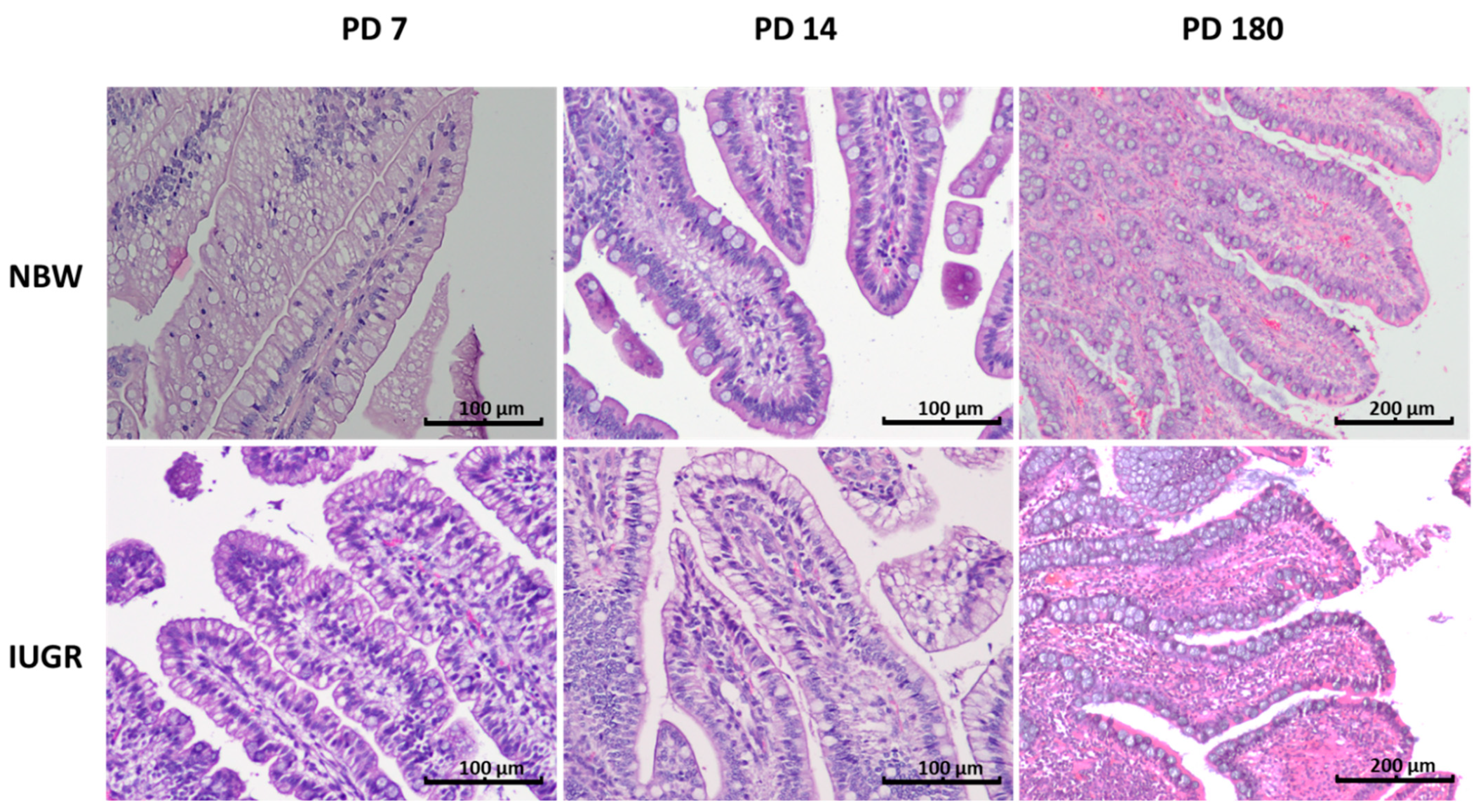
3.4. Goblet Cells
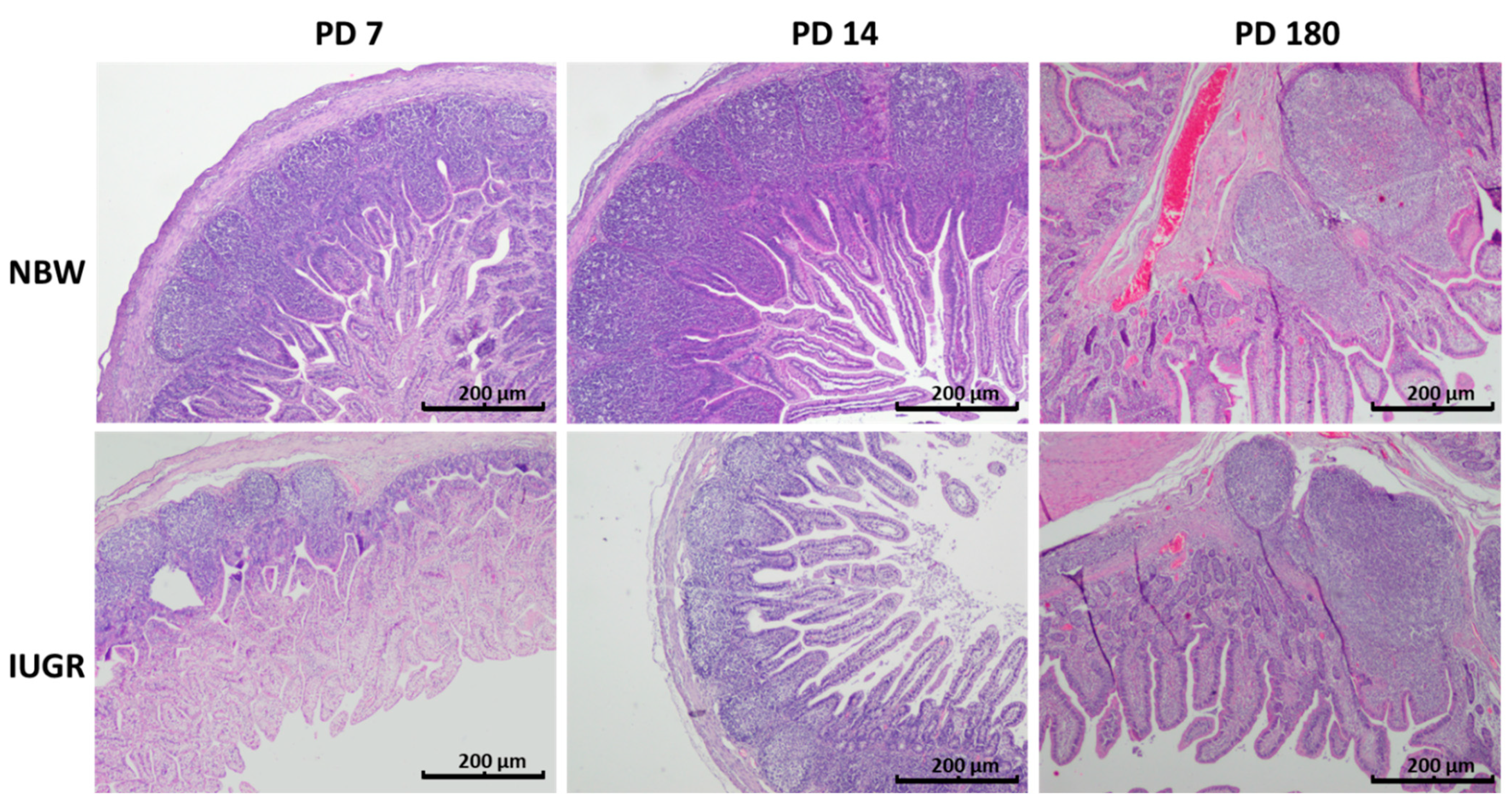
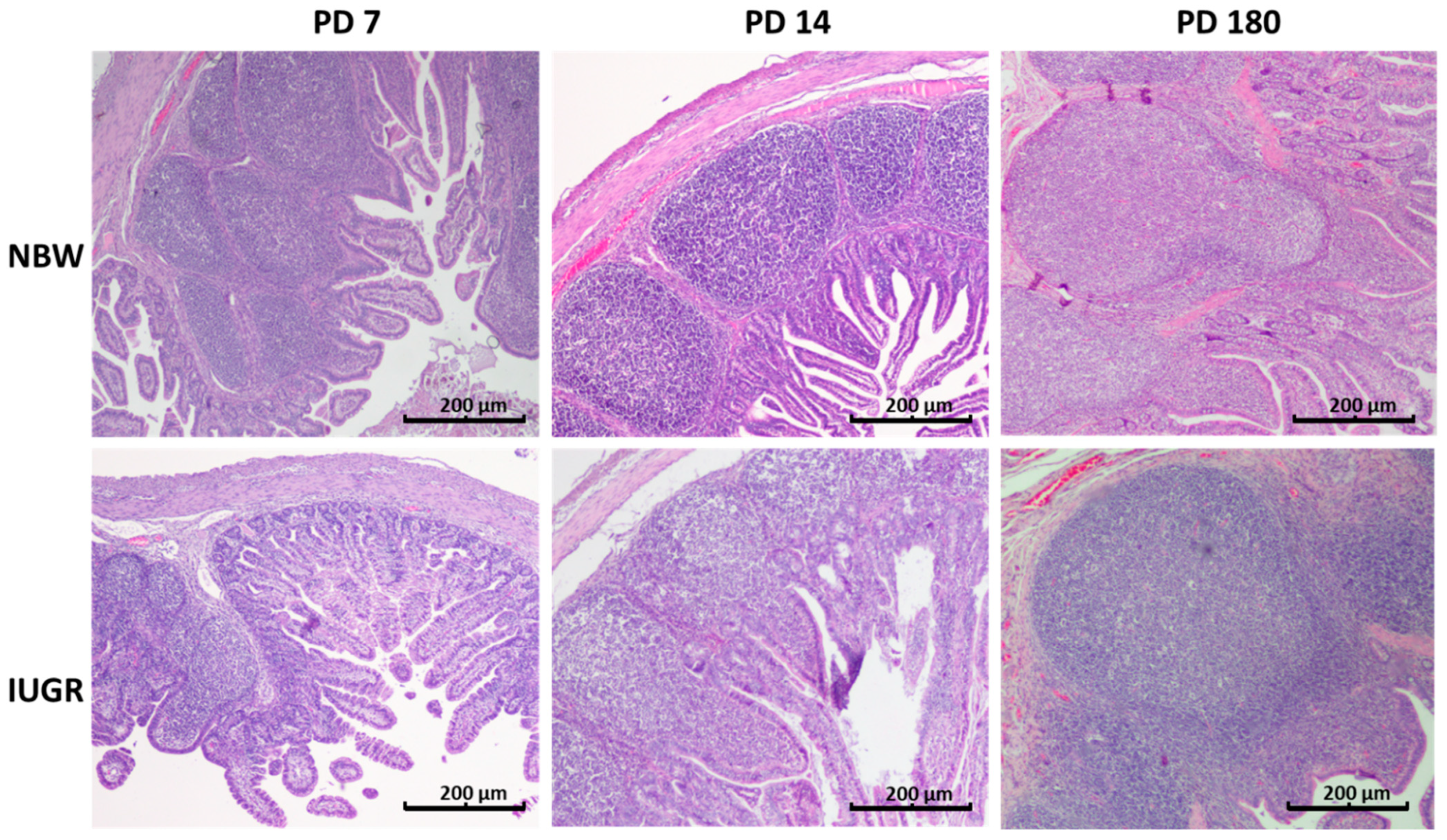
3.5. Peyer’s Patches
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferenc, K.; Pietrzak, P.; Godlewski, M.M.; Piwowarski, J.; Kiliańczyk, R.; Guilloteau, P.; Zabielski, R. Intrauterine growth retarded piglet as a model for humans—Studies on the perinatal development of the gut structure and function. Reprod. Biol. 2014, 14, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Matyba, P.; Florowski, T.; Dasiewicz, K.; Ferenc, K.; Olszewski, J.; Trela, M.; Galemba, G.; Słowiński, M.; Sady, M.; Domańska, D.; et al. Performance and Meat Quality of Intrauterine Growth Restricted Pigs. Animals 2021, 11, 254. [Google Scholar] [CrossRef]
- Ferenc, K.; Pilžys, T.; Skrzypek, T.; Garbicz, D.; Marcinkowski, M.; Dylewska, M.; Gładysz, P.; Skorobogatov, O.; Gajewski, Z.; Grzesiuk, E.; et al. Structure and Function of Enterocyte in Intrauterine Growth Retarded Pig Neonates. Dis. Markers 2017, 3, 1–9. [Google Scholar] [CrossRef]
- Mickiewicz, M.; Zabielski, R.; Grenier, B.; Le Normand, L.; Savary, G.; Holst, J.J.; Oswald, I.P.; Metges, C.C.; Guilloteau, P. Structural and functional development of small intestine in intrauterine growth retarded porcine offspring born to gilts fed diets with differing protein ratios throughout pregnancy. J. Physiol. Pharmacol. 2012, 63, 225–239. [Google Scholar]
- Skrzypek, T.; Valverde Piedra, J.L.; Skrzypek, H.; Kazimierczak, W.; Biernat, M.; Zabielski, R. Gradual disappearance of vacuolated enterocytes in the small intestine of neonatal piglets. J. Physiol. Pharmacol. 2007, 58, 87–95. [Google Scholar] [PubMed]
- Skrzypek, T.H.; Kazimierczak, W.; Skrzypek, H.; Valverde Piedra, J.L.; Godlewski, M.M.; Zabielski, R. Mechanisms involved in the development of the small intestine mucosal layer in postnatal piglets. J. Physiol. Pharmacol. 2018, 69, 127–138. [Google Scholar] [CrossRef]
- Skrzypek, T.; Szymańczyk, S.; Ferenc, K.; Kazimierczak, W.; Szczepaniak, K.; Zabielski, R. The contribution of vacuolated foetal-type enterocytes in the process of maturation of the small intestine in piglets. Invited review. J. Anim. Feed Sci. 2018, 27, 187–201. [Google Scholar] [CrossRef]
- Ferenc, K.; Pietrzak, P.; Wierzbicka, M.; Matyba, P.; Grzesiuk, E.; Gajewski, Z.; Zabielski, R. Alterations in the liver of intrauterine growth retarded piglets may predispose to development of insulin resistance and obesity in later life. J. Physiol. Pharmacol. 2018, 69. [Google Scholar] [CrossRef]
- Dong, L.; Zhong, X.; Ahmad, H.; Li, W.; Wang, Y.; Zhang, L.; Wang, T. Intrauterine growth restriction impairs small intestinal mucosal immunity in neonatal piglets. J. Histochem. Cytochem. 2014, 62, 510–518. [Google Scholar] [CrossRef] [PubMed]
- D’Inca, R.; Kloareg, M.; Gras-Le Guen, C.; Le Huerou-Luron, I. Intrauterine growth restriction modifies the developmental pattern of intestinal structure. transcriptomic profile. and bacterial colonization in neonatal pigs. J. Nutr. 2010, 140, 925–931. [Google Scholar] [CrossRef]
- Tao, S.; Zhou, T.; Saelao, P.; Wang, Y.; Zhu, Y.; Li, T.; Zhou, H.; Wang, J. Intrauterine growth restriction alters the genome-wide DNA methylation profiles in small intestine. liver and longissimus dorsi muscle of newborn piglets. Curr. Protein Pept. Sci. 2019, 20, 713–726. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Liu, C.; Li, N.; Wu, Z.; Li, T.; Han, D.; Li, Z.; Zhao, J.; Wang, J. Membrane proteomic analysis reveals the intestinal development is deteriorated by intrauterine growth restriction in piglets. Funct. Integr. Genom. 2020, 20, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Degroote, J.; Van Ginneken, C.; Van Poucke, M.; Vergauwen, H.; Dam, T.M.T.; Vanrompay, D.; Peelman, L.J.; De Smet, S.; Michiels, J. Intrauterine growth restriction in neonatal piglets affects small intestinal mucosal permeability and mRNA expression of redox-sensitive genes. FASEB J. 2016, 30, 863–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Wang, W.; Yuan, T.; Fu, L.; Zhou, L.; Lin, G.; Zhao, S.; Zhuo, H.; Wu, G.; Wang, J. MicroRNA-29a mediates the impairment of intestinal epithelial integrity induced by intrauterine growth restriction in pig. Am. J. Physiol. -Gastrointest. Liver Physiol. 2017, 312, 434–442. [Google Scholar] [CrossRef]
- Amdi, C.; Krogh, U.; Flummer, C.; Oksbjerg, N.; Hansen, C.F.; Theil, P.K. Intrauterine growth restricted piglets defined by their head shape ingest insufficient amounts of colostrum. J. Anim. Sci. 2013, 91, 5605–5613. [Google Scholar] [CrossRef] [Green Version]
- Hales, J.; Moustsen, V.A.; Nielsen, M.B.; Hansen, C.F. Individual physical characteristics of neonatal piglets affect preweaning survival of piglets born in a noncrated system. J. Anim. Sci. 2013, 91, 4991–5003. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Liu, Y.; Yan, C.; Peng, X.; Xu, Q.; Xuan, Y.; Han, F.; Gang, T.; Zhengfeng, F.; Yan, L.; et al. Postnatal nutritional restriction affects growth and immune function of piglets with intra-uterine growth restriction. Br. J. Nutr. 2015, 114, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Chen, Y.; Li, Y.; Zhang, T.; Ying, Z.; Su, W.; Zhang, L.; Wang, T. l-Threonine improves intestinal mucin synthesis and immune function of intrauterine growth–retarded weanling piglets. Nutrition 2019, 59, 182–187. [Google Scholar] [CrossRef]
- Wang, X.; Wu, W.; Lin, G.; Li, D.; Wu, G.; Wang, J. Temporal Proteomic Analysis Reveals Continuous Impairment of Intestinal Development in Neonatal Piglets with Intrauterine Growth Restriction. J. Proteome Res. 2010, 9, 924–935. [Google Scholar] [CrossRef]
- Huang, S.; Li, N.; Liu, C.; Li, T.; Wang, W.; Jiang, L.; Li, Z.; Han, D.; Tao, S.; Wang, J. Characteristics of the gut microbiota colonization, inflammatory profile, and plasma metabolome in intrauterine growth restricted piglets during the first 12 hours after birth. J. Microbiol. 2019, 57, 748–758. [Google Scholar] [CrossRef]
- Amdi, C.; Lynegaard, J.C.; Thymann, T.; Williams, A.R. Intrauterine growth restriction in piglets alters blood cell counts and impairs cytokine responses in peripheral mononuclear cells 24 days post-partum. Sci. Rep. 2020, 10, 4683. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Chen, Y.; Li, Y.; Wang, T. Protective Effect of Polydatin on Jejunal Mucosal Integrity, Redox Status, Inflammatory Response, and Mitochondrial Function in Intrauterine Growth-Retarded Weanling Piglets. Oxidative Med. Cell. Longev. 2020, 7178123. [Google Scholar] [CrossRef]
- Inoue, R.; Tsukahara, T.; Nakatani, M.; Okutani, M.; Nishibayashi, R.; Ogawa, S.; Harayama, T.; Nagino, T.; Hatanaka, H.; Fukuta, K.; et al. Weaning Markedly Affects Transcriptome Profiles and Peyer’s Patch Development in Piglet Ileum. Front. Immunol. 2015, 6, 630. [Google Scholar] [CrossRef] [Green Version]
- Butler, J.E.; Rainard, P.; Lippolis, J.; Salmon, H.; Kacskovics, I. The Mammary Gland in Mucosal and Regional Immunity. Mucosal Immunol. 2015, 2, 269–306. [Google Scholar] [CrossRef]
- Stokes, C.R.; Bailey, M.; Haverson, K.; Harris, C.; Jones, P.; Inman, C.; Pié, S.; Oswald, I.P.; Williams, B.A.; Akkermans, A.D.L.; et al. Postnatal development of intestinal immune system in piglets: Implications for the process of weaning. Anim. Res. 2004, 53, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Rothkötter, H.; Möllhoff, S.; Pabst, R. The influence of age and breeding conditions on the number and proliferation of intraepithelial lymphocytes in pigs. Scand. J. Immunol. 1999, 50, 31–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, J.E.; Sun, J.; Weber, P.; Navarro, P.; Francis, D. Antibody repertoire development in fetal and newborn piglets, III. Colonization of the gastrointestinal tract selectively diversifies the preimmune repertoire in mucosal lymphoid tissues. Immunology 2000, 100, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Ivaylo, I.I.; Kenya, H. Intestinal Commensal Microbes as Immune Modulators. Cell Host Microbe 2012, 12, 496–508. [Google Scholar] [CrossRef] [Green Version]
- Oliviero, C.; Junnikkala, S.; Peltoniemi, O. The challenge of large litters on the immune system of the sow and the piglets. Reprod. Dom. Anim. 2019, 54, 12–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abreu, M.T. Toll-like receptor signalling in the intestinal epithelium: How bacterial recognition shapes intestinal function. Nat. Rev. Immunol. 2010, 10, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Buettner, M.; Bode, U. Lymph node dissection–understanding the immunological function of lymph nodes. Clin. Exp. Immunol. 2012, 169, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Maroilley, T.; Berri, M.; Lemonnier, G.; Esquerré, D.; Chevaleyre, C.; Mélo, S.; Meurens, F.; Coville, J.L.; Leplat, J.J.; Rau, A.; et al. Immunome differences between porcine ileal and jejunal Peyer’s patches revealed by global transcriptome sequencing of gut-associated lymphoid tissues. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.; Tan, B.; Wang, J.; Liao, S.; Li, J.; Liu, Y.; Yin, Y. Post-natal Growth Retardation Associated With Impaired Gut Hormone Profiles, Immune and Antioxidant Function in Pigs. Front. Endocrinol. 2019, 10, 660. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (Days) | NBW (kg) | IUGR (kg) | p-Value |
|---|---|---|---|
| At birth | 1.58 ± 0.27 | 0.86 ± 0.12 | 0.001 |
| 7 | 2.88 ± 0.52 | 1.64 ± 0.30 | 0.001 |
| 14 | 5.94 ± 1.03 | 3.64 ± 0.91 | 0.001 |
| 180 | 124.0 ± 3.3 | 114.9 ± 5.4 | 0.01 |
| Percentage of Foetal-Type Enterocytes (%) | |||
|---|---|---|---|
| PD 7 | PD 14 | ||
| Duodenum | NBW | Absent | Absent |
| IUGR | 2.12 ± 1.3 | Absent | |
| p-Value | - | - | |
| Proximal jejunum | NBW | Absent | Absent |
| IUGR | 14.02 ± 1.32 | Absent | |
| p-Value | - | - | |
| Middle jejunum | NBW | Very rare | Absent |
| IUGR | 33.4 ± 12.4 a | 1.7 ± 1.53 b | |
| p-Value | 0.01 | - | |
| Distal jejunum | NBW | 34.1 ± 14.6 a | 12.57 ± 3.52 b |
| IUGR | 67.2 ± 30.8 a | 35.1 ± 22.67 b | |
| p-Value | 0.39 | 0.37 | |
| Ileum | NBW | 24.1 ± 8.61 a | 5.54 ± 1.34 b |
| IUGR | 53.55 ± 21.83 a | 18.9 ± 6.81 b | |
| p-Value | 0.01 | 0.001 | |
| Percentage of Intraepithelial Leukocytes (%) | ||||
|---|---|---|---|---|
| PD 7 | PD 14 | PD 180 | ||
| Duodenum | NBW | 15.29 ± 2.45 a | 12.30 ± 2.18 b | 21.3 ± 2.88 c |
| IUGR | 8.87 ± 2.44 a | 13.44 ± 2.42 b | 23.06 ± 5.53 c | |
| p-Value | 0.0001 | NS | NS | |
| Proximal jejunum | NBW | 2.74 ± 0.36 a | 5.26 ± 0.92 b | 39.7 ± 10.12 c |
| IUGR | 4.02 ± 1.32 a | 7.47 ± 0.80 b | 36.19 ± 3.62 c | |
| p-Value | 0.03 | 0.009 | NS | |
| Middle jejunum | NBW | 7.0 ± 1.92 a | 9.36 ± 1.41 b | 20.48 ± 4.00 c |
| IUGR | 13.04 ± 2.17 a | 11.54 ± 1.32 a | 18.36 ± 4.54 b | |
| p-Value | 0.0003 | 0.02 | NS | |
| Distal jejunum | NBW | 11.89 ± 3.70 a | 12.57 ± 3.52 a | 26.6 ± 6.29 b |
| IUGR | 10.74 ± 2.58 a | 11.10 ± 2.67 a | 28.5 ± 2.52 b | |
| p-Value | NS | NS | NS | |
| Ileum | NBW | 6.96 ± 1.49 a | 10.54 ± 1.75 a | 26.2 ± 6.96 b |
| IUGR | 5.55 ± 1.83 a | 9.16 ± 2.19 a | 27.8 ± 9.65 b | |
| p-Value | NS | NS | NS | |
| Percentage of Goblet Cells (%) | ||||
|---|---|---|---|---|
| PD 7 | PD 14 | PD 180 | ||
| Duodenum | NBW | 6.67 ± 1.86 a | 10.81 ± 2.62 b | 7.71 ± 3.46 a,b |
| IUGR | 5.52 ± 1.31 | 6.44 ± 0.68 | 6.74 ± 1.81 | |
| p-Value | NS | 0.0005 | NS | |
| Proximal jejunum | NBW | 7.27 ± 1.69 | 9.10 ± 1.66 | 9.7 ± 3.08 |
| IUGR | 5.85 ± 0.98 | 5.95 ± 1.06 | 6.38 ± 2.57 | |
| p-Value | NS | 0.005 | NS | |
| Middle jejunum | NBW | 5.95 ± 1.39 a | 8.88 ± 1.62 b | 9.66 ± 2.14 b |
| IUGR | 4.93 ± 1.34 a | 6.53 ± 1.3 b | 8.05 ± 3.09 b | |
| p-Value | NS | 0.02 | NS | |
| Distal jejunum | NBW | 9.29 ± 2.62 a | 10.74 ± 2.58 b | 17.0 ± 2.81 c |
| IUGR | 8.74 ± 2.25 a | 8.26 ± 2.17 a | 15.6 ± 5.47 b | |
| p-Value | NS | NS | NS | |
| Ileum | NBW | 22.4 ± 3.49 | 20.54 ± 4.61 | 23.6 ± 6.08 |
| IUGR | 20.15 ± 3.53 | 19.16 ± 3.03 | 18.2 ± 7.10 | |
| p-Value | NS | NS | NS | |
| Distal Jejunum | Ileum | ||||||
|---|---|---|---|---|---|---|---|
| PD 7 | PD 14 | PD 180 | PD 7 | PD 14 | PD 180 | ||
| Average number of cells per Peyer’s patch | NBW | 1450 ± 614 | 1554 ± 454 | N/A | 1203 ± 204 a | 1620 ± 230 b | 6020 ± 2028 c |
| IUGR | 1116 ± 538 | 1003 ± 131 | N/A | 601 ± 173 a | 1340 ± 347 b | 5179 ± 1392 c | |
| p-Value | NS | 0.0261 | 0.0003 | NS | NS | ||
| Average area of a Peyer’s patch (mm2) | NBW | 0.12 ± 0.042 a | 0.266 ± 0.032 b | N/A | 0.238 ± 0.077 a | 0.381 ± 0.03 b | 0.59 ± 0.19 c |
| IUGR | 0.105 ± 0.025 | 0.174 ± 0.08 | N/A | 0.086 ± 0.013 a | 0.34 ± 0.102 b | 0.64 ± 0.13 c | |
| p-Value | 0.001 | 0.0464 | 0.0001 | NS | NS | ||
| Total area of Peyer’s patches per section (mm2) | NBW | 1.67 ± 0.67 | 4.24 ± 3.51 | N/A | 5.21 ± 1.92 a | 9.71 ± 2.44 b | 19.9 ± 10.66 c |
| IUGR | 1.54 ± 1.14 | 3.73 ± 2.15 | N/A | 1.56 ± 0.63 a | 8.13 ± 3.25 b | 20.0 ± 4.52 c | |
| p-Value | NS | NS | 0.0006 | NS | NS | ||
| Average density of cells per Peyer’s patch (number of cells/mm2) | NBW | 13,302 ± 2122 | 7240 ± 2742 | N/A | 5578 ± 1241 a | 5006 ± 1697 a | 10362 ± 2498 b |
| IUGR | 12,177 ± 4582 | 9379 ± 938 | N/A | 7263 ± 1205 a | 4628 ± 1443 b | 8245 ± 2258 a | |
| p-Value | NS | NS | 0.02 | NS | NS | ||
| PD 0–1 | PD 7–8 | PD 14 | PD 35 * | PD 42 | PD 180 | |
|---|---|---|---|---|---|---|
| Foetal-type enterocytes | ND | Duo ↑ Prox ↑↑ Mid ↑↑ Ile ↑↑ | Mid ↑ Ile ↑↑↑ | N/A | ND | ND |
| Intraepithelial leukocytes | Ile↓ | Duo ↑↑↑ Pox ↑ Mid ↑↑↑ | Prox ↑↑ Mid ↑ | N/A | N/A | ND |
| Goblet cells | Mid ↓ Ile ↓ | ND | Duo ↓↓ Prox ↓↓ Mid ↓ | ILE↓ | N/A | ND |
| Peyer’s patches | ↓ | ↓ | ND | N/A | N/A | ND |
| Inflammatory cytokines | TNF-α ↓↓ INF-γ ↓↓ IL-10 ↓↓ | N/A | TNFα ↑↑ | TNF-α ↑↑ INF-γ ↑↑ IL1 ↑↑ IL-6 ↑ | INF-γ ↓ IL-1β ↓ IL-6 ↓ | N/A |
| References | [4,9,13] | [13] | [13] | [18] | [33] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olszewski, J.; Zabielski, R.; Skrzypek, T.; Matyba, P.; Wierzbicka, M.; Adamski, A.; Grzesiuk, E.; Sady, M.; Gajewski, Z.; Ferenc, K. Differences in Intestinal Barrier Development between Intrauterine Growth Restricted and Normal Birth Weight Piglets. Animals 2021, 11, 990. https://doi.org/10.3390/ani11040990
Olszewski J, Zabielski R, Skrzypek T, Matyba P, Wierzbicka M, Adamski A, Grzesiuk E, Sady M, Gajewski Z, Ferenc K. Differences in Intestinal Barrier Development between Intrauterine Growth Restricted and Normal Birth Weight Piglets. Animals. 2021; 11(4):990. https://doi.org/10.3390/ani11040990
Chicago/Turabian StyleOlszewski, Jarosław, Romuald Zabielski, Tomasz Skrzypek, Piotr Matyba, Małgorzata Wierzbicka, Antoni Adamski, Elżbieta Grzesiuk, Maria Sady, Zdzisław Gajewski, and Karolina Ferenc. 2021. "Differences in Intestinal Barrier Development between Intrauterine Growth Restricted and Normal Birth Weight Piglets" Animals 11, no. 4: 990. https://doi.org/10.3390/ani11040990