
Effect of a Phytogenic Water Additive on Growth Performance, Blood Metabolites and Gene Expression of Amino Acid Transporters in Nursery Pigs Fed with Low-Protein/High-Carbohydrate Diets


 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Housing, and Diets
2.2. Growth Performance
2.3. Feed and Fecal Samples Collection
2.4. Blood and Tissue Samples Collection
2.5. Feed and Fecal Samples Composition Analysis
2.6. Plasma Nitrogen-Containing Compounds Analysis
2.7. Muscle Composition Analysis
2.8. Serum Metabolites Analysis
2.9. RNA Isolation and RT-qPCR Analysis
2.10. Total Antioxidant Capacity
2.11. Apparent Fecal Digestibility of Nutrients
2.12. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Muscle Fat and Lean Content
3.3. Plasma Nitrogen-Containing Compounds
3.4. Apparent Fecal Digestibility of Calcium, Phosphorus, and Nitrogen
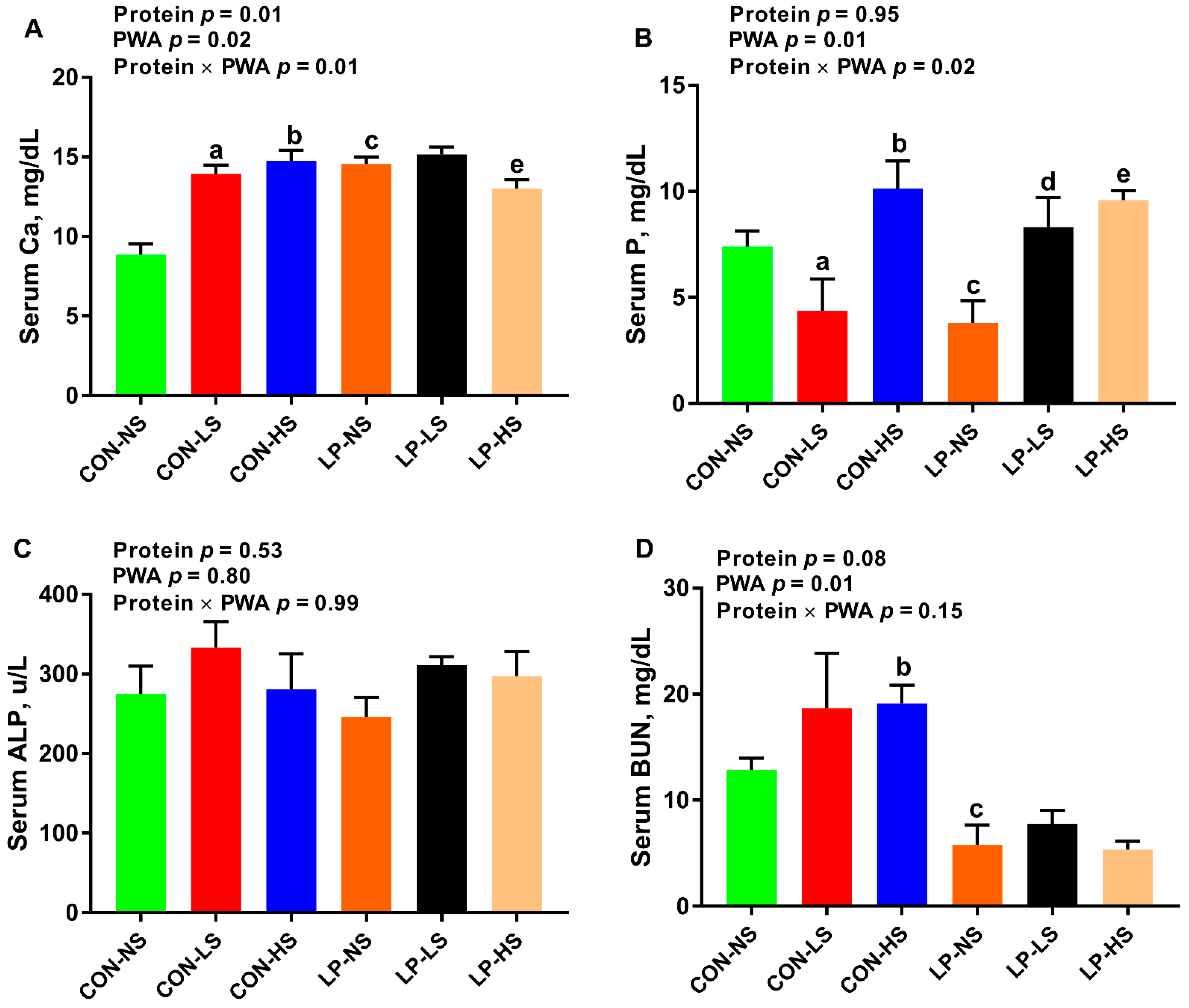
3.5. Serum Calcium, Phosphorus, and Alkaline Phosphatase and Blood Urea Nitrogen
3.6. The mRNA Abundance of Amino Acid Transporters in the Jejunum and Skeletal Muscle
3.7. Serum Total Antioxidant Capacity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aarnink, A.J.A.; Verstegen, M.W.A. Nutrition, Key Factor to Reduce Environmental Load from Pig Production. Livest. Sci. 2007, 109, 194–203. [Google Scholar] [CrossRef]
- Jongbloed, A.W.; Lenis, N.P. Environmental Concerns about Animal Manure. J. Anim. Sci. 1998, 76, 2641–2648. [Google Scholar] [CrossRef]
- Shriver, J.A.; Carter, S.D.; Sutton, A.L.; Richert, B.T.; Senne, B.W.; Pettey, L.A. Effects of Adding Fiber Sources to Reduced-Crude Protein, Amino Acid-Supplemented Diets on Nitrogen Excretion, Growth Performance, and Carcass Traits of Finishing Pigs. J. Anim. Sci. 2003, 81, 492–502. [Google Scholar] [CrossRef] [Green Version]
- Heo, J.-M.; Kim, J.-C.; Hansen, C.F.; Mullan, B.P.; Hampson, D.J.; Pluske, J.R. Effects of Feeding Low Protein Diets to Piglets on Plasma Urea Nitrogen, Faecal Ammonia Nitrogen, the Incidence of Diarrhoea and Performance after Weaning. Arch. Anim. Nutr. 2008, 62, 343–358. [Google Scholar] [CrossRef]
- Le Bellego, L.; Noblet, J. Performance and Utilization of Dietary Energy and Amino Acids in Piglets Fed Low Protein Diets. Livest. Prod. Sci. 2002, 76, 45–58. [Google Scholar] [CrossRef]
- Ball, R.O.; Aherne, F.X. Influence of dietary nutrient density, level of feed intake and weaning age on young pigs. II. Apparent nutrient Digestibility and Incidence and Severity of Diarrhea. Can. J. Anim. Sci. 1987, 67, 1105–1115. [Google Scholar] [CrossRef]
- Yue, L.Y.; Qiao, S.Y. Effects of Low-Protein Diets Supplemented with Crystalline Amino Acids on Performance and Intestinal Development in Piglets over the First 2 Weeks after Weaning. Livest. Sci. 2008, 115, 144–152. [Google Scholar] [CrossRef]
- Kerr, B.J.; Yen, J.T.; Nienaber, J.A.; Easter, R.A. Influences of dietary protein level, amino acid supplementation and environmental temperature on performance, body composition, organ weights and total heat production of growing pigs. J. Anim. Sci. 2003, 81, 1998–2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madrid, J.; Martínez, S.; López, C.; Orengo, J.; López, M.J.; Hernández, F. Effects of low protein diets on growth performance, carcass traits and ammonia emission of barrows and gilts. Anim. Prod. Sci. 2013, 53, 146–153. [Google Scholar] [CrossRef]
- Bartoš, P.; Dolan, A.; Smutný, L.; Šístková, M.; Celjak, I.; Šoch, M.; Havelka, Z. Effects of Phytogenic Feed Additives on Growth Performance and on Ammonia and Greenhouse Gases Emissions in Growing-Finishing Pigs. Anim. Feed Sci. Technol. 2016, 212, 143–148. [Google Scholar] [CrossRef]
- Karásková, K.; Suchý, P.; Straková, E. Current Use of Phytogenic Feed Additives in Animal Nutrition: A Review. Czech J. Anim. Sci. 2015, 60, 521–530. [Google Scholar] [CrossRef] [Green Version]
- Papatsiros, V.G.; Tzika, E.D.; Tassis, P.D.; Kantas, D.; Filippopoulos, L.C.; Papaioannou, D.S. Greek Experience of the Use of Phytogenic Feed Additives in Organic Pig Farming. J. Cell Anim. Biol. 2011, 5, 320–323. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.; Zhang, S.; Wang, H.; Piao, X. Essential Oil and Aromatic Plants as Feed Additives in Non-Ruminant Nutrition: A Review. J. Anim. Sci. Biotech. 2015, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Windisch, W.; Schedle, K.; Plitzner, C.; Kroismayr, A. Use of Phytogenic Products as Feed Additives for Swine and Poultry. J. Anim. Sci. 2008, 86 (Suppl. 14), E140–E148. [Google Scholar] [CrossRef]
- Krauze, M.; Abramowicz, K.; Ognik, K. The Effect of Addition of Probiotic Bacteria (Bacillus subtilis or Enterococcus faecium) or Phytobiotic Containing Cinnamon Oil to Drinking Water on the Heath and Performance of Broiler Chickens. Ann. Anim. Sci. 2020, 20, 191–205. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.F.; Wu, G.Y.; Liao, Y.P.; Hou, Z.P.; Liu, H.J.; Yin, F.G.; Li, T.J.; Huang, R.L.; Zhang, Y.M.; Deng, D.; et al. Effects of Chinese Herbal Ultra-Fine Powder as a Dietary Additive on Growth Performance, Serum Metabolites and Intestinal Health in Early-Weaned Piglets. Livest. Sci. 2007, 108, 272–275. [Google Scholar] [CrossRef]
- Yan, L.; Meng, Q.W.; Kim, I.H. The Effect of an Herb Extract Mixture on Growth Performance, Nutrient Digestibility, Blood Characteristics and Fecal Noxious Gas Content in Growing Pigs. Livest. Sci. 2011, 141, 143–147. [Google Scholar] [CrossRef]
- Manzanilla, E.G.; Pérez, J.F.; Martín, M.; Blandón, J.C.; Baucells, F.; Kamel, C.; Gasa, J. Dietary Protein Modifies Effect of Plant Extracts in the Intestinal Ecosystem of the Pig at Weaning. J. Anim. Sci. 2009, 87, 2029–2037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abousekken, M. Performance, Immune Response and Carcass Quality of Broilers Fed Low Protein Diets Contained Either Moringa oleifera Leaves Meal or Its Extract. J. Am. Sci. 2015, 11, 153–164. [Google Scholar]
- Lei, X.J.; Lee, S.I.; Kim, I.H. Effects of Different Levels of Dietary Protein with or without Plant Extract YGF251 on Growth Performance, Nutrient Digestibility, Blood Profiles, Fecal Microbial Shedding, and Fecal Gas Emission in Growing Pigs. Anim. Sci. J. 2019, 90, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Wenk, C. Herbs and Botanicals as Feed Additives in Monogastric Animals. Asian Australas. J. Anim. Sci. 2003, 16, 282–289. [Google Scholar] [CrossRef]
- Nofrarias, M.; Manzanilla, E.G.; Pujols, J.; Gibert, X.; Majo, N.; Segalés, J.; Gasa, J. Effects of spray-dried porcine plasma and plant extracts on intestinal morphology and on leukocyte cell subsets of weaned pigs. J. Anim. Sci. 2006, 84, 2735–2742. [Google Scholar] [CrossRef]
- Manzanilla, E.G.; Nofrarías, M.; Anguita, M.; Castillo, M.; Perez, J.F.; Martín-Orúe, S.M.; Kamel, C.; Gasa, J. Effects of Butyrate, Avilamycin, and a Plant Extract Combination on the Intestinal Equilibrium of Early-Weaned Pigs. J. Anim. Sci. 2006, 84, 2743–2751. [Google Scholar] [CrossRef]
- Li, P.; Piao, X.; Ru, Y.; Han, X.; Xue, L.; Zhang, H. Effects of Adding Essential Oil to the Diet of Weaned Pigs on Performance, Nutrient Utilization, Immune Response and Intestinal Health. Asian Australas. J. Anim. Sci. 2012, 25, 1617–1626. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Wang, J.P.; Kim, H.J.; Meng, Q.W.; Ao, X.; Hong, S.M.; Kim, I.H. Influence of Essential Oil Supplementation and Diets with Different Nutrient Densities on Growth Performance, Nutrient Digestibility, Blood Characteristics, Meat Quality and Fecal Noxious Gas Content in Grower-Finisher Pigs. Livest. Sci. 2010, 128, 115–122. [Google Scholar] [CrossRef]
- Diao, H.; Zheng, P.; Yu, B.; He, J.; Mao, X.; Yu, J.; Chen, D. Effects of Benzoic Acid and Thymol on Growth Performance and Gut Characteristics of Weaned Piglets. Asian Australas. J. Anim. Sci. 2015, 28, 827–839. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, C.J.; Martínez-Miró, S.; Ariza, J.J.; Madrid, J.; Orengo, J.; Aguinaga, M.A.; Baños, A.; Hernández, F. Effect of Alliaceae Extract Supplementation on Performance and Intestinal Microbiota of Growing-Finishing Pig. Animals 2020, 10, 1557. [Google Scholar] [CrossRef] [PubMed]
- Xue, P.C.; Ragland, D.; Adeola, O. Influence of Dietary Crude Protein and Phosphorus on Ileal Digestion of Phosphorus and Amino Acids in Growing Pigs. J. Anim. Sci. 2017, 95, 2071–2079. [Google Scholar] [CrossRef]
- Xue, P.C.; Ajuwon, K.M.; Adeola, O. Phosphorus and Nitrogen Utilization Responses of Broiler Chickens to Dietary Crude Protein and Phosphorus Levels. Poult. Sci. 2016, 95, 2615–2623. [Google Scholar] [CrossRef]
- Spring, S.; Premathilake, H.; DeSilva, U.; Shili, C.; Carter, S.; Pezeshki, A. Low Protein-High Carbohydrate Diets Alter Energy Balance, Gut Microbiota Composition and Blood Metabolomics Profile in Young Pigs. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Spring, S.; Premathilake, H.; Bradway, C.; Shili, C.; DeSilva, U.; Carter, S.; Pezeshki, A. Effect of Very Low-Protein Diets Supplemented with Branched-Chain Amino Acids on Energy Balance, Plasma Metabolomics and Fecal Microbiome of Pigs. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Zhang, S.; Qiao, S.; Ren, M.; Zeng, X.; Ma, X.; Wu, Z.; Thacker, P.; Wu, G. Supplementation with Branched-Chain Amino Acids to a Low-Protein Diet Regulates Intestinal Expression of Amino Acid and Peptide Transporters in Weanling Pigs. Amino Acids 2013, 45, 1191–1205. [Google Scholar] [CrossRef]
- Reyer, H.; Zentek, J.; Männer, K.; Youssef, I.M.; Aumiller, T.; Weghuber, J.; Wimmers, K.; Mueller, A.S. Possible Molecular Mechanisms by Which an Essential Oil Blend from Star Anise, Rosemary, Thyme, and Oregano and Saponins Increase the Performance and Ileal Protein Digestibility of Growing Broilers. J. Agric. Food Chem. 2017, 65, 6821–6830. [Google Scholar] [CrossRef]
- Omar, J.A.; Hejazi, A.; Badran, R. Performance of broilers supplemented with natural herb extract. J. Anim. Sci. 2016, 6, 68–74. [Google Scholar] [CrossRef] [Green Version]
- National Research Council. Nutrient Requirements of Swine: Eleventh Revised Edition; The National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Shili, C.N.; Broomhead, J.N.; Spring, S.C.; Lanahan, M.B.; Pezeshki, A. A Novel Corn-Expressed Phytase Improves Daily Weight Gain, Protein Efficiency Ratio and Nutrients Digestibility and Alters Fecal Microbiota in Pigs Fed with Very Low Protein Diets. Animals 2020, 10, 1926. [Google Scholar] [CrossRef]
- Horwitz, W. Official Methods of Analysis of AOAC International. Volume I, Agricultural Chemicals, Contaminants, Drugs/Edited by William Horwitz; AOAC International: Gaithersburg, MD, USA, 2010. [Google Scholar]
- Cooper, C.; Packer, N.; Williams, K. Amino Acid Analysis Protocols; Springer Science & Business Media: Totowa, NJ, USA, 2001; Volume 159. [Google Scholar]
- Pezeshki, A.; Muench, G.P.; Chelikani, P.K. Short Communication: Expression of Peptide YY, Proglucagon, Neuropeptide Y Receptor Y2, and Glucagon-like Peptide-1 Receptor in Bovine Peripheral Tissues. J. Dairy Sci. 2012, 95, 5089–5094. [Google Scholar] [CrossRef] [PubMed]
- Thongsong, B.; Wiyaporn, M.; Kalandakanond-Thongsong, S. Blood Glucose, Amino Acid Profiles and Nutrient Transporter Gene Expressions in the Small Intestine of Low and Normal Birthweight Piglets during the Early Suckling Period. Vet. J. 2019, 247, 1–7. [Google Scholar] [CrossRef]
- Wang, H.; Ji, Y.; Wu, G.; Sun, K.; Sun, Y.; Li, W.; Wang, B.; He, B.; Zhang, Q.; Dai, Z.; et al. L-Tryptophan Activates Mammalian Target of Rapamycin and Enhances Expression of Tight Junction Proteins in Intestinal Porcine Epithelial Cells. J. Nutr. 2015, 145, 1156–1162. [Google Scholar] [CrossRef] [Green Version]
- Vigors, S.; Sweeney, T.; O’Shea, C.J.; Browne, J.A.; O’Doherty, J.V. Improvements in Growth Performance, Bone Mineral Status and Nutrient Digestibility in Pigs Following the Dietary Inclusion of Phytase Are Accompanied by Modifications in Intestinal Nutrient Transporter Gene Expression. Br. J. Nutr. 2014, 112, 688–697. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.; Liu, M.; Ren, W.; Duan, J.; Yang, G.; Zhao, Y.; Fang, R.; Chen, L.; Li, T.; Yin, Y. Effects of Dietary Supplementation with Glutamate and Aspartate on Diquat-Induced Oxidative Stress in Piglets. PLoS ONE 2015, 10, e0122893. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative CT Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Safdar, K.; Muhammad, K.; Naz, R. Effect of Herbal Extracts on Serum Minerals, Lipid Profile and Anti-NDV-HI Antibody Levels of Vaccinated Broiler Chicks. Pak. J. Zool. 2016, 48, 1715–1719. [Google Scholar]
- Schenck, P.A.; Chew, D.J.; Nagode, L.A.; Rosol, T. Disorders of Calcium: Hypercalcemia and Hypocalcemia. Fluid Electrolyte Acid Base Disord. Small Anim. Pract. 2006, 4, 122–194. [Google Scholar] [CrossRef]
- Hagemoser, W.A.; Goff, J.P.; Sanderson, T.P.; Haynes, J.S. Osteopenic Disease in Growing Pigs: Diagnostic Methods Using Serum and Urine Calcium and Phosphorus Values, Parathormone Assay, and Bone Analysis. J. Vet. Diagn. Investig. 2000, 12, 524–534. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-J.; Kim, J.-H.; Kim, J.-W.; Kim, S.-H.; Eun, J.-S.; Hong, S.-M. PSVIII-17 In Vitro and in Vivo Investigations of Effects of Natural Phytogenic Compounds on Muscle Cell Gene Expressions and Growth Performance and Carcass Composition of Finishing Pigs. J. Anim. Sci. 2019, 97 (Suppl. 3), 303–304. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Dai, Z.; Li, D.; Wang, J.; Wu, Z. Amino Acid Nutrition in Animals: Protein Synthesis and Beyond. Annu. Rev. Anim. Biosci. 2014, 2, 387–417. [Google Scholar] [CrossRef]
- Kong, X.; Wu, G.; Yin, Y. Roles of Phytochemicals in Amino Acid Nutrition. Front. Biosci. 2011, 3, 372–384. [Google Scholar]
- Lan, R.X.; Li, T.S.; Kim, I.H. Effects of Essential Oils Supplementation in Different Nutrient Densities on Growth Performance, Nutrient Digestibility, Blood Characteristics and Fecal Microbial Shedding in Weaning Pigs. Anim. Feed Sci. Technol. 2016, 214, 77–85. [Google Scholar] [CrossRef]
- Kanai, Y.; Hediger, M.A. The Glutamate and Neutral Amino Acid Transporter Family: Physiological and Pharmacological Implications. Eur. J. Pharmacol. 2003, 479, 237–247. [Google Scholar] [CrossRef]
- Burckhardt, B.C.; Burckhardt, G. Interaction of Excitatory Amino Acid Transporters 1–3 (EAAT1, EAAT2, EAAT3) with N-Carbamoylglutamate and N-Acetylglutamate. Cell. Physiol. Biochem. 2017, 43, 1907–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.C.; Donaldson, P.J. Focus on Molecules: The Cystine/Glutamate Exchanger (System x(c)(−)). Exp. Eye Res. 2011, 92, 162–163. [Google Scholar] [CrossRef] [PubMed]
- Bröer, A.; Klingel, K.; Kowalczuk, S.; Rasko, J.E.J.; Cavanaugh, J.; Bröer, S. Molecular Cloning of Mouse Amino Acid Transport System B0, a Neutral Amino Acid Transporter Related to Hartnup Disorder. J. Biol. Chem. 2004, 279, 24467–24476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Ren, W.; Duan, J.; Wu, L.; Chen, S.; Li, T.; Yin, Y.; Wu, G. Dietary Arginine Supplementation Enhances Intestinal Expression of SLC7A7 and SLC7A1 and Ameliorates Growth Depression in Mycotoxin-Challenged Pigs. Amino Acids 2014, 46, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Drummond, M.J.; Fry, C.S.; Glynn, E.L.; Timmerman, K.L.; Dickinson, J.M.; Walker, D.K.; Gundermann, D.M.; Volpi, E.; Rasmussen, B.B. Skeletal Muscle Amino Acid Transporter Expression Is Increased in Young and Older Adults Following Resistance Exercise. J. Appl. Physiol. 2011, 111, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Hyde, R.; Taylor, P.M.; Hundal, H.S. Amino Acid Transporters: Roles in Amino Acid Sensing and Signalling in Animal Cells. Biochem. J. 2003, 373, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Verrey, F.; Closs, E.I.; Wagner, C.A.; Palacin, M.; Endou, H.; Kanai, Y. CATs and HATs: The SLC7 Family of Amino Acid Transporters. Pflug. Arch. 2004, 447, 532–542. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diets 1 | |||||
|---|---|---|---|---|---|
| N1 | N2 | N3 | |||
| CON | LP | CON | LP | ||
| Ingredients, % | |||||
| Corn, yellow dent 2 | 32.21 | 45.04 | 64.72 | 57.06 | 73.49 |
| Soybean meal, 47.5% CP 2 | 15.00 | 36.22 | 13.94 | 38.39 | 19.95 |
| Fish meal, menhaden 2 | 6.00 | 4.90 | 4.98 | 2.07 | 2.11 |
| Whey, dried 2 | 25.00 | 5.87 | 5.97 | - | - |
| Lactose 2 | 7.00 | - | - | - | - |
| Corn starch 2 | - | 5.87 | 5.97 | - | - |
| Plasma spray-dried 2 | 6.00 | - | - | - | - |
| Soy protein concentrate 2 | 2.20 | - | - | - | - |
| Soybean oil 2 | 4.00 | - | - | - | - |
| Dicalcium phosphate 2 | 0.67 | 0.76 | 0.85 | 0.99 | 1.15 |
| Limestone 2 | 0.45 | 0.54 | 0.67 | 0.62 | 0.68 |
| Nursery vitamin premix 3 | 0.05 | 0.19 | 0.19 | 0.18 | 0.20 |
| Salt 2 | 0.50 | 0.49 | 0.50 | 0.57 | 0.52 |
| Chromium oxide 2 | - | 0.50 | 0.50 | 0.50 | 0.52 |
| Trace mineral premix 4 | 0.06 | 0.07 | 0.06 | 0.07 | 0.08 |
| Selplex 2 | 0.05 | - | - | - | - |
| Choline Cl 2 | 0.03 | - | - | - | - |
| Zinc oxide, 72% Zn 2 | 0.35 | - | - | - | - |
| L-Lysine, sulfate 2 | 0.17 | - | 1.12 | - | 0.88 |
| DL-Methionine 2 | 0.18 | - | 0.14 | - | 0.08 |
| L-Threonine 2 | 0.07 | - | 0.28 | - | 0.22 |
| L-Tryptophan 2 | - | - | 0.13 | - | 0.10 |
| Calculated Chemical Composition 5 | |||||
| Dry matter, % | 92.34 | 90.72 | 90.72 | 89.66 | 89.60 |
| ME, Mcal/kg | 3.52 | 3.40 | 3.40 | 3.30 | 3.32 |
| Crude protein, % | 22.99 | 24.71 | 17.03 | 24.26 | 17.83 |
| Crude fiber, % | 1.39 | 2.31 | 2.01 | 2.64 | 2.39 |
| Crude fat, % | 6.68 | 3.36 | 3.46 | 3.57 | 3.66 |
| Nitrogen, % | 3.65 | 3.95 | 2.72 | 3.88 | 2.85 |
| Calcium, % | 0.89 | 0.80 | 0.80 | 0.70 | 0.70 |
| Phosphorous, % | 0.79 | 0.71 | 0.63 | 0.68 | 0.62 |
| Available phosphorous, % | 0.59 | 0.40 | 0.40 | 0.32 | 0.33 |
| SID Lysine, % | 1.54 | 1.35 | 1.35 | 1.24 | 1.23 |
| SID Threonine, % | 0.97 | 0.84 | 0.80 | 0.80 | 0.76 |
| SID Methionine, % | 0.51 | 0.38 | 0.41 | 0.36 | 0.35 |
| SID Tryptophan, % | 0.27 | 0.27 | 0.28 | 0.27 | 0.27 |
| Analyzed Chemical Composition 6 | |||||
| Dry matter, % | 90.10 | 89.00 | 87.00 | 91.30 | 87.40 |
| Crude protein, % | 22.70 | 24.70 | 17.30 | 23.00 | 17.70 |
| Carbohydrate, % kcal | 45.33 | 70.21 | 79.98 | 77.79 | 86.10 |
| Crude fiber, % | 1.30 | 2.00 | 1.60 | 3.50 | 2.10 |
| Calcium, % | 0.85 | 0.81 | 0.82 | 0.80 | 0.85 |
| Phosphorus, % | 0.75 | 0.65 | 0.56 | 0.70 | 0.61 |
| Nitrogen, % | 3.60 | 3.90 | 2.80 | 3.68 | 2.80 |
| Genes 1 | Sequence (5′ → 3′) | Location on Template | Amplicon Length (bp) | GenBank Accession Number |
|---|---|---|---|---|
| SLC15A1 | F: AGCATCTTCTTCATCGTGGTCAA R: GTCTTGAACTTCCCCAGCCA | 43–65 229–248 | 206 | NM_214347.1 |
| SLC7A9 | F: ATCGGTCTGGCGTTTTAT R: GGATCTAGCACCCTGTCA | 816–833 943–960 | 145 | XM_021093176.1 |
| SLC7A8 | F: TTTCCAGGAACCTGACATCG R: ACATTGCAGTGACATAAGCG | 576–595 756–775 | 200 | XM_003128550.6 |
| SLC6A19 | F: CACAACAACTGCGAGAAGGA R: CCGTTGATAAGCGTCAGGAT | 1101–1120 1236–1255 | 155 | XM_003359855.4 |
| SLC3A1 | F: TTTCCGCAATCCTGATGTTC R: GGGTCTTATTCACTTGGGTC | 1107–1126 1233–1252 | 146 | NM_001123042.1 |
| SLC7A11 | F: CGGCTCCTGGGAAATTTCTC R: ACCATTCATGGAGCCAAAGC | 1297–1316 1349–1368 | 72 | XM_021101587.1 |
| SLC7A1 | F: TCTCATCCTAACGGGACTTTTAACTC R: GACCAGAACGTTGATACACGTGAA | 2525–2550 2586–2609 | 85 | XM_021065165.1 |
| β-Actin | F: CTGCGGCATCCACGAAACT R: AGGGCCGTGATCTCCTTCTG | 944–962 1071–1090 | 147 | XM_021086047.1 |
| Diets 1 | SEM 2 | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Parameters | CON-NS | CON-LS | CON-HS | LP-NS | LP-LS | LP-HS | Protein | PWA 3 | Protein × PWA | |
| Initial BW 4, kg | 8.82 | 9.10 | 9.63 | 9.05 | 8.76 | 8.76 | 0.17 | 0.36 | 0.78 | 0.46 |
| Final BW 4, kg | 25.34 * | 26.08 | 29.09 * | 24.89 | 25.00 | 23.25 | 0.56 | 0.02 | 0.72 | 0.09 |
| ADG 4, kg/d | 0.59 | 0.60 | 0.70 b | 0.57 | 0.58 | 0.52 | 0.01 | 0.01 | 0.75 | 0.07 |
| ADFI 4, kg/d | 0.86 | 0.95 | 0.97 | 0.84 | 0.91 | 0.86 | 0.02 | 0.23 | 0.30 | 0.69 |
| ADWI 4, L/d | 3.09 | 2.98 | 4.17 | 2.78 | 3.54 d | 2.98 | 0.17 | 0.35 | 0.31 | 0.12 |
| G:F 4, kg/kg | 0.69 | 0.63 | 0.72 | 0.67 | 0.62 | 0.61 | 0.01 | 0.01 | 0.97 | 0.72 |
| G:P 4, kg/kg | 2.97 | 2.83 | 3.09 | 3.83 c | 3.58 | 3.38 | 0.09 | 0.01 | 0.33 | 0.12 |
| Muscle lean% | 82.83 | 83.35 | 86.00 | 78.58 | 80.32 | 88.7 e | 0.86 | 0.01 | 0.01 | 0.01 |
| Muscle fat% | 16.62 | 16.06 | 13.51 | 20.85 | 13.33 | 9.41 e | 0.82 | 0.01 | 0.01 | 0.01 |
| Diets 1 | SEM 2 | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Parameters | CON-NS | CON-LS | CON-HS | LP-NS | LP-LS | LP-HS | Protein | PWA 3 | Protein × PWA | |
| BWG 4, kg | ||||||||||
| Wk 1 | 1.98 | 2.32 | 2.97 | 1.36 | 1.24 | 2.77 | 0.12 | 0.04 | 0.47 | 0.18 |
| Wk 2 | 4.05 | 4.24 | 4.20 | 3.33 | 4.13 | 3.22 | 0.13 | 0.03 | 0.19 | 0.41 |
| Wk 3 | 5.01 | 4.80 | 4.91 | 4.96 | 5.09 | 3.90 | 0.18 | 0.39 | 0.47 | 0.20 |
| Wk 4 | 5.48 | 5.88 | 7.37 b | 6.22 | 5.77 | 4.75 | 0.21 | 0.14 | 0.77 | 0.03 |
| CFI 4, kg | ||||||||||
| Wk1 | 3.35 | 3.63 | 3.76 | 2.97 | 3.10 | 3.82 | 0.14 | 0.32 | 0.19 | 0.69 |
| Wk 2 | 5.11 | 5.83 | 6.17 | 4.61 | 5.33 | 5.80 | 0.19 | 0.21 | 0.04 | 0.98 |
| Wk 3 | 7.30 | 7.72 | 7.38 | 6.79 | 7.83 | 6.85 | 0.22 | 0.49 | 0.34 | 0.80 |
| Wk 4 | 8.28 | 8.92 | 9.67 | 9.33 | 9.49 | 7.86 | 0.25 | 0.89 | 0.72 | 0.05 |
| CPI 4, kg | ||||||||||
| Wk1 | 0.83 | 0.90 | 0.93 | 0.51 | 0.54 | 0.66 | 0.03 | 0.01 | 0.26 | 0.84 |
| Wk 2 | 1.17 *# | 1.34 | 1.41 # | 0.81 *$ | 0.94 | 1.02 $ | 0.04 | 0.01 | 0.04 | 0.97 |
| Wk 3 | 1.67 * | 1.77 | 1.69 | 1.20 * | 1.38 | 1.21 | 0.05 | 0.01 | 0.38 | 0.89 |
| Wk 4 | 1.89 | 2.18 | 2.25 | 1.62 | 1.64 | 1.33 | 0.06 | 0.01 | 0.42 | 0.03 |
| CWI 4, L | ||||||||||
| Wk 1 | 12.31 | 15.95 | 14.45 | 12.42 | 10.90 | 13.95 | 0.92 | 0.34 | 0.73 | 0.46 |
| Wk 2 | 24.19 | 23.18 | 25.02 | 12.54 c | 19.52 | 22.93 e | 1.50 | 0.05 | 0.30 | 0.36 |
| Wk 3 | 25.51 | 22.90 | 30.71 | 26.09 | 29.98 | 23.65 | 1.55 | 0.96 | 0.93 | 0.20 |
| Wk 4 | 30.04 | 26.43 | 34.94 | 26.13 * | 38.39 * | 24.72 | 1.85 | 0.81 | 0.63 | 0.04 |
| G:F 4, kg/kg | ||||||||||
| Wk 1 | 0.59 | 0.64 | 0.79 | 0.46 | 0.40 | 0.73 | 0.05 | 0.01 | 0.03 | 0.60 |
| Wk 2 | 0.79 | 0.72 | 0.68 | 0.73 | 0.77 | 0.57 | 0.02 | 0.55 | 0.01 | 0.31 |
| Wk 3 | 0.69 | 0.62 | 0.66 | 0.73 | 0.66 | 0.57 | 0.02 | 0.54 | 0.13 | 0.15 |
| Wk 4 | 0.66 | 0.63 | 0.76 | 0.67 | 0.61 | 0.60 | 0.01 | 0.16 | 0.54 | 0.38 |
| G:P 4, kg/kg | ||||||||||
| Wk 1 | 2.39 | 2.58 | 3.19 | 2.67 | 2.30 | 4.20 e | 0.20 | 0.94 | 0.03 | 0.14 |
| Wk 2 | 3.46 | 3.16 | 2.98 | 4.11 | 4.39 | 3.16 | 0.10 | 0.02 | 0.01 | 0.22 |
| Wk 3 | 3.00 | 2.71 | 2.91 | 4.13 c | 3.69 | 3.22 e | 0.09 | 0.01 | 0.06 | 0.16 |
| Wk 4 | 2.90 | 2.70 | 3.28 | 3.84 c | 3.52 | 3.57 | 0.07 | 0.01 | 0.56 | 0.27 |
| Item | Diets 1 | SEM 2 | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CON-NS | CON-LS | CON-HS | LP-NS | LP-LS | LP-HS | Protein | PWA | Protein × PWA | ||
| Valine | 343.7 | 372.7 | 518.0 b | 130.1 c | 246.6 d | 86.5 | 26.3 | 0.01 | 0.01 | 0.01 |
| Methionine | 41.2 * | 59.8 * | 68.6 | 64.5 | 72.3 | 73.0 | 3.2 | 0.02 | 0.04 | 0.41 |
| Threonine | 328.3 $* | 388.4 | 460.0 $ | 631.5 * | 660.2 | 590.7 | 28.9 | 0.01 | 0.10 | 0.26 |
| Isoleucine | 171.9 | 190.6 | 301.3 b | 100.4 c# | 121.7 # | 75.9 | 13.4 | 0.01 | 0.01 | 0.01 |
| Leucine | 268.9 | 314.1 a | 428.6 b | 255.9 # | 294.7 # | 214.3 | 12.9 | 0.01 | 0.01 | 0.01 |
| Phenylalanine | 126.4 * | 146.2 | 194.7 b | 100.8 *# | 118.8 # | 89.0 | 6.6 | 0.01 | 0.01 | 0.01 |
| Tryptophan | 55.8 | 70.4 | 109.4 b | 81.2 | 93.7 | 112.8 | 4.9 | 0.02 | 0.01 | 0.42 |
| Lysine | 189.6 * | 267.6 * | 353.6 b | 529.0 c* | 618.3 | 365.7 * | 29.0 | 0.01 | 0.09 | 0.01 |
| Histidine | 136.3 | 137.3 | 182.2 b | 74.2 c | 95.3 d | 50.4 e | 8.0 | 0.01 | 0.28 | 0.01 |
| Arginine | 39.7 | 105.2 | 393.8 b | 63.8 | 79.4 | 124.6 | 11.9 | 0.01 | 0.01 | 0.01 |
| Aspartic acid | 79.2 | 76.7 | 41.67 b | 58.3 | 78.6 | 30.17 | 4.6 | 0.16 | 0.01 | 0.42 |
| Serine | 307.4 | 382.4 a | 401.3 b | 395.7 c | 420.5 | 285.6 e | 11.9 | 0.84 | 0.02 | 0.01 |
| Glutamic acid | 444.0 | 485.8 | 261.6 b | 499.9 | 594.9 | 330.3 e | 24.9 | 0.03 | 0.01 | 0.81 |
| Glutamine | 632.8 | 683.9 | 840.5 | 770.4 | 892.4 | 862.8 | 29.9 | 0.06 | 0.05 | 0.64 |
| Glycine | 1420.1 | 1772.4 a | 1701.3 b | 2168.0 c | 2027.7 | 1887.3 | 54.5 | 0.01 | 0.45 | 0.01 |
| Alanine | 778.4 | 917.4 a | 989.1 b | 1054.4 c | 1262.6 d | 915.5 | 34.0 | 0.01 | 0.01 | 0.01 |
| Tyrosine | 167.3 | 199.6 | 305.1 b | 159.2 | 181.9 | 129.9 | 10.5 | 0.01 | 0.03 | 0.01 |
| Asparagine | 169.3 | 208.7 | 291.9 b | 164.0 | 181.4 | 150.1 | 11.9 | 0.01 | 0.10 | 0.02 |
| Proline | 457.2 | 528.6 a | 676.9 b | 505.6 # | 538.3 | 396.8 # | 17.6 | 0.01 | 0.07 | 0.01 |
| Ammonia | 367.3 | 452.7 | 245.3 | 403.1 | 424.0 | 232.1 e | 19.1 | 0.62 | 0.01 | 0.25 |
| Creatinine | 115.6 | 131.1 | 161.7 | 112.9 | 129.8 | 81.7 | 6.2 | 0.01 | 0.43 | 0.01 |
| Taurine | 266.7 | 281.0 | 141.4 b | 273.8 | 357.3 | 96.4 e | 19.1 | 0.27 | 0.01 | 0.01 |
| Sarcosine | 50.7 | 66.1 a | 62.7 b | 73.6 c | 80.9 | 74.0 | 2.2 | 0.01 | 0.02 | 0.31 |
| 3,methylhistidine | 10.9 | 10.2 | 12.5 | 6.71 c | 7.6 | 6.8 | 0.4 | 0.01 | 0.42 | 0.08 |
| 1,methylhistidine | 58.4 | 70.3 | 67.3 | 56.7 | 57.9 | 60.2 | 3.0 | 0.26 | 0.63 | 0.78 |
| Ethanolamine | 38.1 | 47.1 | 25.0 | 29.4 | 20.3 | 17.9 | 3.0 | 0.01 | 0.01 | 0.26 |
| Carnosine | 31.9 | 29.7 | 30.4 | 22.7 | 23.1 | 19.2 | 1.7 | 0.01 | 0.81 | 0.84 |
| Hydroxylysine | 11.5 | 14.6 | 14.1 | 9.6 | 9.8 | 12.5 | 0.5 | 0.01 | 0.03 | 0.24 |
| Citrulline | 74.5 | 89.6 | 96.4 | 70.1 | 75.8 | 79.1 | 2.7 | 0.02 | 0.04 | 0.53 |
| Ornithine | 352.8 $ | 360.5 | 285.5 $ | 251.5 # | 311.4 # | 125.2 | 17.2 | 0.01 | 0.01 | 0.16 |
| α-aminobutyric acid | 35.6 | 41.0 | 13.4 | 39.6 | 34.0 | 17.7 | 3.1 | 0.93 | 0.01 | 0.63 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shili, C.N.; Habibi, M.; Sutton, J.; Barnes, J.; Burch-Konda, J.; Pezeshki, A. Effect of a Phytogenic Water Additive on Growth Performance, Blood Metabolites and Gene Expression of Amino Acid Transporters in Nursery Pigs Fed with Low-Protein/High-Carbohydrate Diets. Animals 2021, 11, 555. https://doi.org/10.3390/ani11020555
Shili CN, Habibi M, Sutton J, Barnes J, Burch-Konda J, Pezeshki A. Effect of a Phytogenic Water Additive on Growth Performance, Blood Metabolites and Gene Expression of Amino Acid Transporters in Nursery Pigs Fed with Low-Protein/High-Carbohydrate Diets. Animals. 2021; 11(2):555. https://doi.org/10.3390/ani11020555
Chicago/Turabian StyleShili, Cedrick N., Mohammad Habibi, Julia Sutton, Jessie Barnes, Jacob Burch-Konda, and Adel Pezeshki. 2021. "Effect of a Phytogenic Water Additive on Growth Performance, Blood Metabolites and Gene Expression of Amino Acid Transporters in Nursery Pigs Fed with Low-Protein/High-Carbohydrate Diets" Animals 11, no. 2: 555. https://doi.org/10.3390/ani11020555