
Effects of Protein Restriction and Subsequent Realimentation on Body Composition, Gut Microbiota and Metabolite Profiles in Weaned Piglets

 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Sample Collection
2.3. Chemical Analyses
2.4. Serum Biochemical Parameters
2.5. DNA Extraction and 16S rRNA Amplification
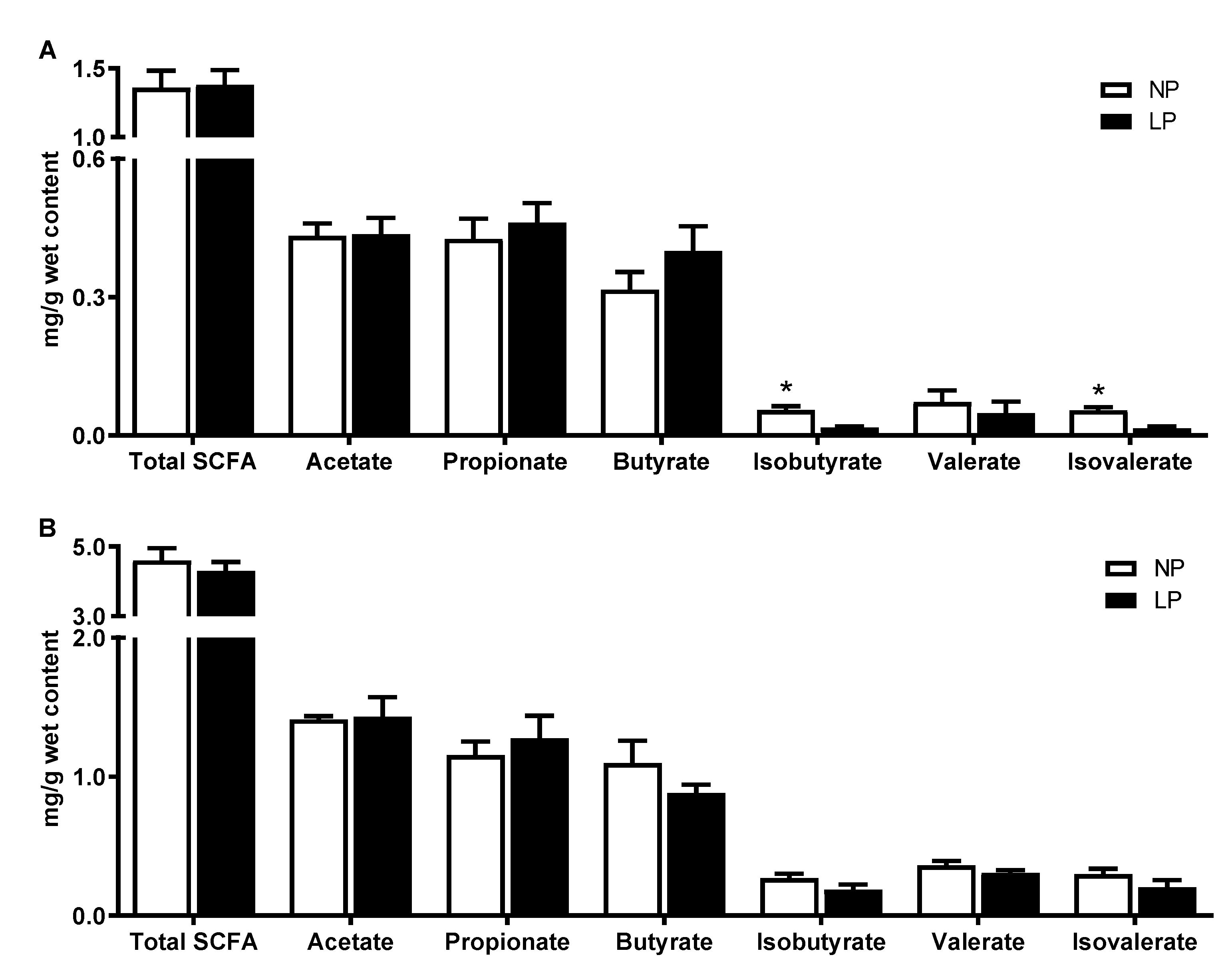
2.6. Microbial Metabolite Analysis
2.7. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Body Composition
3.3. Serum Biochemical Parameters
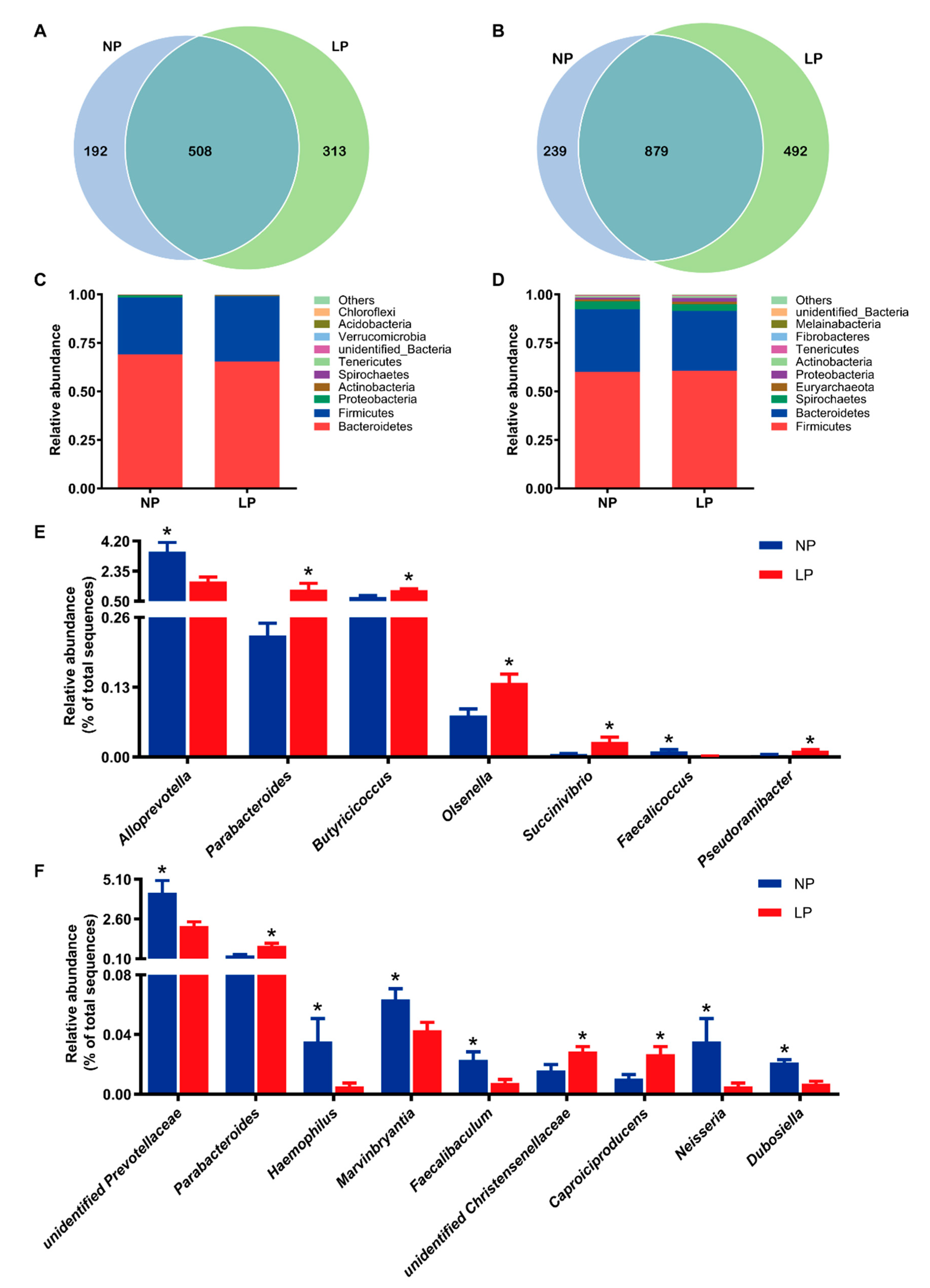
3.4. Colonic Bacterial Community Structure
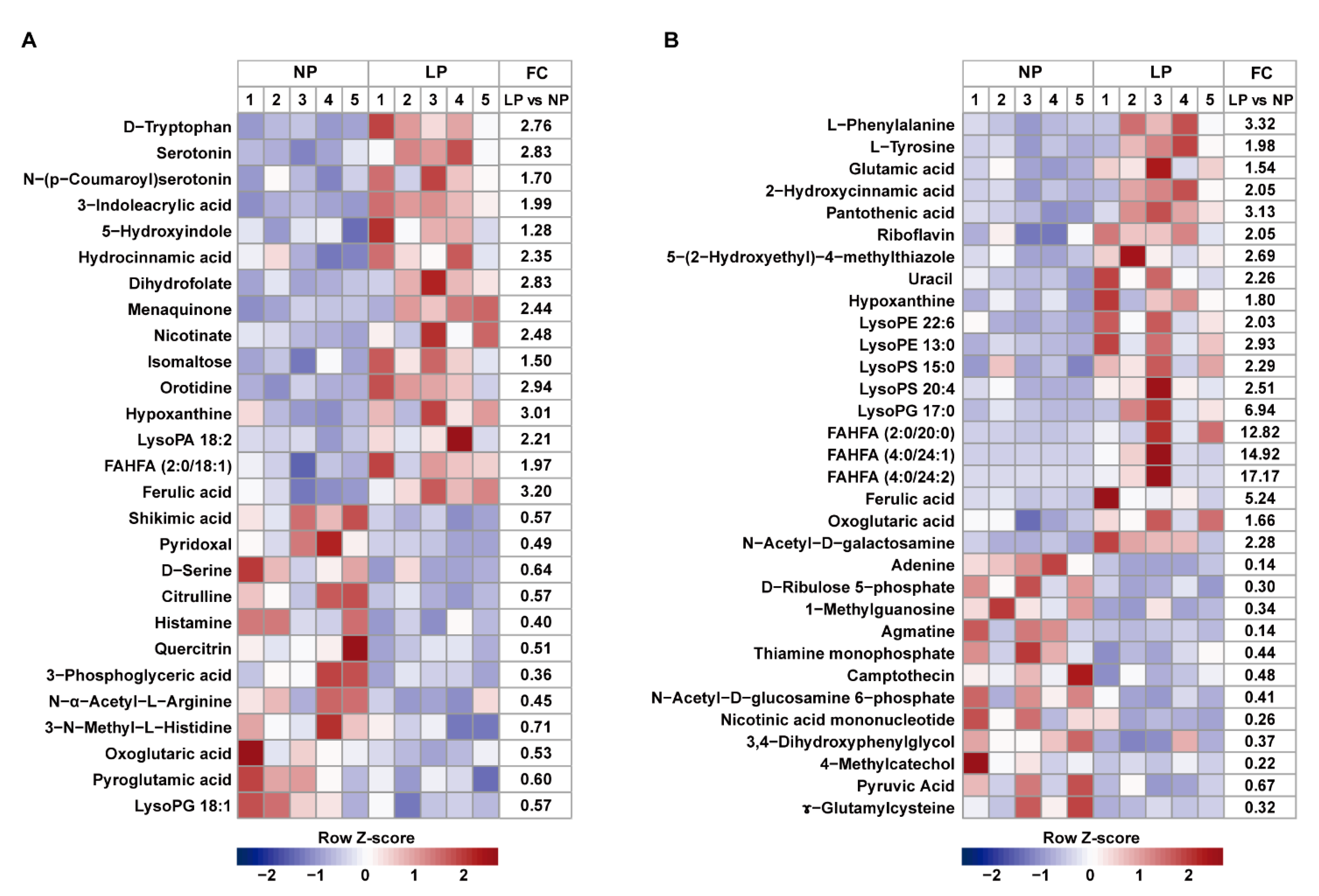
3.5. Metabolite Profiles in the Colonic Digesta
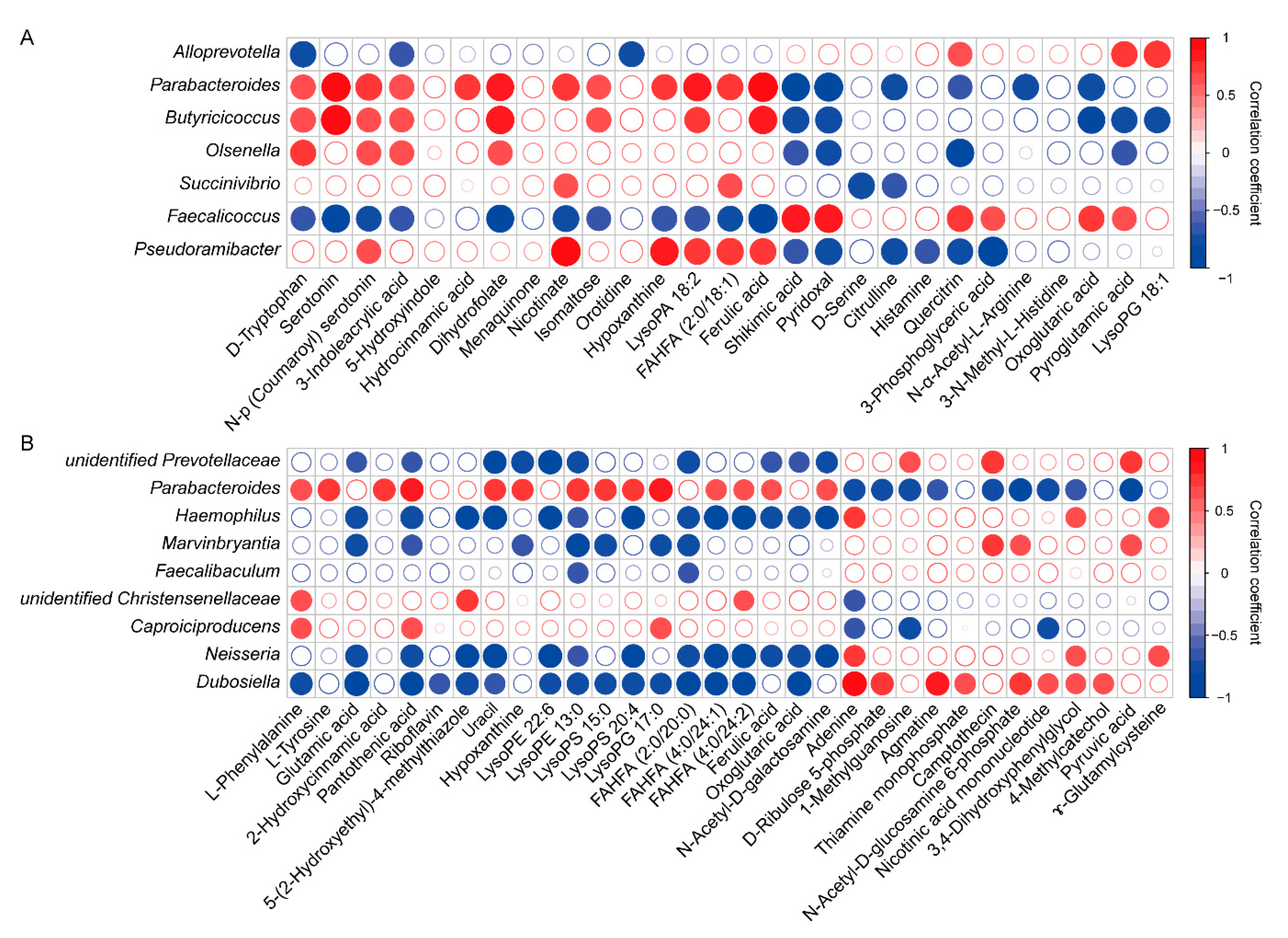
3.6. Correlation between Microbiota Community and Metabolites
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pluske, J.R.; Hampson, D.J.; Williams, I.H. Factors influencing the structure and function of the small intestine in the weaned pig: A review. Livest. Prod. Sci. 1997, 51, 215–236. [Google Scholar] [CrossRef]
- Wellock, I.J.; Fortomaris, P.D.; Houdijk, J.G.M.; Kyriazakis, I. The effect of dietary protein supply on the performance and risk of post-weaning enteric disorders in newly weaned pigs. Anim. Sci. 2006, 82, 327–335. [Google Scholar] [CrossRef]
- Wellock, I.J.; Fortomaris, P.D.; Houdijk, J.G.M.; Kyriazakis, I. Effects of dietary protein supply, weaning age and experimental enterotoxigenic Escherichia coli infection on newly weaned pigs: Health. Animal 2008, 2, 834–842. [Google Scholar] [CrossRef]
- Nyachoti, C.M.; Omogbenigun, F.O.; Rademacher, M.; Blank, G. Performance responses and indicators of gastrointestinal health in early-weaned pigs fed low-protein amino acid-supplemented diets. J. Anim. Sci. 2006, 84, 125–134. [Google Scholar] [CrossRef]
- Yu, D.; Zhu, W.; Hang, S. Effects of Long-Term Dietary Protein Restriction on Intestinal Morphology, Digestive Enzymes, Gut Hormones, and Colonic Microbiota in Pigs. Animals 2019, 9, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Q.; Zhu, Y.; Wang, J.; Yang, H.; Wang, J.; Zhu, W. Protein restriction and succedent realimentation affecting ileal morphology, ileal microbial composition and metabolites in weaned piglets. Animal 2019, 13, 2463–2472. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Fu, H.Y.; Qiu, S.N.; Teng, T.; Bai, G.D.; Ju, D.X.; Sun, Y.C.; Shi, B.M. Effects of early protein restriction on the growth performance and gut development of pigs fed diets with or without antibiotic. Animal 2020, 14, 1392–1401. [Google Scholar] [CrossRef] [PubMed]
- Pluske, J.R.; Dividich, J.L.; Verstegen, M.W.A. Weaning the Pig: Concepts and Consequences; Wageningen Academic Publishers: Wageningen, The Netherlands, 2003. [Google Scholar]
- Hinson, R.; Schinckel, A.; Radcliffe, J.; Allee, G.; Sutton, A.; Richert, B. Effect of feeding reduced crude protein and phosphorus diets on weaning-finishing pig growth performance, carcass characteristics, and bone characteristics. J. Anim. Sci. 2008, 87, 1502–1517. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.W.; O’Quinn, P.R.; Goodband, R.D.; Tokach, M.D.; Nelssen, J.L. Effects of Low-Protein, Amino Acid-Fortified Diets Formulated on a Net Energy Basis on Growth Performance and Carcass Characteristics of Finishing Pigs. J. Appl. Anim. Res. 1999, 15, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Morazán, H.; Alvarez-Rodriguez, J.; Seradj, A.R.; Balcells, J.; Babot, D. Trade-offs among growth performance, nutrient digestion and carcass traits when feeding low protein and/or high neutral-detergent fiber diets to growing-finishing pigs. Anim. Feed Sci. Technol. 2015, 207, 168–180. [Google Scholar] [CrossRef]
- Qiu, Y.; Liu, S.; Hou, L.; Li, K.; Wang, L.; Gao, K.; Yang, X.; Jiang, Z. Supplemental Choline Modulates Growth Performance and Gut Inflammation by Altering the Gut Microbiota and Lipid Metabolism in Weaned Piglets. J. Nutr. 2021, 151, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Li, Y.; Han, H.; Liu, Z.J.; Zeng, X.; Li, T.; Yin, Y. Long-Term Effects of Lysine Concentration on Growth Performance, Intestinal Microbiome, and Metabolic Profiles in a Pig model. Food Funct. 2018, 9, 4153–4163. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Li, Z.; Chen, W.; Rong, T.; Wang, G.; Ma, X. Microbiome-Metabolomics Analysis Investigating the Impacts of Dietary Starch Types on the Composition and Metabolism of Colonic Microbiota in Finishing Pigs. Front. Microbiol. 2019, 10, 1143. [Google Scholar] [CrossRef] [Green Version]
- NRC. Nutrient Requirements of Swine, 11th ed.; National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Jones, C.K.; Gabler, N.K.; Main, R.G.; Patience, J.F. Characterizing growth and carcass composition differences in pigs with varying weaning weights and postweaning performance. J. Anim. Sci. 2012, 90, 4072–4080. [Google Scholar] [CrossRef] [Green Version]
- AOAC. Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2000. [Google Scholar]
- Xiong, Y.; Yi, H.; Wu, Q.; Jiang, Z.; Wang, L. Effects of acute heat stress on intestinal microbiota in grow-finishing pigs, and associations with feed intake and serum profile. J. Appl. Microbiol. 2020, 128, 840–852. [Google Scholar] [CrossRef]
- Sun, Z.; Li, J.; Dai, Y.; Wang, W.; Shi, R.; Wang, Z.; Ding, P.; Lu, Q.; Jiang, H.; Pei, W.; et al. Indigo Naturalis Alleviates Dextran Sulfate Sodium-Induced Colitis in Rats via Altering Gut Microbiota. Front. Microbiol. 2020, 11, 731. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.M.; Kim, J.C.; Hansen, C.F.; Mullan, B.P.; Hampson, D.J.; Pluske, J.R. Feeding a diet with decreased protein content reduces indices of protein fermentation and the incidence of postweaning diarrhea in weaned pigs challenged with an enterotoxigenic strain of Escherichia coli. J. Anim. Sci. 2009, 87, 2833–2843. [Google Scholar] [CrossRef] [Green Version]
- Kil, D.Y.; Stein, H.H. Invited Review: Management and feeding strategies to ameliorate the impact of removing antibiotic growth promoters from diets fed to weanling pigs. Can. J. Anim. Sci. 2010, 90, 447–460. [Google Scholar] [CrossRef]
- Chaosap, C.; Parr, T.; Wiseman, J. Effect of compensatory growth on performance, carcass composition and plasma IGF-1 in grower finisher pigs. Animal 2011, 5, 749–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebret, B.; Heyer, A.; Gondret, F.; Louveau, I. The response of various muscle types to a restriction -re-alimentation feeding strategy in growing pigs. Animal 2007, 1, 849–857. [Google Scholar] [CrossRef] [Green Version]
- Conde-Aguilera, J.; Aguinaga, M.A.; Lara, L.; Aguilera, J.; Nieto, R. Carcass traits and organ weights of 10–25-kg body weight Iberian pigs fed diets with different protein-to-energy ratio. Anim. Feed Sci. Technol. 2011, 164, 116–124. [Google Scholar] [CrossRef]
- Ruiz-Ascacibar, I.; Stoll, P.; Kreuzer, M.; Boillat, V.; Spring, P.; Bee, G. Impact of amino acid and CP restriction from 20 to 140 kg BW on performance and dynamics in empty body protein and lipid deposition of entire male, castrated and female pigs. Animal 2016, 11, 394–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, D.; Yao, K.; Chu, W.; Li, T.; Huang, R.; Yin, Y.; Liu, Z.; Zhang, J.; Wu, G. Impaired translation initiation activation and reduced protein synthesis in weaned piglets fed a low-protein diet. J. Nutr. Biochem. 2009, 20, 544–552. [Google Scholar] [CrossRef]
- Drouillard, J.; Britton, R.; Bauer, M.; Gramlich, S.; Wester, T.; Ferrell, C. Growth, body composition, and visceral organ mass and metabolism in lambs during and after metabolizable protein or net energy restrictions. J. Anim. Sci. 1991, 69, 3357–3375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bikker, P.; Verstegen, M.; Kemp, B.; Bosch, M. Performance and Body Composition of Finishing Gilts (45 to 85 Kilograms) as Affected by Energy Intake and Nutrition in Earlier Life: I. Growth of the Body and Body Components. J. Anim. Sci. 1996, 74, 806–816. [Google Scholar] [CrossRef] [Green Version]
- de Meer, K.; Smolders, H.C.; Meesterburrie, J.; de Sain-van der Velden, M.; Voorbij, H.A.; Okken, A.; Reijngoud, D.J.; Kulik, W. A single food bolus stimulates albumin synthesis in growing piglets. J. Pediatr. Gastroenterol. Nutr. 2000, 31, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, C.; Boisen, S. Studies on the Response Time for Plasma Urea Nitrogen as a Rapid Measure for Dietary Protein Quality in Pigs. Acta Agric. Scand. A Anim. 2001, 51, 209–216. [Google Scholar] [CrossRef]
- Wang, J.H.; Wu, C.C.; Feng, J. Effect of dietary antibacterial peptide and zinc-methionine on performance and serum biochemical parameters in piglets. Czech J. Anim. Sci. 2011, 56, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Millet, S.; Aluwe, M.; De Boever, J.; De Witte, B.; Douidah, L.; Van den Broeke, A.; Leen, F.; De Cuyper, C.; Ampe, B.; De Campeneere, S. The effect of crude protein reduction on performance and nitrogen metabolism in piglets (four to nine weeks of age) fed two dietary lysine levels. J. Anim. Sci. 2018, 96, 3824–3836. [Google Scholar] [CrossRef]
- Wu, Y.; Jiang, Z.; Zheng, C.; Wang, L.; Zhu, C.; Yang, X.; Wen, X.; Ma, X. Effects of protein sources and levels in antibiotic-free diets on diarrhea, intestinal morphology, and expression of tight junctions in weaned piglets. Anim. Nutr. 2015, 1, 170–176. [Google Scholar] [CrossRef]
- Fang, L.H.; Jin, Y.H.; Do, S.H.; Hong, J.S.; Kim, B.O.; Han, T.H.; Kim, Y.Y. Effects of dietary energy and crude protein levels on growth performance, blood profiles, and nutrient digestibility in weaning pigs. Asian-Australas. J. Anim. Sci. 2019, 32, 556–563. [Google Scholar]
- Ijaz, M.U.; Ahmed, M.I.; Zou, X.; Hussain, M.; Zhang, M.; Zhao, F.; Xu, X.; Zhou, G.; Li, C. Beef, Casein, and Soy Proteins Differentially Affect Lipid Metabolism, Triglycerides Accumulation and Gut Microbiota of High-Fat Diet-Fed C57BL/6J Mice. Front. Microbiol. 2018, 9, 2200. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tsai, T.; Deng, F.; Wei, X.; Chai, J.; Knapp, J.; Apple, J.; Maxwell, C.V.; Lee, J.A.; Li, Y.; et al. Longitudinal investigation of the swine gut microbiome from birth to market reveals stage and growth performance associated bacteria. Microbiome 2019, 7, 109. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, R.; Middelkoop, A.; Bolhuis, J.E.; Kleerebezem, M. Legitimate and Reliable Determination of the Age-Related Intestinal Microbiome in Young Piglets; Rectal Swabs and Fecal Samples Provide Comparable Insights. Front. Microbiol. 2019, 10, 1886. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.; Tiezzi, F.; Schillebeeckx, C.; McNulty, N.P.; Schwab, C.; Shull, C.; Maltecca, C. Host contributes to longitudinal diversity of fecal microbiota in swine selected for lean growth. Microbiome 2018, 6, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.X.; Pan, Q.; Ma, F.; Li, P.J.; Xu, B.C.; Chen, C.G. Amelioration of Growth Performance, Lipid Accumulation, and Intestinal Health in Mice by a Cooked Mixture of Lean Meat and Resistant Starch. Mol. Nutr. Food Res. 2019, 63, e1801364. [Google Scholar] [CrossRef]
- Hou, Q.; Kwok, L.Y.; Zheng, Y.; Wang, L.; Guo, Z.; Zhang, J.; Huang, W.; Wang, Y.; Leng, L.; Li, H.; et al. Differential fecal microbiota are retained in broiler chicken lines divergently selected for fatness traits. Sci. Rep. 2016, 6, 37376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, C.; Gao, R.; Yan, X.; Huang, L.; Qin, H. Probiotics improve gut microbiota dysbiosis in obese mice fed a high-fat or high-sucrose diet. Nutrition 2019, 60, 175–184. [Google Scholar] [CrossRef]
- Kong, F.; Gao, Y.; Tang, M.; Fu, T.; Diao, Q.; Bi, Y.; Tu, Y. Effects of dietary rumen-protected Lys levels on rumen fermentation and bacterial community composition in Holstein heifers. Appl. Microbiol. Biotechnol. 2020, 104, 6623–6634. [Google Scholar] [CrossRef] [PubMed]
- Filippo, C.; Di Paola, M.; Ramazzotti, M.; Albanese, D.; Pieraccini, G.; Banci, E.; Miglietta, F.; Cavalieri, D.; Lionetti, P. Diet, Environments, and Gut Microbiota. A Preliminary Investigation in Children Living in Rural and Urban Burkina Faso and Italy. Front. Microbiol. 2017, 8, 1979. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Xiao, X.; Zhang, Q.; Zheng, J.; Li, M.; Wang, X.; Deng, M.; Zhai, X.; Liu, J. Gut microbiota might be a crucial factor in deciphering the metabolic benefits of perinatal genistein consumption in dams and adult female offspring. Food Funct. 2019, 10, 4505–4521. [Google Scholar] [CrossRef] [Green Version]
- Louis, S.; Tappu, R.M.; Damms-Machado, A.; Huson, D.H.; Bischoff, S.C. Characterization of the Gut Microbial Community of Obese Patients Following a Weight-Loss Intervention Using Whole Metagenome Shotgun Sequencing. PLoS ONE 2016, 11, e0149564. [Google Scholar] [CrossRef]
- Kang, Y.; Li, Y.; Du, Y.; Guo, L.; Chen, M.; Huang, X.; Yang, F.; Hong, J.; Kong, X. Konjaku flour reduces obesity in mice by modulating the composition of the gut microbiota. Int. J. Obes. 2019, 43, 1631–1643. [Google Scholar] [CrossRef]
- Forbes, J.D.; Chen, C.Y.; Knox, N.C.; Marrie, R.A.; El-Gabalawy, H.; de Kievit, T.; Alfa, M.; Bernstein, C.N.; Van Domselaar, G. A comparative study of the gut microbiota in immune-mediated inflammatory diseases-does a common dysbiosis exist? Microbiome 2018, 6, 221. [Google Scholar] [CrossRef]
- Waters, J.L.; Ley, R.E. The human gut bacteria Christensenellaceae are widespread, heritable, and associated with health. BMC Biol. 2019, 17, 83. [Google Scholar] [CrossRef]
- Zhang, W.J.; Su, W.W.; Li, P.B.; Rao, H.Y.; Lin, Q.W.; Zeng, X.; Chen, T.B.; Yan, Z.H.; Liu, H.; Yao, H.L. Naoxintong Capsule Inhibits the Development of Cardiovascular Pathological Changes in Bama Minipig Through Improving Gut Microbiota. Front. Pharmacol. 2019, 10, 1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Tian, G.; Chen, D.; Zheng, P.; Yu, J.; He, J.; Mao, X.; Huang, Z.; Luo, Y.; Luo, J.; et al. Dietary protein levels and amino acid supplementation patterns alter the composition and functions of colonic microbiota in pigs. Anim. Nutr. 2020, 6, 143–151. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, Q.; Sun, S.; Huang, B.; Zhang, Y.; Xu, Y.; Zhang, S.; Xiang, H. Probiotics-fermented Massa Medicata Fermentata ameliorates weaning stress in piglets related to improving intestinal homeostasis. Appl. Microbiol. Biotechnol. 2018, 102, 10713–10727. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hu, C.; Cheng, C.; Cui, J.; Ji, Y.; Hao, X.; Li, Q.; Ren, W.; Deng, B.; Yin, Y.; et al. Unraveling the association of fecal microbiota and oxidative stress with stillbirth rate of sows. Theriogenology 2019, 136, 131–137. [Google Scholar] [CrossRef]
- Shahi, S.K.; Freedman, S.N.; Mangalam, A.K. Gut microbiome in multiple sclerosis: The players involved and the roles they play. Gut Microbes 2017, 8, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Rajilić-Stojanović, M.; Biagi, E.; Heilig, H.G.; Kajander, K.; Kekkonen, R.A.; Tims, S.; de Vos, W.M. Global and deep molecular analysis of microbiota signatures in fecal samples from patients with irritable bowel syndrome. Gastroenterology 2011, 141, 1792–1801. [Google Scholar] [CrossRef]
- Marri, P.R.; Paniscus, M.; Weyand, N.J.; Rendón, M.A.; Calton, C.M.; Hernández, D.R.; Higashi, D.L.; Sodergren, E.; Weinstock, G.M.; Rounsley, S.D.; et al. Genome sequencing reveals widespread virulence gene exchange among human Neisseria species. PLoS ONE 2010, 5, e11835. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Macchietto, M.G.; Liu, X.; Lu, Y.; Ma, Y.; Guo, H.; Saqui-Salces, M.; Bernlohr, D.A.; Chen, C.; Shen, S.; et al. Identification of gut microbiota and microbial metabolites regulated by an antimicrobial peptide lipocalin 2 in high fat diet-induced obesity. Int. J. Obes. 2021, 45, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Rist, V.T.; Weiss, E.; Eklund, M.; Mosenthin, R. Impact of dietary protein on microbiota composition and activity in the gastrointestinal tract of piglets in relation to gut health: A review. Animal 2013, 7, 1067–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Z.L.; Wu, G.; Zhu, W.Y. Amino acid metabolism in intestinal bacteria: Links between gut ecology and host health. Front. Biosci. 2011, 16, 1768–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef] [Green Version]
- Kepert, I.; Fonseca, J.; Müller, C.; Milger, K.; Hochwind, K.; Kostric, M.; Fedoseeva, M.; Ohnmacht, C.; Dehmel, S.; Nathan, P.; et al. D-tryptophan from probiotic bacteria influences the gut microbiome and allergic airway disease. J. Allergy Clin. Immunol. 2017, 139, 1525–1535. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Wang, Z.; Qin, Z.; Cao, J.; Chen, Y. Role of serotonin in the intestinal mucosal epithelium barrier in weaning mice undergoing stress-induced diarrhea. J. Mol. Histol. 2018, 49, 85–97. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, S.; Ma, N.; Johnston, L.J.; Wu, C.; Ma, X. Metabolites of microbiota response to tryptophan and intestinal mucosal immunity: A therapeutic target to control intestinal inflammation. Med. Res. Rev. 2020. [Google Scholar] [CrossRef] [PubMed]
- Bosch, S.; El Manouni El Hassani, S.; Brizzio Brentar, M.; Ayada, I.; Bakkali, A.; Jansen, E.E.W.; Struys, E.A.; Benninga, M.A.; de Boer, N.K.H.; de Meij, T.G.J. Fecal Amino Acid Profiles Exceed Accuracy of Serum Amino Acids in Diagnosing Pediatric Inflammatory Bowel Disease. J. Pediatr. Gastroenterol. Nutr. 2020, 71, 371–375. [Google Scholar] [CrossRef]
- Grosheva, I.; Zheng, D.; Levy, M.; Polansky, O.; Lichtenstein, A.; Golani, O.; Dori-Bachash, M.; Moresi, C.; Shapiro, H.; Del Mare-Roumani, S.; et al. High-Throughput Screen Identifies Host and Microbiota Regulators of Intestinal Barrier Function. Gastroenterology 2020, 159, 1807–1823. [Google Scholar] [CrossRef]
- Collins, S.M.; Surette, M.; Bercik, P. The interplay between the intestinal microbiota and the brain. Nat. Rev. Microbiol. 2012, 10, 735–742. [Google Scholar] [CrossRef]
- Li, Y.; Guo, Y.; Wen, Z.; Jiang, X.; Ma, X.; Han, X. Weaning Stress Perturbs Gut Microbiome and Its Metabolic Profile in Piglets. Sci. Rep. 2018, 8, 18068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turroni, S.; Fiori, J.; Rampelli, S.; Schnorr, S.L.; Consolandi, C.; Barone, M.; Biagi, E.; Fanelli, F.; Mezzullo, M.; Crittenden, A.N.; et al. Fecal metabolome of the Hadza hunter-gatherers: A host-microbiome integrative view. Sci. Rep. 2016, 6, 32826. [Google Scholar] [CrossRef]
- Turroni, S.; Brigidi, P.; Cavalli, A.; Candela, M. Microbiota-Host Transgenomic Metabolism, Bioactive Molecules from the Inside. J. Med. Chem. 2018, 61, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Ge, W.; Wang, J.; Liu, W.; Zhao, Y.; Shen, W.; Li, L. Zearalenone-induced aberration in the composition of the gut microbiome and function impacts the ovary reserve. Chemosphere 2020, 244, 125493. [Google Scholar] [CrossRef]
- Yore, M.M.; Syed, I.; Moraes-Vieira, P.M.; Zhang, T.; Herman, M.A.; Homan, E.A.; Patel, R.T.; Lee, J.; Chen, S.; Peroni, O.D.; et al. Discovery of a class of endogenous mammalian lipids with anti-diabetic and anti-inflammatory effects. Cell 2014, 159, 318–332. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Moraes-Vieira, P.M.; Castoldi, A.; Aryal, P.; Yee, E.U.; Vickers, C.; Parnas, O.; Donaldson, C.J.; Saghatelian, A.; Kahn, B.B. Branched Fatty Acid Esters of Hydroxy Fatty Acids (FAHFAs) Protect against Colitis by Regulating Gut Innate and Adaptive Immune Responses. J. Biol. Chem. 2016, 291, 22207–22217. [Google Scholar] [CrossRef] [Green Version]
- Syed, I.; Rubin de Celis, M.F.; Mohan, J.F.; Moraes-Vieira, P.M.; Vijayakumar, A.; Nelson, A.T.; Siegel, D.; Saghatelian, A.; Mathis, D.; Kahn, B.B. PAHSAs attenuate immune responses and promote β cell survival in autoimmune diabetic mice. J. Clin. Investig. 2019, 129, 3717–3731. [Google Scholar] [CrossRef] [Green Version]
- Brejchova, K.; Balas, L.; Paluchova, V.; Brezinova, M.; Durand, T.; Kuda, O. Understanding FAHFAs: From structure to metabolic regulation. Prog. Lipid Res. 2020, 79, 101053. [Google Scholar] [CrossRef] [PubMed]
- Kuda, O.; Brezinova, M.; Rombaldova, M.; Slavikova, B.; Posta, M.; Beier, P.; Janovska, P.; Veleba, J.; Kopecky, J., Jr.; Kudova, E.; et al. Docosahexaenoic Acid-Derived Fatty Acid Esters of Hydroxy Fatty Acids (FAHFAs) With Anti-inflammatory Properties. Diabetes 2016, 65, 2580–2590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paluchova, V.; Vik, A.; Cajka, T.; Brezinova, M.; Brejchova, K.; Bugajev, V.; Draberova, L.; Draber, P.; Buresova, J.; Kroupova, P.; et al. Triacylglycerol-Rich Oils of Marine Origin are Optimal Nutrients for Induction of Polyunsaturated Docosahexaenoic Acid Ester of Hydroxy Linoleic Acid (13-DHAHLA) with Anti-Inflammatory Properties in Mice. Mol. Nutr. Food Res. 2020, 64, e1901238. [Google Scholar] [CrossRef] [PubMed]
- Gowda, S.G.B.; Liang, C.; Gowda, D.; Hou, F.; Kawakami, K.; Fukiya, S.; Yokota, A.; Chiba, H.; Hui, S.P. Identification of short-chain fatty acid esters of hydroxy fatty acids (SFAHFAs) in a murine model by nontargeted analysis using ultra-high-performance liquid chromatography/linear ion trap quadrupole-Orbitrap mass spectrometry. Rapid Commun. Mass Spectrom. 2020, 34, e8831. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; Milani, C.; de Giori, G.S.; Sesma, F.; van Sinderen, D.; Ventura, M. Bacteria as vitamin suppliers to their host: A gut microbiota perspective. Curr. Opin. Biotechnol. 2013, 24, 160–168. [Google Scholar] [CrossRef]
- Digby, J.E.; Martinez, F.; Jefferson, A.; Ruparelia, N.; Chai, J.; Wamil, M.; Greaves, D.R.; Choudhury, R.P. Anti-inflammatory effects of nicotinic acid in human monocytes are mediated by GPR109A dependent mechanisms. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 669–676. [Google Scholar] [CrossRef] [Green Version]
- Kunisawa, J.; Hashimoto, E.; Ishikawa, I.; Kiyono, H. A pivotal role of vitamin B9 in the maintenance of regulatory T cells in vitro and in vivo. PLoS ONE 2012, 7, e32094. [Google Scholar] [CrossRef] [Green Version]
- Brestoff, J.R.; Artis, D. Commensal bacteria at the interface of host metabolism and the immune system. Nat. Immunol. 2013, 14, 676–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Restriction Phase d 0 to 14 | Realimentation Phase d 15 to 25 kg of BW | |
|---|---|---|---|
| NP-20% | LP-16% | NP-19% | |
| Ingredients, % | |||
| Corn | 34.94 | 45.08 | 68.43 |
| Expanded corn | 10.00 | 10.00 | |
| Expanded soybean | 8.00 | 4.00 | |
| Soybean meal, enzyme treated | 10.00 | 7.92 | 10.00 |
| Soybean meal | 3.20 | 14.46 | |
| Fish meal | 3.20 | 2.00 | 2.00 |
| Whey protein concentrate | 3.20 | 2.00 | |
| Whey powder | 15.00 | 15.00 | |
| Yeast extract | 2.00 | 2.00 | |
| Soybean hulls | 2.00 | 2.00 | |
| Soybean oil | 1.00 | 1.00 | 1.00 |
| Sucrose | 2.50 | 2.50 | |
| Salt | 0.20 | 0.20 | 0.35 |
| Dicalcium phosphate | 1.05 | 1.47 | 0.55 |
| Limestone | 0.93 | ||
| Calcium Citrate | 0.90 | 0.88 | |
| L-Lys HCl | 0.59 | 0.99 | 0.47 |
| DL-Met | 0.10 | 0.18 | 0.06 |
| L-Thr | 0.19 | 0.37 | 0.12 |
| L-Trp | 0.02 | 0.09 | 0.01 |
| L-Val | 0.11 | 0.32 | |
| L-Ile | 0.20 | ||
| Phytase | 0.02 | 0.02 | 0.02 |
| Zinc oxide | 0.18 | 0.18 | |
| Choline chloride | 0.20 | 0.20 | 0.20 |
| Titanium dioxide | 0.40 | 0.40 | 0.40 |
| Premix 1 | 1.00 | 1.00 | 1.00 |
| Total | 100.00 | 100.00 | 100.00 |
| Calculated nutrient composition 2, % | |||
| NE 3, MJ/kg | 10.82 | 10.92 | 10.50 |
| SID 4 Lys | 1.41 | 1.41 | 1.23 |
| SID 4 Met | 0.41 | 0.41 | 0.36 |
| SID 4 Thr | 0.83 | 0.83 | 0.73 |
| SID 4 Trp | 0.23 | 0.23 | 0.20 |
| SID 4 Val | 0.89 | 0.89 | 0.78 |
| SID 4 Ile | 0.73 | 0.72 | 0.71 |
| Calcium | 0.82 | 0.82 | 0.75 |
| Total phosphorus | 0.72 | 0.72 | 0.61 |
| Analyzed nutrient composition, % | |||
| CP 5 | 20.09 | 16.33 | 19.35 |
| Item | NP | LP | SEM | p-Value |
|---|---|---|---|---|
| Restriction phase (day 0 to 14) | ||||
| Initial BW, kg | 6.39 | 6.38 | 0.02 | 0.861 |
| ADG, g | 324 | 261 | 18.1 | 0.041 |
| ADFI, g | 418 | 372 | 17.0 | 0.093 |
| G:F | 0.77 | 0.70 | 0.02 | 0.022 |
| Diarrhea incidence, % | 2.00 | 0.29 | 0.55 | 0.060 |
| BW at end of phase, kg | 10.9 | 10.0 | 0.26 | 0.043 |
| Realimentation phase (day 15 to target BW) | ||||
| ADG, g | 524 | 543 | 7.9 | 0.153 |
| ADFI, g | 859 | 888 | 10.1 | 0.086 |
| G:F | 0.61 | 0.61 | 0.01 | 1.000 |
| Diarrhea incidence, % | 5.33 | 2.61 | 1.21 | 0.151 |
| Final BW, kg | 24.9 | 25.1 | 0.23 | 0.579 |
| Overall (day 0 to target BW) | ||||
| ADG, g | 455 | 452 | 9.5 | 0.814 |
| ADFI, g | 708 | 718 | 8.2 | 0.413 |
| G:F | 0.64 | 0.63 | 0.01 | 0.283 |
| Diarrhea incidence, % | 3.76 | 1.56 | 0.82 | 0.095 |
| Days on experiment | 40.8 | 41.5 | 0.81 | 0.537 |
| Item | NP | LP | SEM | p-Value |
|---|---|---|---|---|
| Day 14 of the experiment | ||||
| Water, g/kg | 709 | 708 | 4.3 | 0.845 |
| Protein, g/kg | 163 | 157 | 1.8 | 0.034 |
| Lipid, g/kg | 90.1 | 97.3 | 4.24 | 0.261 |
| Ash, g/kg | 31.0 | 31.4 | 1.40 | 0.846 |
| End of the experiment | ||||
| Water, g/kg | 694 | 689 | 4.6 | 0.427 |
| Protein, g/kg | 168 | 166 | 1.6 | 0.451 |
| Lipid, g/kg | 107 | 113 | 6.5 | 0.523 |
| Ash, g/kg | 28.2 | 27.1 | 1.46 | 0.622 |
| Item | NP | LP | SEM | p-Value |
|---|---|---|---|---|
| Day 14 of the experiment | ||||
| Total protein, g/L | 51.2 | 46.1 | 1.79 | 0.085 |
| Albumin, g/L | 29.6 | 23.8 | 1.31 | 0.017 |
| Urea, mmol/L | 2.82 | 1.66 | 0.25 | 0.018 |
| Glucose, mmol/L | 7.57 | 6.48 | 0.55 | 0.203 |
| Triglyceride, mmol/L | 0.49 | 0.50 | 0.05 | 0.845 |
| Creatinine, μmol/L | 71.3 | 61.8 | 5.30 | 0.245 |
| End of the experiment | ||||
| Total protein, g/L | 56.8 | 54.8 | 1.48 | 0.357 |
| Albumin, g/L | 31.8 | 29.0 | 1.08 | 0.104 |
| Urea, mmol/L | 3.97 | 4.50 | 0.58 | 0.543 |
| Glucose, mmol/L | 4.46 | 4.04 | 0.58 | 0.621 |
| Triglyceride, mmol/L | 0.48 | 0.49 | 0.04 | 0.908 |
| Creatinine, μmol/L | 83.2 | 83.9 | 5.33 | 0.919 |
| Item | NP | LP | SEM | p-Value |
|---|---|---|---|---|
| Day 14 of the experiment | ||||
| Observed species | 377 | 420 | 30.3 | 0.345 |
| ACE | 427 | 460 | 25.1 | 0.385 |
| Chao1 | 503 | 591 | 37.0 | 0.143 |
| Shannon | 5.27 | 5.25 | 0.18 | 0.951 |
| Simpson | 0.92 | 0.92 | 0.01 | 0.872 |
| End of the experiment | ||||
| Observed species | 636 | 706 | 16.6 | 0.017 |
| ACE | 656 | 729 | 18.6 | 0.025 |
| Chao1 | 643 | 714 | 17.6 | 0.022 |
| Shannon | 6.28 | 6.28 | 0.14 | 0.996 |
| Simpson | 0.96 | 0.96 | 0.01 | 0.681 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, L.; Wang, L.; Qiu, Y.; Xiong, Y.; Xiao, H.; Yi, H.; Wen, X.; Lin, Z.; Wang, Z.; Yang, X.; et al. Effects of Protein Restriction and Subsequent Realimentation on Body Composition, Gut Microbiota and Metabolite Profiles in Weaned Piglets. Animals 2021, 11, 686. https://doi.org/10.3390/ani11030686
Hou L, Wang L, Qiu Y, Xiong Y, Xiao H, Yi H, Wen X, Lin Z, Wang Z, Yang X, et al. Effects of Protein Restriction and Subsequent Realimentation on Body Composition, Gut Microbiota and Metabolite Profiles in Weaned Piglets. Animals. 2021; 11(3):686. https://doi.org/10.3390/ani11030686
Chicago/Turabian StyleHou, Lei, Li Wang, Yueqin Qiu, YunXia Xiong, Hao Xiao, Hongbo Yi, Xiaolu Wen, Zeling Lin, Zhikang Wang, Xuefen Yang, and et al. 2021. "Effects of Protein Restriction and Subsequent Realimentation on Body Composition, Gut Microbiota and Metabolite Profiles in Weaned Piglets" Animals 11, no. 3: 686. https://doi.org/10.3390/ani11030686