
Effects of Feed Supplementation on Nesfatin-1, Insulin, Glucagon, Leptin, T3, Cortisol, and BCS in Milking Ewes Grazing on Semi-Natural Pastures

 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Location, Animals, and Diets
2.2. BCS, Blood Collection, and Assays
2.3. Statistical Analysis
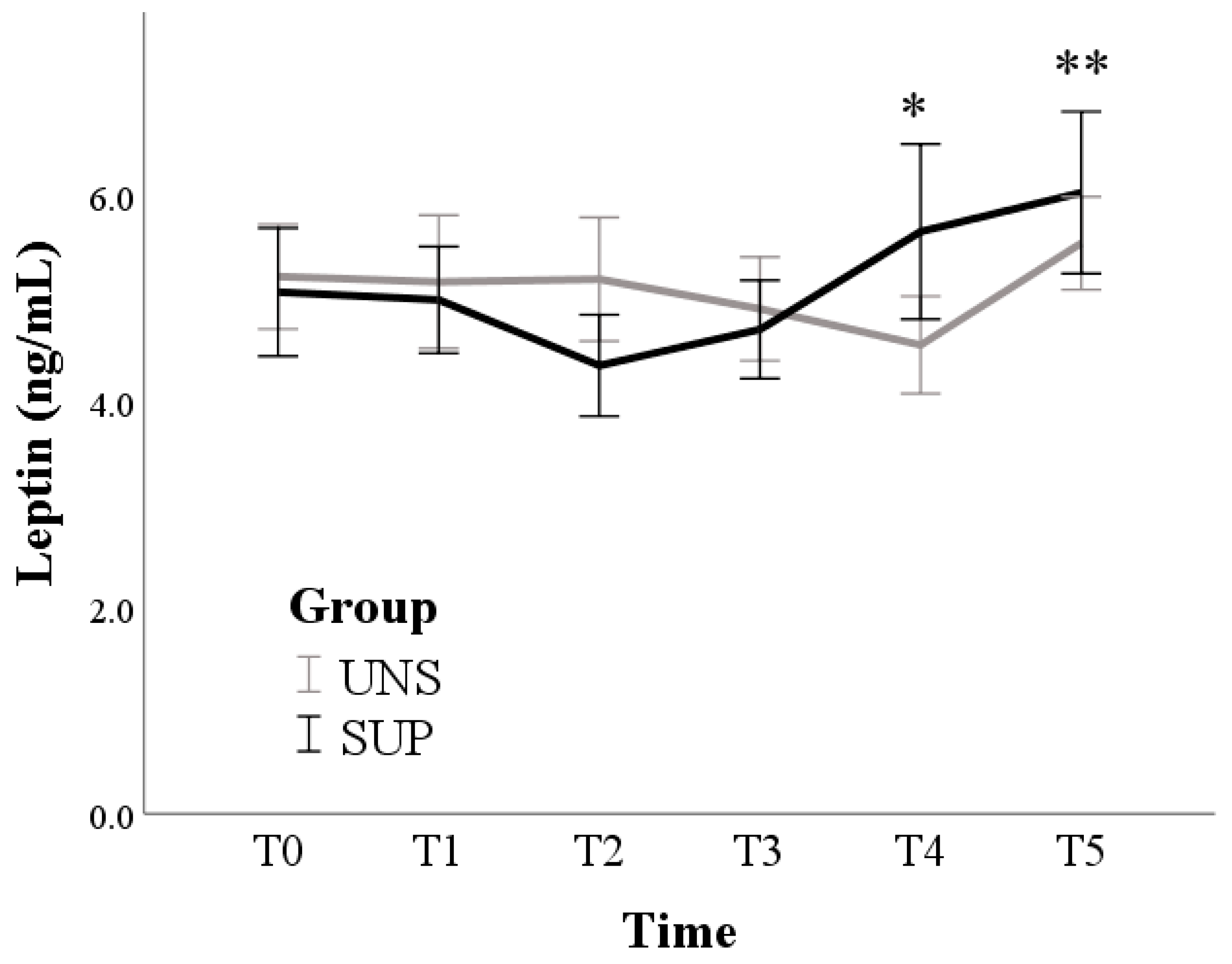
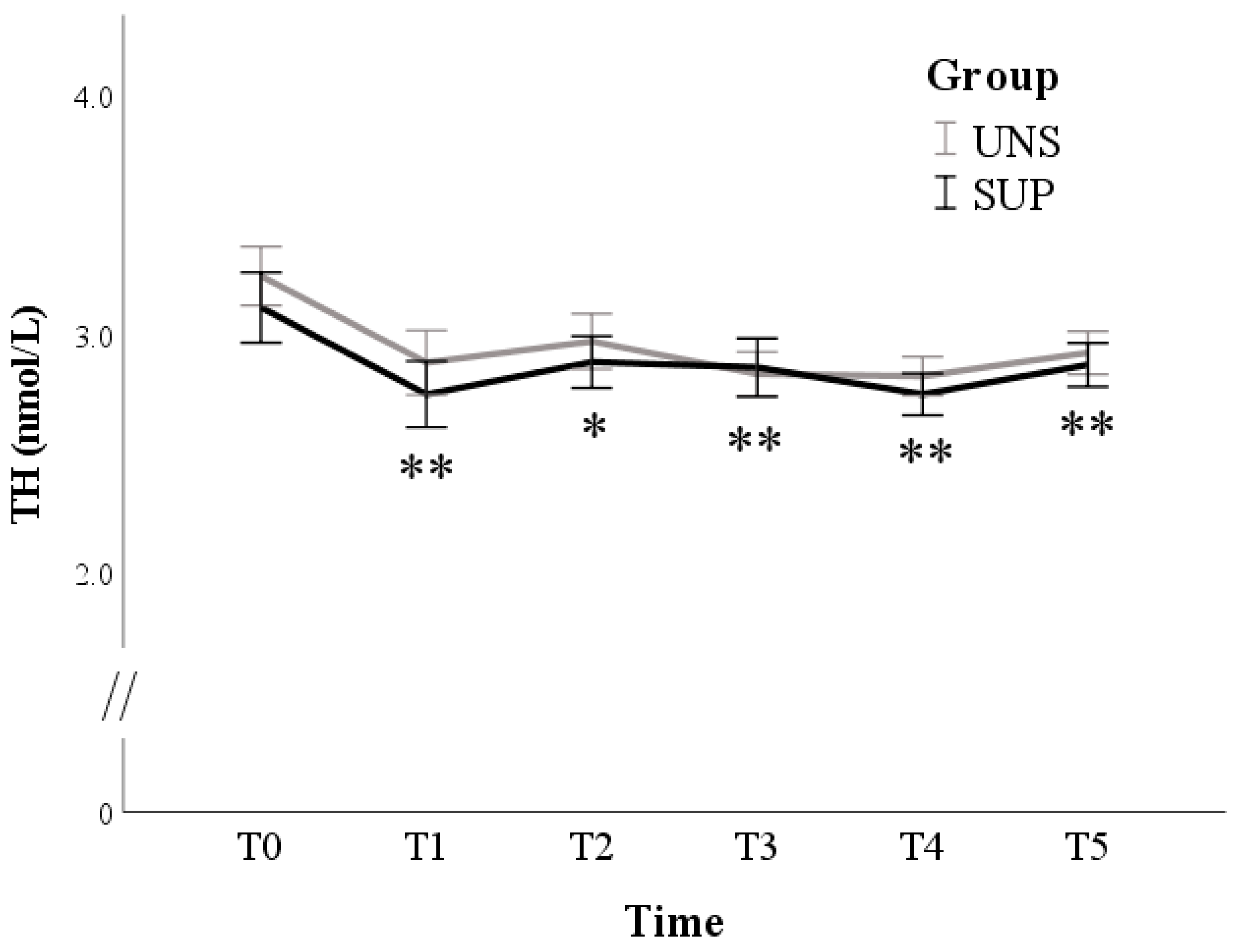
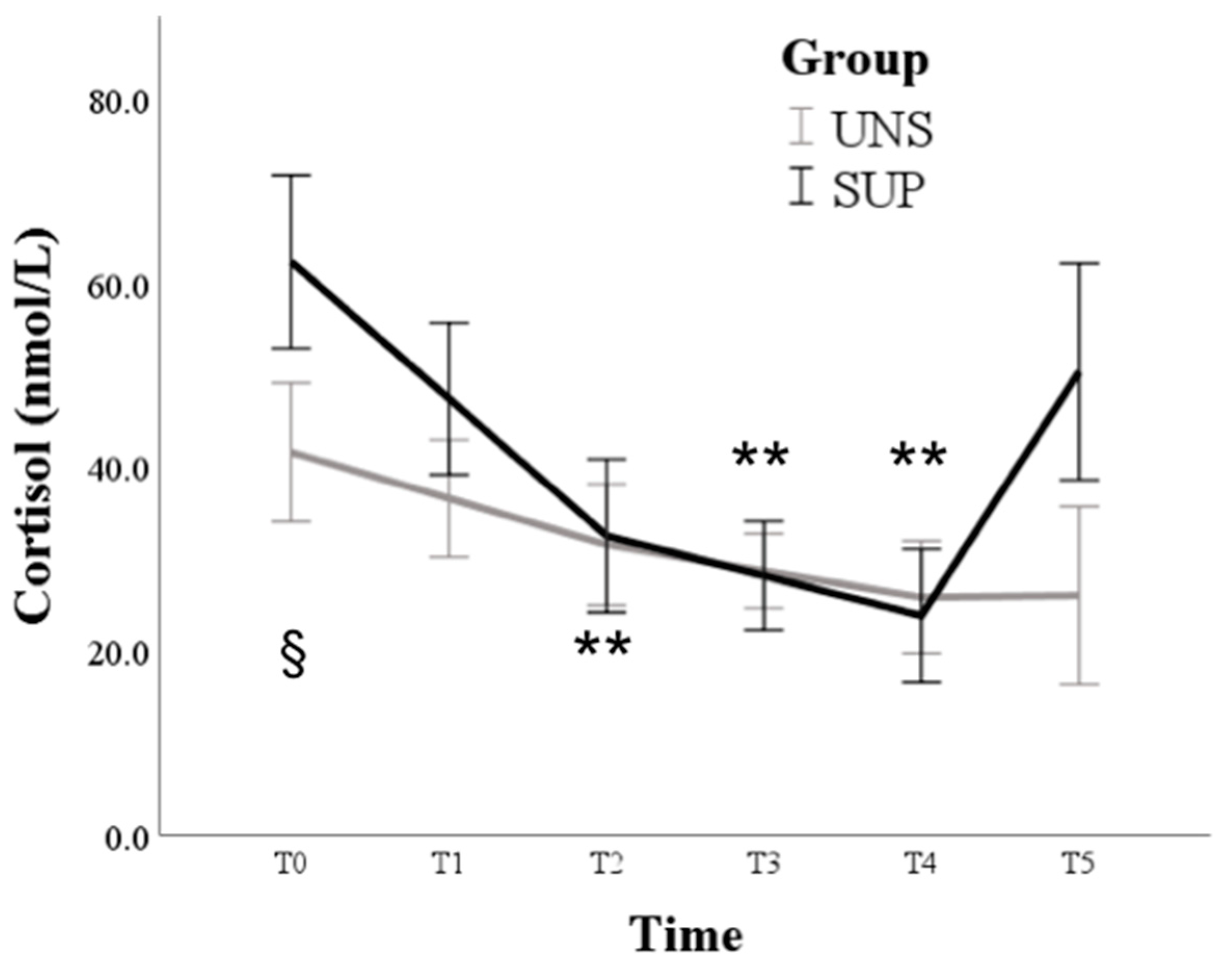
3. Results
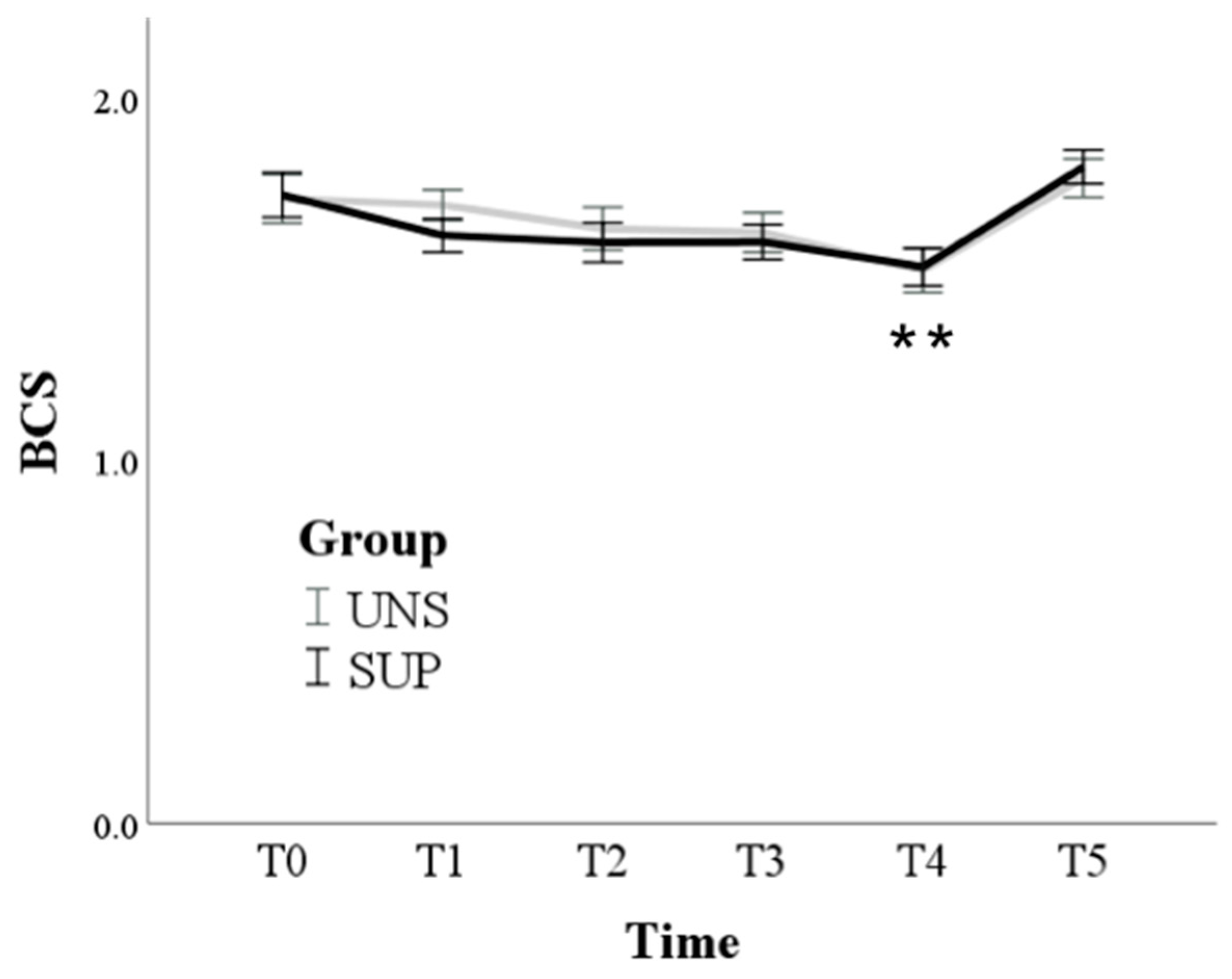
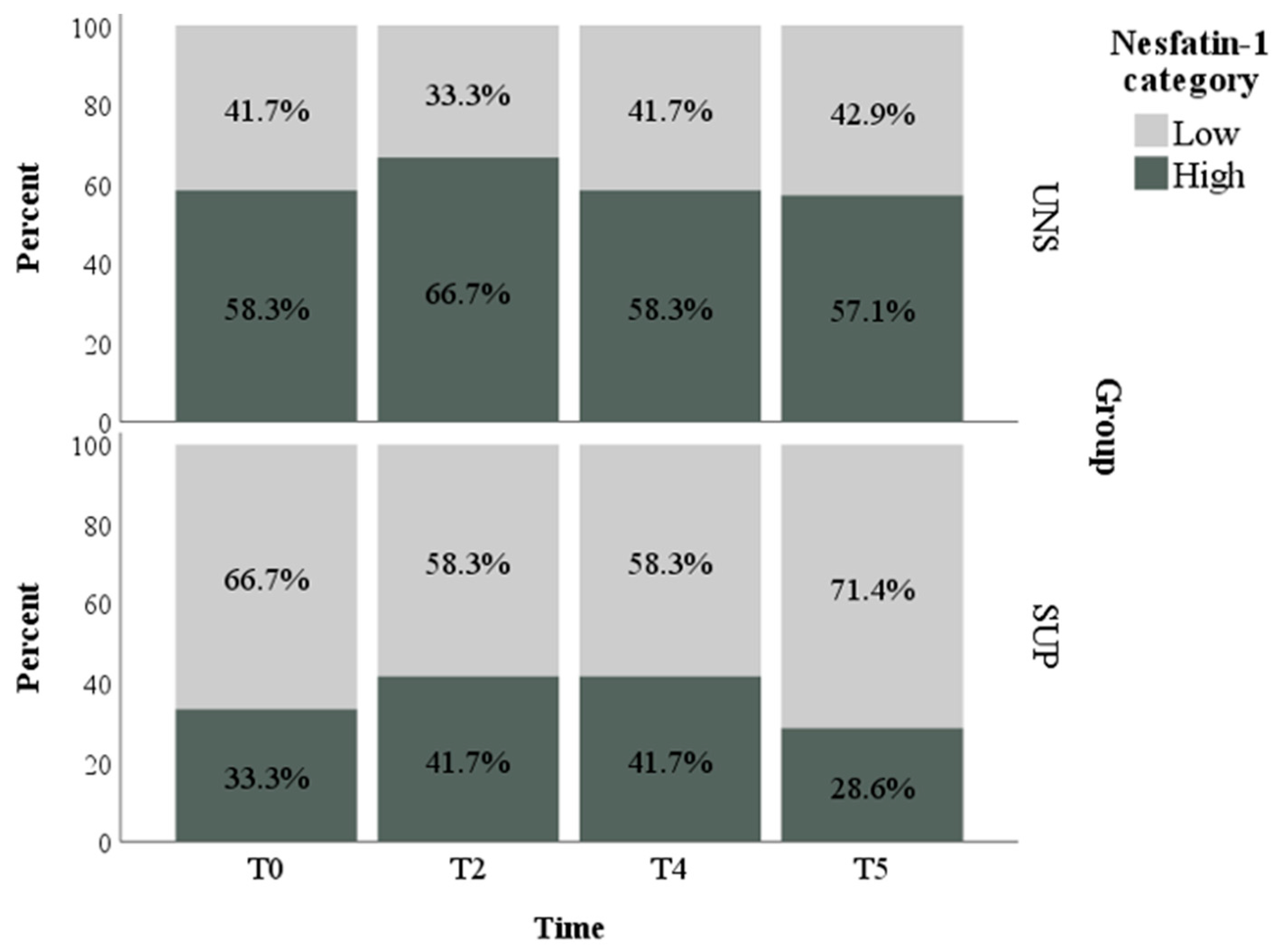
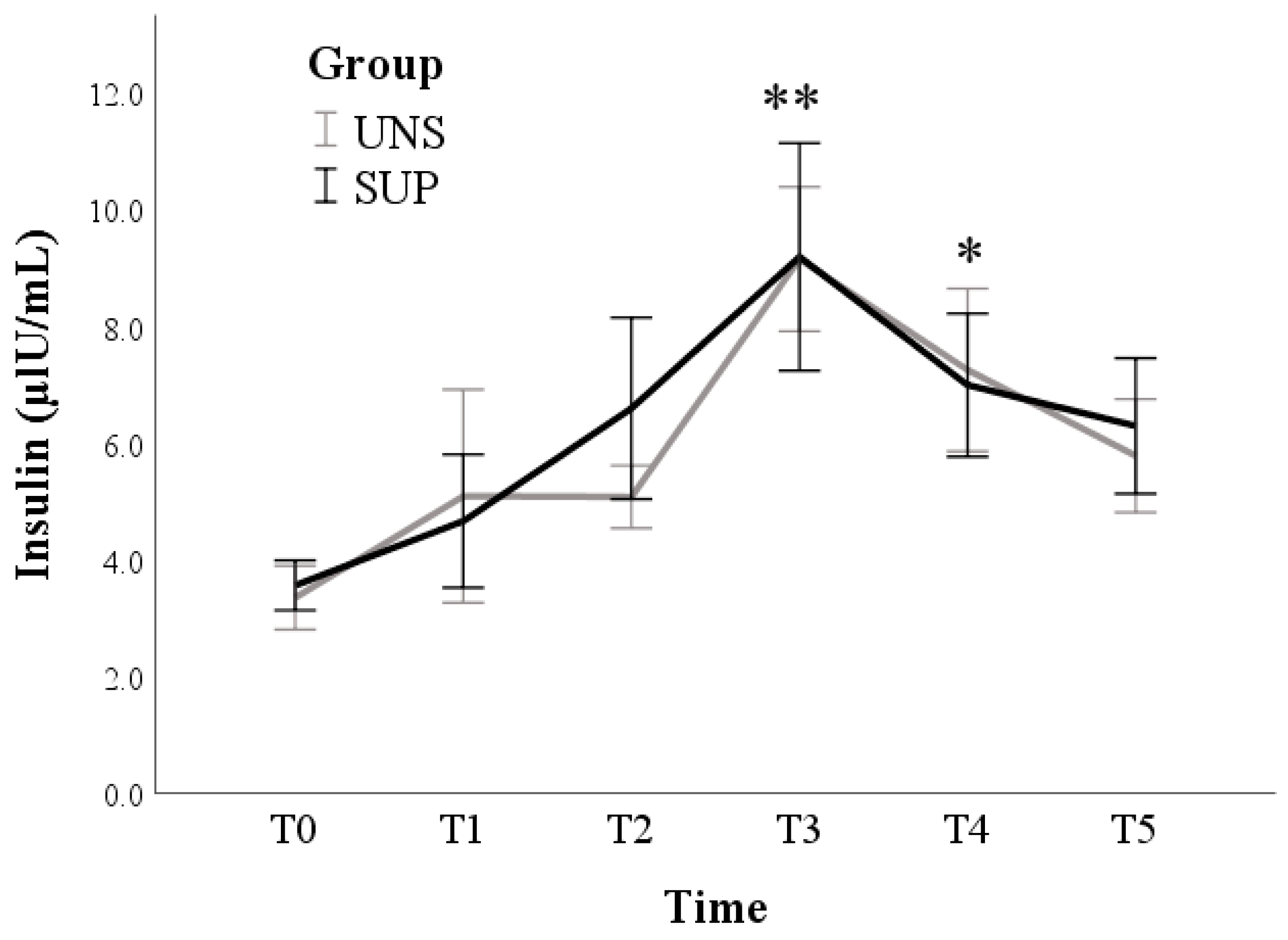
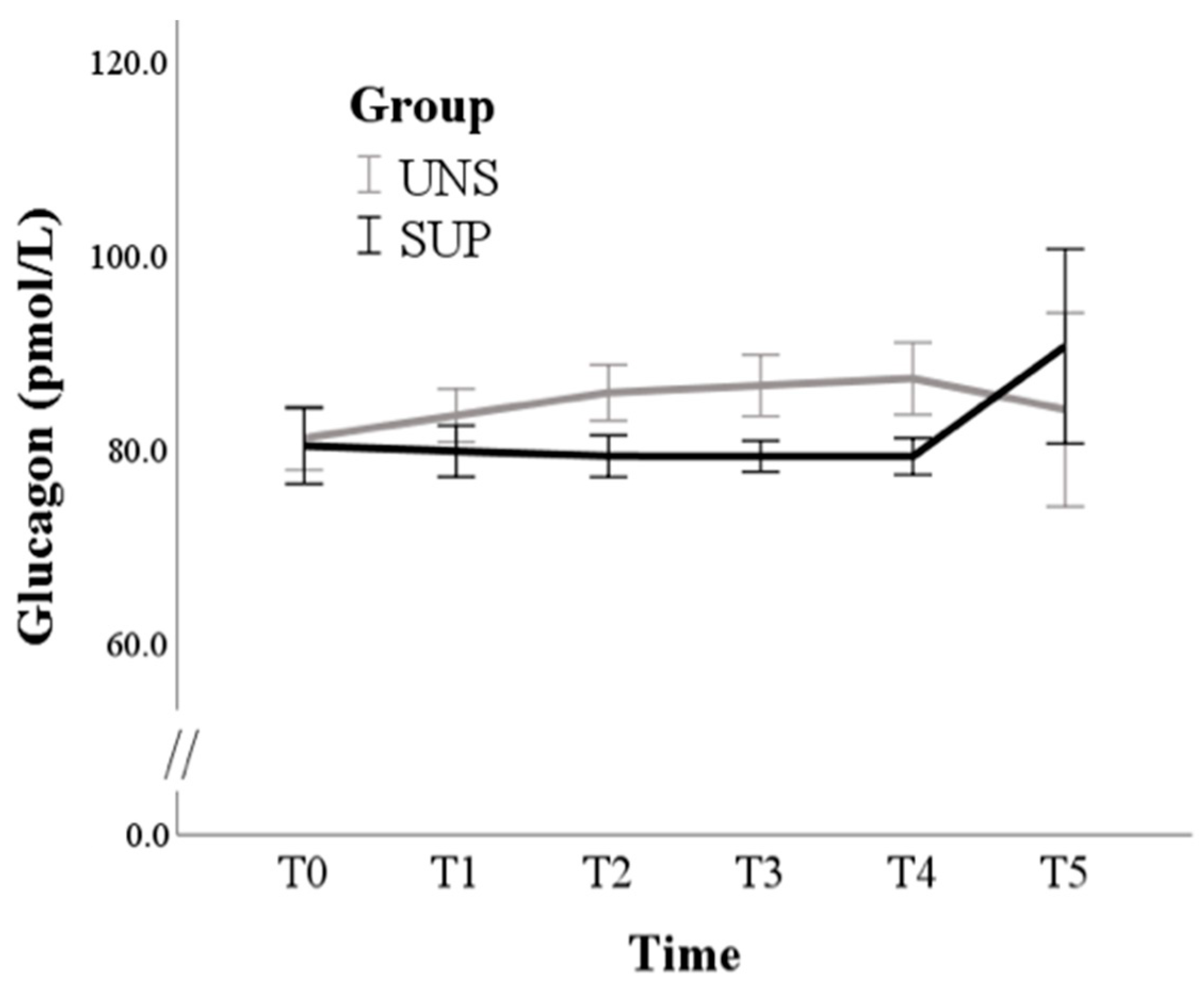
3.1. Univariable Approach
3.2. Multivariable Approach
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scocco, P.; Piermarteri, K.; Malfatti, A.; Tardella, F.M.; Catorci, A. Effects of summer rainfall variations on sheep body state and farming sustainability in sub-Mediterranean pastoral systems. Span. J. Agric. Res. 2016, 14, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Scocco, P.; Piermarteri, K.; Malfatti, A.; Tardella, F.M.; Catorci, A. Increase of drought stress negatively affects the sustainability of extensive sheep farming in sub-Mediterranean climate. J. Arid. Environ. 2016, 128, 50–58. [Google Scholar] [CrossRef]
- Scocco, P.; Mercati, F.; Tardella, F.M.; Catorci, A. Increase of forage dryness indices differentiated anatomical response in the sheep rumen compartment. Microsc. Res. Tech. 2016, 79, 738–743. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, F.; Lionello, P. Climate change projections for the Mediterranean region. Glob. Planet. Chang. 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Arzeni, A.; Storti, D. Le strategie per lo sviluppo rurale nelle Aree interne colpite dal sisma. Agriregionieuropa 2017, 51, 1–8. [Google Scholar]
- Robinson, J.; Ashworth, C.; Ashfort, J.; Rooke, L.; McEvoy, T. Nutrition and fertility in ruminant livestock. Anim. Feed Sci. Technol. 2006, 126, 259–276. [Google Scholar] [CrossRef]
- Dore, R.; Levata, L.; Lehnert, H.; Schulz, C. Nesfatin-1: Functions and physiology of a novel regulatory peptide. J. Endocrinol. 2017, 232, R45–R65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goebel, M.; Stengel, A.; Wang, A.; Tachè, Y. Central nesfatin-1 reduces the nocturnal food intake in mice be reducing meal size and increasing inter-meal intervals. Peptides 2011, 32, 36–43. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Gao, L.; Tang, H.; Yin, Y.; Zao, J.; Mulholland, M.; Zangh, W. Peripheral effects of nesfatin-1 on glucose homeostasis. PLoS ONE 2013, 8, e71513. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, N.; Maejima, Y.; Sedbazar, U.; Ando, A.; Kurita, H.; Damdinorj, B.; Takano, E.; Gantulga, D.; Iwasaki, Y.; Kurashina, T. Stressor-responsive central nesfatin-1 activates corticotropin-realising hormone, noradrenaline and serotonine neurons and evokes hipotalamic-pituitary-adrenal axis. Aging 2010, 2, 775–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vas, S.; Adori, C.; Konczol, K.; Katai, Z.; Pap, D.; Papp, R.S.; Bagdy, G.; Palkovitis, M.; Thot, Z.E. Nesfatin-1/NUCB2 as a potential new element of sleep regulation in rats. PLoS ONE 2013, 6, e59809. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Zhang, K.; Song, M.; Li, X.; Luo, L.; Tian, Y.; Zhang, Y.; Li, Y.; Zhangh, X.; Ling, Y. Role of nesfatin-1 in the reproductive axis of male rat. Sci. Rep. 2016, 6, 32877. [Google Scholar] [CrossRef] [PubMed]
- Prajapati, G.N.; Kumar, A.; Laxmi, A.N.; Singh, N.K.; Mathur, A.K. Parallelism of nesfatin-1 and leptin with metabolic parameters and progesterone at puberty in Murrah buffalo heifers. J. Anim. Res. 2018, 7, 1–6. [Google Scholar]
- Laarveld, B.; Christensen, D.A.; Brockman, R.P. The effect of insulin on net metabolism of glucose and amino acids by the bovine mammary gland. Endocrinology 1981, 108, 2217–2221. [Google Scholar] [CrossRef] [PubMed]
- Todini, L.; Trabalza-Marinucci, M.; Malfatti, A.; Barbato, O.; Stradaioli, G.; Cavallucci, C.; Debenedetti, A. Pre- and post-feeding plasma Gastrin-17 and Insulin concentrations and feed intake in female goats during different physiological stages. Small Rumin. Res. 2007, 71, 38–47. [Google Scholar] [CrossRef]
- Block, S.S.; Rhoads, R.P.; Bauman, D.E.; Ehrhardt, R.A.; McGuire, M.A.; Crooker, B.A.; Griinari, J.M.; Mackle, T.R.; Weber, W.J.; Amburgh, M.E.; et al. Demonstration of a role for insulin in the regulation of leptin in lactating dairy cows. J. Dairy Sci. 2003, 86, 3508–3515. [Google Scholar] [CrossRef]
- Jiang, G.; Zhang, B.B. Glucagon and regulation of glucose metabolism. Am. J. Physiol. Metab. 2003, 284, E671–E678. [Google Scholar] [CrossRef] [Green Version]
- Vaughan, M.; Steinberg, G.D. Effect of hormones on lipolysis and esterification of free fatty acid during incubation of adipose tissue in vitro. J. Lipid Res. 1963, 4, 193–199. [Google Scholar] [CrossRef]
- Woods, S.C.; Lutz, T.A.; Geary, N.; Langhans, W. Pancreatic signals controlling food intake; Isulin, Glucagon and Amilyn. Philos. Trans. R. Soc. B Biol. Sci. 2006, 311, 1219–1235. [Google Scholar] [CrossRef] [Green Version]
- Houseknecht, K.L.; Baile, C.A.; Matteri, R.L.; Spurlock, M.E. The biology of leptin: A review. J. Anim. Sci. 1998, 76, 1405–1420. [Google Scholar] [CrossRef] [Green Version]
- Krasnow, S.M.; Steiner, R.A. Physiological Mechanisms Integrating Metabolism and Reproduction. In Knobil and Neill’s Physiology of Reproduction; Neill, J.D., Ed.; Academic Press: San Diego, CA, USA, 1998; pp. 2553–2626. [Google Scholar]
- Blache, D.; Tellam, R.L.; Chagas, L.M.; Blackberry, M.A.; Vercoe, P.E.; Martin, G.B. Level of nutrition affects leptin concentrations in plasma and cerebrospinal fluid in sheep. J. Endocrinol. 2000, 165, 625–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laud, K.; Gourdou, I.; Belair, L.; Keisler, D.H.; Djiane, J. Detection and regulation of leptin receptor mRNA in ovine mammary epithelial cells during pregnancy and lactation. FEBS Lett. 1999, 463, 194–198. [Google Scholar] [CrossRef] [Green Version]
- Todini, L. Thyroid hormones in small ruminants: Effects of endogenous, environmental and nutritional factors. Animal 2007, 1, 997–1008. [Google Scholar] [CrossRef] [Green Version]
- Maurya, V.P.; Sejian, V.; Kumar, D.; Naqvi, S.M.K. Impact of heat stress, nutritional stress and their combinations on the adaptive capability of Malpura sheep under hot semi-arid tropical environment. J. Anim. Behav. Biometeorol. 2019, 7, 31–38. [Google Scholar]
- Caldeira, R.M.; Belo, A.T.; Santos, C.C.; Vazques, M.I.; Portugal, A.V. The effect of long-term feed restriction and over-nutrition on body condition score, blood metabolites and hormonal profiles in ewes. Small Rumin. Res. 2007, 68, 242–255. [Google Scholar] [CrossRef]
- Mercati, F.; Maranesi, M.; Dall’Aglio, C.; Petrucci, L.; Pasquariello, R.; Tardella, F.M.; de Felice, E.; Scocco, P. Apelin System in Mammary Gland of Sheep Reared in Semi-Natural Pastures of the Central Apennines. Animals 2018, 8, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scocco, P.; Rivaroli, S.; Mercati, F.; Tardella, F.M.; Malfatti, A.; de Felice, E.; Catorci, A. Anatomy for economy: Starting from the rumen keratinization degree to enhance the farm income. Econ. Agro Aliment. 2018, 20, 261–272. [Google Scholar] [CrossRef]
- Scocco, P.; Mercati, F.; Brusaferro, A.; Ceccarelli, P.; Belardinelli, C.; Malfatti, A. Keratinization degree of rumen epithelium and body condition score in sheep grazing on Brachipodium rupestre. Vet. Ital. 2013, 49, 211–217. [Google Scholar]
- Altman, D.G.; Bland, J.J. Uncertainty and sampling error. BMJ 2014, 349, g7064. [Google Scholar] [CrossRef] [Green Version]
- Garson, G.D. Discriminant Function Analysis; Blue Book; Statistical Associates Publishers: Asheboro, NC, USA, 2012. [Google Scholar]
- Agradi, S.; Curone, G.; Negroni, D.; Vigo, D.; Brecchia, G.; Bronzo, V.; Chiesa, L.; Peric, T.; Danes, D.; Menchetti, L. Determination of fatty acids profile in original brown cows dairy products and relationship with alpine pasture farming system. Animals 2020, 10, 1231. [Google Scholar] [CrossRef]
- Herbein, J.H.; Aiello, R.J.; Eckler, L.I.; Pearson, R.E.; Akers, R.M. Glucagon, Insulin, Growth Hormone, and Glucose Concentrations in Blood Plasma of Lactating Dairy Cows. J. Dairy Sci. 1985, 68, 320–325. [Google Scholar] [CrossRef]
- Brockman, R.P.; Laarveld, B. Hormonal regulation of metabolism in ruminants: A review. Livest. Prod. Sci. 1986, 14, 313–334. [Google Scholar] [CrossRef]
- Antunovic, Z.; Novoselec, J.; Sauerwein, H.; Speranda, M.; Vegara, M.; Pavic, V. Blood metabolic profile and some of hormones concentration in ewes during different physiological status. Bulg. J. Agric. Sci. 2011, 17, 687–695. [Google Scholar]
- Ehrhardt, R.A.; Slepetis, R.M.; Bell, A.W.; Boisclair, Y.R. Maternal leptin is elevated during pregnancy in sheep. Domest. Anim. Endocrinol. 2001, 21, 85–96. [Google Scholar] [CrossRef]
- Sorensen, A.; Adam, C.L.; Findlay, P.A.; Marie, M.; Thomas, L.; Travers, M.T.; Vernon, R.G. Leptin secretion and hypothalamic neuropeptide and receptor gene expression in sheep. Am. J. Physiol. Integr. Comp. Physiol. 2002, 282, R1227–R1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-García, E.; Tesniere, A.; Camous, S.; Bocquier, F.; Barillet, F.; Hassoun, P. The effects of parity, litter size, physiological state, and milking frequency on the metabolic profile of Lacaune dairy ewes. Domest. Anim. Endocrinol. 2015, 50, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Bekeova, E.; Elecko, J.; Krajnicakova, M.; Hendrichovsky, V.; Maracek, I. Dynamics of changes in concentrations of cholesterol and thyroid and ovarian hormones in blood-serum during postparturient period of ewes. Vet. Med. UVTIZ CSFR 1991, 36, 673–684. [Google Scholar]
- Antunović, Z.; Novoselec, J.; Speranda, M.; Vegara, M.; Pavić, V.; Mioč, B.; Djidara, M. Changes in biochemical and hematological parameters and metabolic hormones in Tsigai ewes blood in the first third of lactation. Arch. Anim. Breed. 2011, 4, 535–545. [Google Scholar] [CrossRef]
- Todini, L.; Malfatti, A.; Valbonesi, A.; Trabalza-Marinucci, M.; Debenedetti, A. Plasma total T3 and T4 concentrations in goats at different physiological stages, as affected by the energy intake. Small Rumin. Res. 2007, 68, 285–290. [Google Scholar] [CrossRef]
- Da Silva, W.E.; Leite, J.H.G.M.; da Silva, W.S.T.; Paiva, R.D.M.; de Sousa, J.E.R.; Facanha, D.A.E. Seasonal variations in thermoregulatory patterns enable Morada Nova sheep to adapt to Brazilian semi-arid. Semin. Agrar. Sci. 2019, 40, 1577–1594. [Google Scholar] [CrossRef] [Green Version]
- Rathwa, S.D.; Vasava, A.A.; Pathan, M.M.; Madhira, S.P.; Patel, Y.G.; Pande, A.M. Effect of season on physiological, biochemical, hormonal, and oxidative stress parameters of indigenous sheep. Vet. World 2017, 10, 650–654. [Google Scholar] [CrossRef] [Green Version]
- Menchetti, L.; Curone, G.; Andoni, E.; Barbato, O.; Troisi, A.; Fioretti, B.; Polisca, A.; Codini, M.; Canali, C.; Vigo, D.; et al. Impact of Goji Berries (Lycium barbarum) Supplementation on the Energy Homeostasis of Rabbit Does: Uni- and Multivariate Approach. Animals 2020, 10, 2000. [Google Scholar] [CrossRef]
- Gupta, V.K.; Sharma, S.D.; Vihan, V.S.; Kumar, A. Serum enzymes and thyroid hormone in sub-clinical ketosis in goats and sheep reared under organized farming system. Ind. J. Anim. Sci. 2008, 78, 1199–1201. [Google Scholar]
- Ribeiro, M.N.; Ribeiro, N.L.; Bozzi, R.; Costa, R.G. Physiological and biochemical blood variables of goats subjected to heat stress—A review. J. Appl. Anim. Res. 2008, 46, 1036–1041. [Google Scholar] [CrossRef] [Green Version]
- Maurya, V.P.; Sejian, V.; Kumar, D.; Naqvi, S.M.K. Biological ability of Malpura rams to counter heat stress challenges and its consequences on production performance in a semi-arid tropical environment. Biol. Rhythm. Res. 2018, 49, 479–493. [Google Scholar] [CrossRef]
- Inbaraj, S.; Kundu, A.; De, A.K.; Sunder, J.; Sejian, V. Seasonal changes in blood biochemical and endocrine responses of different indigenous goat breeds of tropical island agro-ecological environment. Biol. Rhythm. Res. 2018, 49, 412–421. [Google Scholar] [CrossRef]
- Yilmaz, M.; Altin, T.; Kiral, F.; Taskin, T.; Asici, G.S.E.; Kizilkaya, K. Effects of the season on physiological and endocrine traits and on HSP70 in Saanen goats under Mediterranean climate conditions. Med. Weter. 2018, 74, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Sejian, V.; Indu, S.; Naqvi, S.M.K. Impact of short term exposure to different environmental temperature on the blood biochemical and endocrine responses of Malpura ewes under semi-arid tropical environment. Ind. J. Anim. Sci. 2013, 83, 1155–1159. [Google Scholar]
- Sejian, V.; Maurya, V.P.; Naqvi, S.M.K. Adaptive capability as indicated by endocrine and biochemical responses of Malpura ewes subjected to combined stresses (thermal and nutritional) in a semi-arid tropical environment. Int. J. Biometeorol. 2010, 54, 653–661. [Google Scholar] [CrossRef]
- Dall’Aglio, C.; Scocco, P.; Maranesi, M.; Petrucci, L.; Acuti, G.; de Felice, E.; Mercati, F. Immunohistochemical identification of resistin in the uterus of ewes subjected to different diets: Preliminary results. Eur. J. Histochem. 2019, 63, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Mercati, F.; Scocco, P.; Maranesi, M.; Acuti, G.; Petrucci, L.; Cocci, P.; Renzi, A.; de Felice, E.; Dall’Aglio, C. Apelin system detection in the reproductive apparatus of ewes grazing on semi-natural pasture. Theriogenology 2019, 139, 156–166. [Google Scholar] [CrossRef]
- Scocco, P.; Mercati, F.; Catorci, A.; Maranesi, M.; Acuti, G.; de Felice, E.; Petrucci, L.; Dall’Aglio, C. Histochemistry as supporting tool in grassland ecosystem management: Apelin system detection in ewe reproductive apparatus. Eur. J. Histochem. 2019, 63, 31. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Discriminant Loadings | Wilks’ Lambda | p Value |
|---|---|---|---|
| BCS | −0.566 | 0.880 | 0.018 |
| Nesfatin-1 * | 0.515 | 0.899 | 0.031 |
| Cortisol | −0.249 | 0.974 | 0.288 |
| TH | 0.126 | 0.993 | 0.588 |
| Insulin # | 0.078 | 0.997 | 0.737 |
| Leptin | 0.042 | 0.999 | 0.857 |
| Glucagon | 0.037 | 0.999 | 0.875 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbato, O.; De Felice, E.; Todini, L.; Menchetti, L.; Malfatti, A.; Scocco, P. Effects of Feed Supplementation on Nesfatin-1, Insulin, Glucagon, Leptin, T3, Cortisol, and BCS in Milking Ewes Grazing on Semi-Natural Pastures. Animals 2021, 11, 682. https://doi.org/10.3390/ani11030682
Barbato O, De Felice E, Todini L, Menchetti L, Malfatti A, Scocco P. Effects of Feed Supplementation on Nesfatin-1, Insulin, Glucagon, Leptin, T3, Cortisol, and BCS in Milking Ewes Grazing on Semi-Natural Pastures. Animals. 2021; 11(3):682. https://doi.org/10.3390/ani11030682
Chicago/Turabian StyleBarbato, Olimpia, Elena De Felice, Luca Todini, Laura Menchetti, Alessandro Malfatti, and Paola Scocco. 2021. "Effects of Feed Supplementation on Nesfatin-1, Insulin, Glucagon, Leptin, T3, Cortisol, and BCS in Milking Ewes Grazing on Semi-Natural Pastures" Animals 11, no. 3: 682. https://doi.org/10.3390/ani11030682