
Arginine Alters miRNA Expression Involved in Development and Proliferation of Rat Mammary Tissue

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Design
Verification of miRNA and Target Genes Expression by qPCR
2.2. Collection of Rat Mammary Gland Tissue
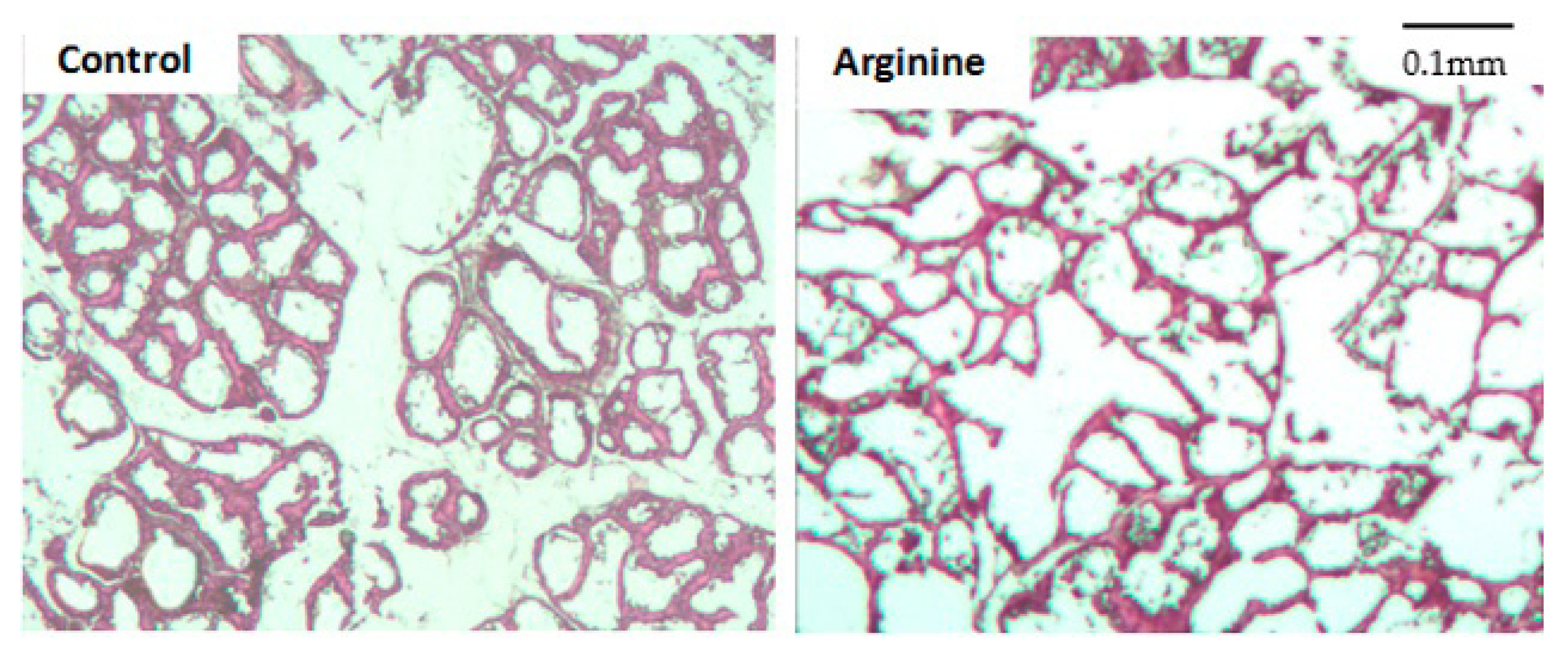
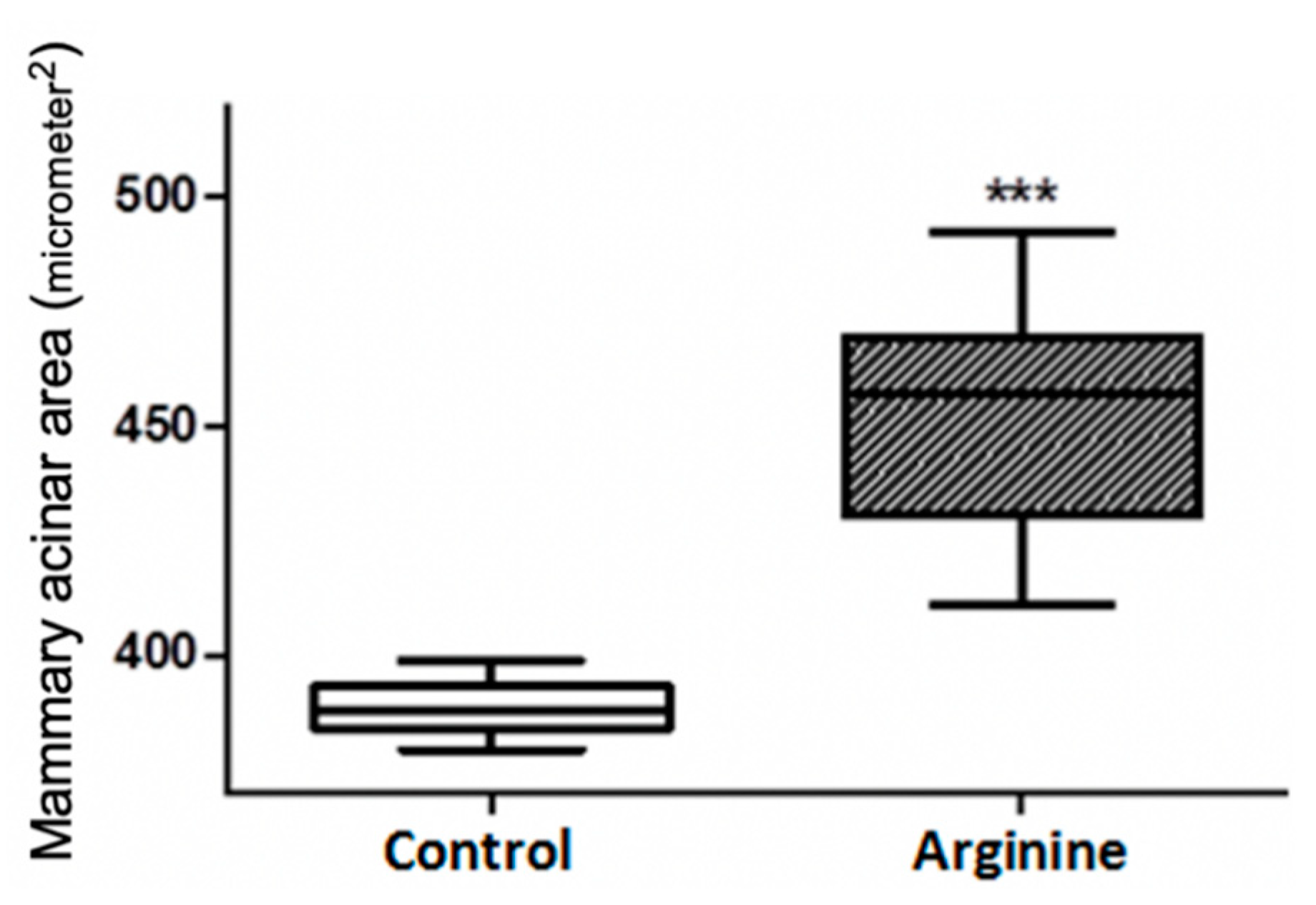
2.3. Mammary Histological Section Preparation and Acinar Area Measurement
2.4. Mammary RNA Extraction and Sequencing Library Construction
2.5. Cluster Generation and High-Throughput Sequencing Analysis
2.6. Data Comparison and Classification
2.7. Saturation Analysis
2.8. Differential Expression Analysis of miRNAs and Target Gene Prediction
2.9. Novel miRNA Prediction
2.10. High Throughput Sequencing Analysis
2.10.1. RNA Quality Detection
2.10.2. Library Quality Control
2.10.3. Saturation Analysis
3. Results
3.1. Effect on the Development of Mammary Tissue
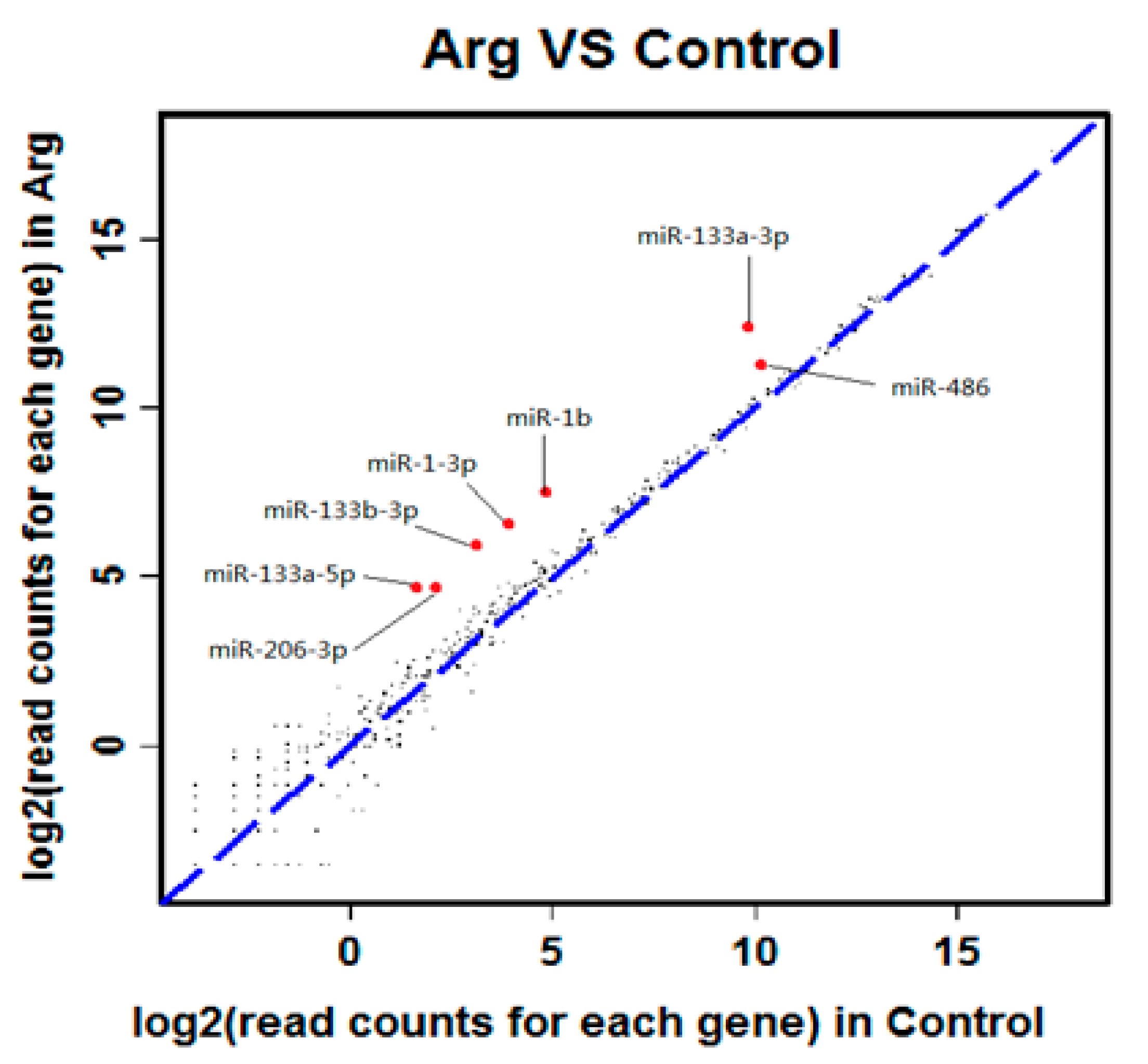
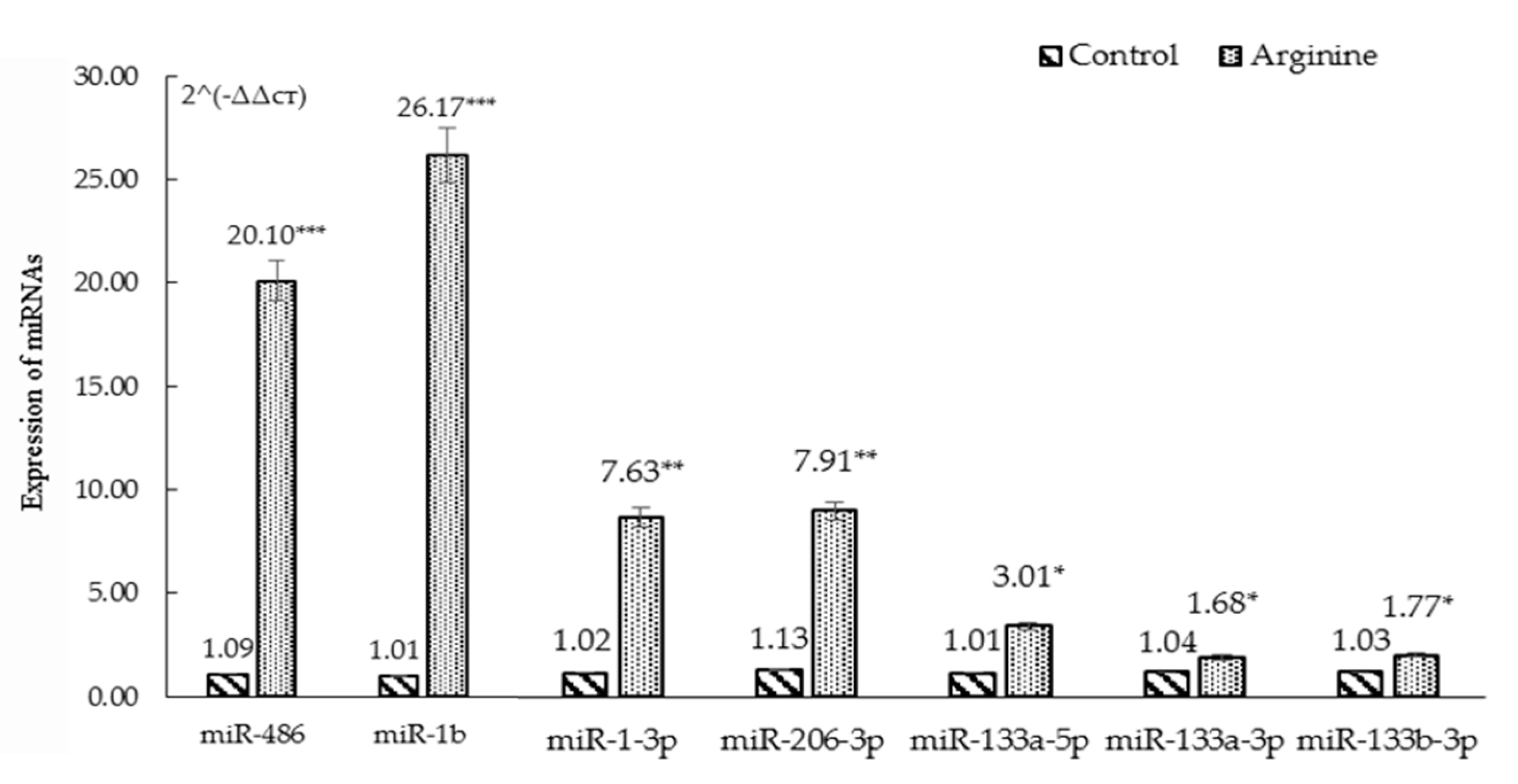
3.2. High Throughput Sequencing Analysis and RT-PCR Verification
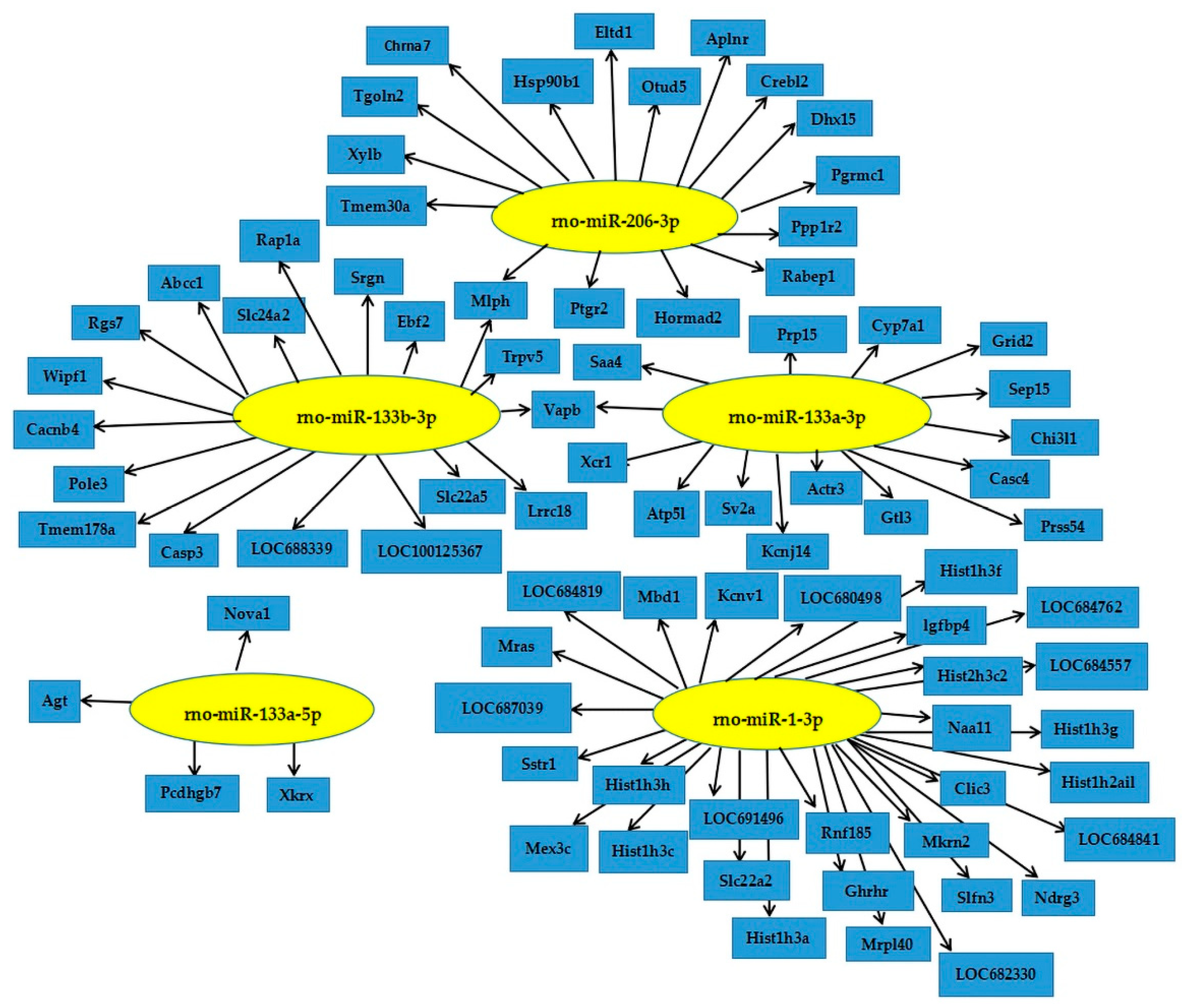
3.3. Prediction and Enrichment Analysis of Target Genes of Differentially Expressed miRNA
3.4. Target Gene Prediction of Differentially Expressed miRNAs and RT-PCR Validation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prizant, R.L.; Barash, I. Negative effects of the amino acids Lys, His, and Thr on S6K1 phosphorylation in mammary epithelial cells. J. Cell Biochem. 2008, 105, 1038–1047. [Google Scholar] [CrossRef]
- Kim, S.W.; Wu, G. Regulatory role for amino acids in mammary gland growth and milk synthesis. Amino Acids 2009, 37, 89–95. [Google Scholar] [CrossRef]
- Cant, J.P.; DePeters, E.J.; Baldwin, R.L. Mammary up-take of energy metabolites in dairy cows fed fat and its relationship to milk protein depression. J. Dairy Sci. 1993, 76, 2254–2265. [Google Scholar] [CrossRef]
- Toerien, C.A.; Trout, D.R.; Cant, J.P. Nutritional stimulation of milk protein yield of cows is associated with changes in phosphorylation of mammary eukaryotic initiation factor 2 and ribosomal s6 kinase. J. Nutr. 2010, 140, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Ehmann, V.L.; Peterson, W.D. To grow mouse mammary epithelial cell in culture. J. Cell Biol. 1984, 98, 1026–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wang, Y.; Wang, M.; Zhou, G.; Chen, L.; Ding, L.; Bu, D.; Loor, J. Arginine Supply Impacts the Expression of Candidate microRNA Controlling Milk Casein Yield in Bovine Mammary Tissue. Animals 2020, 10, 797. [Google Scholar] [CrossRef]
- Wu, G.; Meininger, C.J. Regulation of nitric oxide synthesis by dietary factors. Annu. Rev. Nutr. 2002, 22, 61–86. [Google Scholar] [CrossRef]
- Wu, G.; Morris, S.M., Jr. Arginine metabolism: Nitric oxide and beyond. Biochem. J. 1998, 336, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Mabjeesh, S.J.; Kyle, C.E.; Macrae, J.C.; Hanigan, M.D.; Bequette, B.J. Vascular Sources of Amino Acids for Milk Protein Synthesis in Goats at Two Stages of Lactation. J. Dairy Sci. 2002, 85, 919–929. [Google Scholar] [CrossRef]
- Chen, L.M.; Li, Z.T.; Wang, M.Z.; Wang, H.R. Preliminary report of arginine on synthesis and gene expression of casein in bovine mammary epithelial cell. IRJAS 2013, 4, 17–23. [Google Scholar]
- Wang, M.; Xu, B.; Wang, H.; Bu, D.; Wang, J.; Loor, J.J. Effects of Arginine concentration on the in vitro expression of Casein and mTOR pathway related genes in mammary epithelial cells from dairy cattle. PLoS ONE 2014, 9, e95985. [Google Scholar] [CrossRef]
- Wang, M.Z.; Moisá, M.S.; Khan, M.J.; Wang, J.; Bu, D.; Loor, J.J. MicroRNA expression patterns in the bovine mammary gland are affected by stage of lactation. J. Dairy Sci. 2012, 95, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muroya, S.; Hagi, T.; Kimura, A.; Aso, H.; Matsuzaki, M.; Nomura, M. Lactogenic hormones alter cellular and extracellular microRNA expression in bovine mammary epithelial cell culture. J. Anim. Sci. Biotechnol. 2016, 4, 504–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, L.; Carbone, E.T.; Yi, E.M.; Bauer, D.B.; Lindstrom, K.A.; Parker, J.M.; Austin, J.A.; Seo, Y.; Gandhi, A.D.; Wilkerson, J.D. Assessing cervical dislocation as a humane euthanasia method in mice. J. Am. Assoc. Lab Anim. Sci. 2012, 51, 352–356. [Google Scholar] [PubMed]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C. Ain-93 purified diets for laboratory rodents: Final report of the American institute of nutrition ad hoc writing committee on the reformulation of the ain-76a rodent diet. J. Nutr. 1993, 123, 1939. [Google Scholar] [CrossRef] [PubMed]
- Mestdagh, P.; Hartmann, N.; Baeriswyl, L.; Andreasen, D.; Bernard, N.; Chen, C.; Cheo, D.; D’Andrade, P.; DeMayo, M.; Dennis, L.; et al. Corrigendum: Evaluation of quantitative miRNA expression platforms in the microRNA quality control (miRQC) study. Nat. Methods 2014, 11, 971. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Liu, S.; Wang, H.; Wang, M.; Yu, L. Relative significances of pH and substrate starch level to roles of Streptococcus bovis S1 in rumen acidosis. AMB Express 2016, 6, 80. [Google Scholar] [CrossRef] [Green Version]
- Shackleton, M.; Vaillant, F.; Simpson, K.J.; Stingl, J.; Smyth, G.K.; Asselin-Labat, M.L.; Wu, L.; Lindeman, G.J.; Visvader, J.E. Generation of a functional mammary gland from a single stem cell. Nature 2006, 439, 84–88. [Google Scholar] [CrossRef]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef] [Green Version]
- Aken, B.L.; Ayling, S.; Barrell, D.; Clarke, L.; Curwen, V.; Fairley, S.; Fernandez Banet, J.; Billis, K.; García Girón, C.; Hourlier, T.; et al. The Ensembl gene annotation system. Database J. Biol. Databases Curation 2016, baw093. [Google Scholar] [CrossRef]
- Sai Lakshmi, S.; Agrawal, S. piRNABank: A web resource on classified and clustered Piwi-interacting RNAs. Nucleic Acids Res. 2008, 36, D173–D177. [Google Scholar] [CrossRef]
- Kalvari, I.; Nawrocki, E.P.; Argasinska, J.; Quinones-Olvera, N.; Finn, R.D.; Bateman, A.; Petrov, A.I. Non-Coding RNA Analysis Using the Rfam Database. Curr. Protoc. Bioinform. 2018, 62, e51. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, R.C.; Krajewska, M.; Krnacik, S.; Jaeger, R.; Weiher, H.; Krajewski, S. Apoptosis in the terminal endbud of the murine mammary gland: A mechanism of ductal morphogenesis. Development 1996, 122, 4013–4022. [Google Scholar]
- Bequette, B.J.; Backwell, F.R. Amino acid supply and metabolism by the ruminant mammary gland. Proc. Nutr. Soc. 1997, 56, 593–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapierre, H.; Lobley, G.E.; Doepel, L.; Raggio, G.; Rulquin, H.; Lemosquet, S. Triennial lactation symposium: Mammary metabolism of amino acids in dairy cows. J. Anim. Sci. 2012, 90, 1708–1721. [Google Scholar] [CrossRef]
- Mezl, V.A.; Knox, W.E. Metabolism of arginine in lactating rat mammary gland. Biochem. J. 1977, 166, 105–113. [Google Scholar] [CrossRef] [Green Version]
- O’Quinn, P.R.; Knabe, D.A.; Wu, G. Arginine catabolism in lactating porcine mammary tissue. J. Anim. Sci. 2002, 80, 467. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Davis, T.A.; Kim, S.W.; Li, P.; Marc, R.J. Arginine metabolism and nutrition in growth, health and disease. Amino Acids 2009, 37, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Mateo, R.D.; Wu, G.; Moon, H.K.; Carroll, J.A.; Kim, S.W. Effects of dietary arginine supplementation during gestation and lactation on the performance of lactating primiparous sows and nursing piglets. J. Anim. Sci. 2008, 86, 827. [Google Scholar] [CrossRef]
- Song, L.; Tuan, R.S. MicroRNAs and cell differentiation in mammalian development. Birth Defects Res. C Embryo. Today 2006, 78, 140–149. [Google Scholar] [CrossRef]
- Wang, C.; Li, Q. Identification of differentially expressed microRNAs during the development of Chinese murine mammary gland. J. Genet. Genom. 2007, 34, 966–973. [Google Scholar] [CrossRef]
- Avril-Sassen, S.; Goldstein, L.D.; Stingl, J.; Blenkiron, C.; Le Quesne, J.; Spiteri, I.; Karagavriilidou, K.; Watson, C.J.; Tavaré, S.; Miska, E.A.; et al. Characterization of microRNA expression in post-natal mouse mammary gland development. BMC Genom. 2009, 10, 548. [Google Scholar] [CrossRef] [Green Version]
- Lagos-quintana, M.; Rauhut, R.; Yalcin, A.; Meyer, J.; Lendeckel, W.; Tuschl, T. Identification of tissue-specific microRNAs from mouse. Curr. Biol. 2002, 12, 735–739. [Google Scholar] [CrossRef] [Green Version]
- Mccarthy, J.J. The myomir network in skeletal muscle plasticity. Exerc. Sport Sci. Rev. 2011, 39, 150. [Google Scholar] [CrossRef]
- Liu, N.; Olson, E.N. MicroRNA regulatory networks in cardiovascular development. Dev. Cell 2010, 18, 510–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Townley-Tilson, W.H.; Callis, T.E.; Wang, D. MicroRNAs 1, 133, and 206: Critical factors of skeletal and cardiac muscle development, function, and disease. Int. J. Biochem. Cell Biol. 2010, 42, 1252–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.F.; Mandel, E.M.; Thomson, J.M.; Wu, Q.; Callis, T.E.; Hammond, S.M. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nat. Genet. 2006, 38, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Ambros, V. An extensive class of small RNAs in Caenorhabditis elegans. Science 2001, 294, 862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sempere, L.F.; Freemantle, S.; Pitharowe, I.; Moss, E.; Dmitrovsky, E.; Ambros, V. Expression profiling of mammalian microRNAs uncovers a subset of brain-expressed microRNAs with possible roles in murine and human neuronal differentiation. Genome Biol. 2004, 5, 1–11. [Google Scholar]
- Fu, H.; Tie, Y.; Xu, C.; Zhang, Z.; Zhu, J.; Shi, Y.; Jiang, H.; Sun, Z.; Zheng, X. Identification of human fetal liver miRNAs by a novel method. Febs Lett. 2005, 579, 3849–3854. [Google Scholar] [CrossRef] [Green Version]
- Shaham, L.; Vendramini, E.; Ge, Y.; Goren, Y.; Birger, Y.; Tijssen, M.R.; McNulty, M.; Geron, I.; Schwartzman, O.; Goldberg, L.; et al. Microrna-486-5p is an erythroid oncomir of the myeloid leukemias of down syndrome. Blood 2015, 125, 1292–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.S.; Li, L.; Li, L.; Chu, S.; Shiang, K.D.; Li, M.; Sun, H.Y.; Xu, J.; Xiao, F.; Sun, G.; et al. Microrna-486 regulates normal erythropoiesis and enhances growth and modulates drug response in cml progenitors. Blood 2015, 125, 1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.; Liu, S.; Wietelmann, A.; Kojonazarov, B.; Atzberger, A.; Tang, C.; Schermuly, R.T.; Gröne, H.J.; Offermanns, S. Developmental vascular remodeling defects and postnatal kidney failure in mice lacking Gpr116 (Adgrf5) and Eltd1 (Adgrl4). PLoS ONE 2017, 12, e0183166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickson, M.E.; Sigmund, C.D. Genetic basis of hypertension: Revisiting angiotensinogen. Hypertension 2006, 48, 14–20. [Google Scholar] [CrossRef]
- Lee, K.Y.; Chan KY, Y.; Tsang, K.S.; Chen, Y.C.; Kung, H.F.; Ng, P.C.; Li, C.K.; Leung, K.T.; Li, K. Ubiquitous expression of Makorin-2 in normal and malignant hematopoietic cells and its growth promoting activity. PLoS ONE 2014, 9, e92706. [Google Scholar] [CrossRef]
- Park, K.C.; Lee, D.C.; Yeom, Y.I. NDRG3-mediated lactate signaling in hypoxia. BMB Rep. 2015, 48, 301–302. [Google Scholar] [CrossRef] [Green Version]
- Mickiewicz, A.; Rybarczyk, A.; Sarzynska, J.; Figlerowicz, M.; Blazewicz, J. AmiRNA Designer-new method of artificial miRNA design. Acta Biochim. Pol. 2016, 63, 71–77. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Control Group | Experimental Group |
|---|---|---|
| Cornstarch | 397.47 | 378.29 |
| Casein (>85% protein) | 200 | 200 |
| Dextrinized cornstarch (90%–94% tetrasaccharides) | 110.38 | 125.6 |
| Sucrose | 100 | 100 |
| Soybean oil (no additives) | 70 | 70 |
| Fiber | 50 | 50 |
| Mineral mix (AIN-93G-MX) | 35 | 35 |
| Vitamin mix (AIN-93-VX) | 10 | 10 |
| L-Cystine | 3 | 3 |
| Choline bitartrate (41.1% choline) | 2.5 | 2.5 |
| Tert-butylhydroquinone | 0.014 | 0.014 |
| L-Arginine | 0 | 6.4 * |
| Glutamic acid | 21.62 ** | 0 |
| Nutrient content | ||
| Digestible energy (MJ/kg) | 15.925 | 15.925 |
| CP (%) | 18.287 | 18.287 |
| E/P | 0.871 | 0.871 |
| L-Arginine (%) | 0.64 | 1.28 |
| Glutamic acid (%) | 5.55 | 3.63 |
| No. | miRNA | Control Tpm | Arg Tpm | FC (Arg/Control) | p-Value | FDR | Expression Level |
|---|---|---|---|---|---|---|---|
| 1 | miR-181a-1-3p | 613.34 | 422.53 | 0.69 | 2.2 × 10−9 | 4.1 × 10−8 | down |
| 2 | miR-141-3p | 21,066.09 | 14,791.93 | 0.70 | 0 | 0 | down |
| 3 | miR-192-5p | 903.41 | 654.03 | 0.72 | 1.9 × 10−10 | 3.8 × 10−9 | down |
| 4 | miR-200a-3p | 2234.33 | 1643.09 | 0.74 | 8.3 × 10−22 | 3.0 × 10−20 | down |
| 5 | miR-181c-5p | 2070.42 | 1531.36 | 0.74 | 1.2 × 10−19 | 3.7 × 10−18 | down |
| 6 | miR-200b-3p | 4278.14 | 3427.11 | 0.80 | 7.8 × 10−23 | 3.3 × 10−21 | down |
| 7 | miR-429 | 2578.68 | 2074.59 | 0.80 | 6.8 × 10−14 | 1.7 × 10−12 | down |
| 8 | miR-101a-3p | 5917.868 | 5023.77 | 0.85 | 3.0 × 10−18 | 8.6 × 10−17 | down |
| 9 | let-7f-5p | 17,008.48 | 14,530.47 | 0.85 | 1.9 × 10−46 | 1.1 × 10−44 | down |
| 10 | let-7g-5p | 3540.63 | 3050.85 | 0.86 | 7.9 × 10−10 | 1.5 × 10−8 | down |
| 11 | miR-148a-5p | 7142.35 | 6155.98 | 0.86 | 2.3 × 10−18 | 7.0 × 10−17 | down |
| 12 | miR-181a-5p | 48,487.41 | 42,283.78 | 0.87 | 0 | 0 | down |
| 13 | miR-186-5p | 4608.16 | 4080.20 | 0.89 | 6.8× 10−9 | 1.1 × 10−7 | down |
| 14 | miR-26b-5p | 2903.73 | 2592.21 | 0.89 | 1.7 × 10−5 | 2.7 × 10−4 | down |
| 15 | miR-148a-3p | 237,987.63 | 214,042.02 | 0.90 | 0 | 0 | down |
| 16 | miR-31a-5p | 2602.24 | 2368.22 | 0.91 | 6.5 × 10−4 | 7.4 × 10−3 | down |
| 17 | miR-30e-5p | 5237.25 | 4815.07 | 0.92 | 1.4 × 10−5 | 2.2 × 10−4 | down |
| 18 | let-7i-5p | 3746.97 | 3451.53 | 0.92 | 3.3 × 10−4 | 4.0 × 10−3 | down |
| 19 | let-7a-5p | 4848.41 | 4531.16 | 0.93 | 6.6 × 10−4 | 7.3 × 10−3 | down |
| 20 | miR-30a-5p | 28,750.68 | 27,008.46 | 0.94 | 6.2 × 10−15 | 1.6 × 10−13 | down |
| 21 | miR-375-3p | 43,642.12 | 41,710.08 | 0.96 | 8.5 × 10−13 | 2.0 × 10−11 | down |
| 22 | miR-26a-5p | 54,560.24 | 53,471.05 | 0.98 | 1.2 × 10−4 | 1.5 × 10−4 | down |
| 23 | miR-10a-5p | 36,589.32 | 37,617.76 | 1.03 | 5.0 × 10−4 | 5.9 × 10−3 | up |
| 24 | miR-30d-5p | 9351.35 | 9923.60 | 1.06 | 7.7 × 10−5 | 1.0 × 10−3 | up |
| 25 | miR-27b-3p | 13,604.16 | 14,733.03 | 1.08 | 6.6 × 10−11 | 1.4 × 10−9 | up |
| 26 | let-7c-5p | 4151.56 | 4503.59 | 1.08 | 2.5 × 10−4 | 3.1 × 10−4 | up |
| 27 | miR-151-3p | 3099.21 | 3377.26 | 1.09 | 8.1 × 10−4 | 8.4 × 10−4 | up |
| 28 | miR-126a-5p | 6980.50 | 7703.56 | 1.10 | 5.7 × 10−9 | 1.0 × 10−7 | up |
| 29 | miR-146b-5p | 8007.73 | 8901.84 | 1.11 | 1.7 × 10−11 | 3.8 × 10−10 | up |
| 30 | miR-99b-5p | 1625.39 | 1824.40 | 1.12 | 9.5 × 10−4 | 9.6 × 10−3 | up |
| 31 | miR-205 | 1579.97 | 1785.98 | 1.13 | 5.2 × 10−3 | 6.0 × 10−3 | up |
| 32 | miR-92a-3p | 1888.21 | 2152.01 | 1.14 | 4.8 × 10−5 | 6.8 × 10−4 | up |
| 33 | miR-191a-5p | 8581.33 | 9838.75 | 1.15 | 7.2 × 10−20 | 2.4 × 10−18 | up |
| 34 | miR-22-3p | 33,500.82 | 38,702.25 | 1.16 | 1.1 × 10−83 | 8.2 × 10−82 | up |
| 35 | miR-10b-5p | 168,843.19 | 196,136.60 | 1.16 | 0 | 0 | up |
| 36 | miR-181b-5p | 792.72 | 935.12 | 1.18 | 7.5 × 10−4 | 8.0 × 10−3 | up |
| 37 | miR-199a-3p | 1008.57 | 1203.49 | 1.19 | 4.4 × 10−5 | 6.4 × 10−4 | up |
| 38 | miR-126a-3p | 1729.27 | 2068.87 | 1.20 | 5.7 × 10−8 | 9.4 × 10−7 | up |
| 39 | miR-16-5p | 4468.45 | 5442.55 | 1.22 | 3.5 × 10−22 | 1.4 × 10−20 | up |
| 40 | miR-21-5p | 7391.90 | 9253.61 | 1.25 | 1.3 × 10−46 | 8.1 × 10−45 | up |
| 41 | miR-378a-3p | 6381.27 | 8094.68 | 1.27 | 1.9 × 10−45 | 9.8 × 10−44 | up |
| 42 | miR-145-5p | 280.35 | 367.30 | 1.31 | 7.3 × 10−4 | 7.9 × 10−3 | up |
| 43 | miR-150-5p | 232.95 | 327.95 | 1.41 | 7.0 × 10−5 | 9.6 × 10−4 | up |
| 44 | miR-486 | 1157.86 | 2573.86 | 2.22 | 0 | 0 | up |
| 45 | miR-133a-3p | 900.15 | 5278.65 | 5.86 | 0 | 0 | up |
| 46 | miR-206-3p | 4.26 | 25.52 | 5.99 | 1.9 × 10−4 | 2.4 × 10−3 | up |
| 47 | miR-1b | 27.67 | 168.76 | 6.10 | 9.3 × 10−26 | 4.3 × 10−24 | up |
| 48 | miR-1-3p | 14.47 | 96.03 | 6.63 | 7.2 × 10−16 | 2.0 × 10−14 | up |
| 49 | miR-133b-3p | 8.66 | 58.73 | 6.78 | 3.9 × 10−10 | 7.7 × 10−9 | up |
| 50 | miR-133a-5p | 2.98 | 24.93 | 8.367 | 2.7 × 10−5 | 4.1 × 10−4 | up |
| miRNAs | GO Enrichment | KEGG Pathway Enrichment | p- Value | FDR | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Different Genein All GO | Up Gene | Down Gene | All Gene in All GO | Different Genein All Pathway | Up Gene | Down Gene | All Gene in All Pathway | Score | |||
| miR-1-3p | 273 | 0 | 273 | 19,087 | 109 | 0 | 109 | 7597 | >1 | <0.05 | <0.25 |
| miR-206-3p | 271 | 0 | 271 | 19,087 | 108 | 0 | 108 | 7597 | >1 | <0.05 | <0.25 |
| miR-133a-3p | 222 | 0 | 222 | 19,087 | 80 | 0 | 80 | 7597 | >1 | <0.05 | <0.25 |
| miR-133b-3p | 222 | 0 | 222 | 19,087 | 80 | 0 | 80 | 7597 | >1 | <0.05 | <0.25 |
| miR-133a-5p | 129 | 0 | 129 | 19,087 | 52 | 0 | 52 | 7597 | >1 | <0.05 | <0.25 |
| Total different gene | 558 | 240 | |||||||||
| Gene ID | Gene Symbol | NCBI Accession | Description | miRNAs |
|---|---|---|---|---|
| 296566 | Clic3 | NM_001013080.2 | chloride intracellular channel 3 | rno-miR-1-3p |
| 25321 | Ghrhr | NM_012850.1 | growth hormone releasing hormone receptor | rno-miR-1-3p |
| 360622 | Igfbp4 | NM_001004274.2 | insulin-like growth factor binding protein 4 | rno-miR-1-3p |
| 60326 | Kcnv1 | NM_021697.1 | potassium channel, subfamily V, member 1 | rno-miR-1-3p |
| 291439 | Mbd1 | NM_001011924.1 | methyl-CpG binding domain protein 1 | rno-miR-1-3p |
| 307271 | Mex3c | NM_001107377.1 | mex-3 homolog C (C. elegans) | rno-miR-1-3p |
| 297525 | Mkrn2 | NM_001008314.1 | makorin, ring finger protein, 2 | rno-miR-1-3p |
| 25482 | Mras | NM_012981.2 | muscle RAS oncogene homolog | rno-miR-1-3p |
| 287962 | Mrpl40 | NM_001024865.1 | mitochondrial ribosomal protein L40 | rno-miR-1-3p |
| 289482 | Naa11(Ard1b) | NM_001024742.1 | ARD1 homolog B (S. cerevisiae) | rno-miR-1-3p |
| 296318 | Ndrg3 | NM_001013923.1 | N-myc downstream regulated gene 3 | rno-miR-1-3p |
| 360967 | Rnf185 | NM_001024271.1 | ring finger protein 185 | rno-miR-1-3p |
| 29503 | Slc22a2 | NM_031584.2 | solute carrier family 22 (organic cation transporter), member 2 | rno-miR-1-3p |
| 114247 | Slfn3 | NM_053687.1 | schlafen 3 | rno-miR-1-3p |
| 25033 | Sstr1 | NM_012719.2 | somatostatin receptor 1 | rno-miR-1-3p |
| 362453 | Crebl2 | NM_001015027.1 | cAMP responsive element binding protein-like 2 | rno-miR-206-3p |
| 289693 | Dhx15 | NM_001191597.1 | DEAH (Asp-Glu-Ala-His) box polypeptide 15 | rno-miR-206-3p |
| 64124 | Adgrl4(Eltd1) | NM_022294.1 | EGF, latrophilin and seven transmembrane domain containing 1 | rno-miR-206-3p |
| 362862 | Hsp90b1 | NM_001012197.2 | heat shock protein 90kDa beta (Grp94), member 1 | rno-miR-206-3p |
| 192361 | Ppp1r2 | NM_138823.2 | protein phosphatase 1, regulatory (inhibitor) subunit 2 | rno-miR-206-3p |
| 54190 | Rabep1 | NM_019124.1 | rabaptin, RAB GTPase binding effector protein 1 | rno-miR-206-3p |
| 192152 | Tgoln2 | NM_138840.2 | trans-golgi network protein | rno-miR-206-3p |
| 300857 | Tmem30a | NM_001004248.1 | transmembrane protein 30A | rno-miR-206-3p |
| 113922 | 15-Sep | NM_133297.2 | selenoprotein | rno-miR-133a-3p |
| 300677 | Atp5l | NM_212516.2 | ATP synthase, H+ transporting, mitochondrial F0 complex, subunit G | rno-miR-133a-3p |
| 24565 | Abcc1 | NM_022281.2 | ATP-binding cassette, subfamily C (CFTR/MRP), member 1 | rno-miR-133b-3p |
| 25402 | Casp3 | NM_012922.2 | caspase 3 | rno-miR-133b-3p |
| 298098 | Pole3 | NM_001007652.2 | polymerase (DNA directed), epsilon 3 (p17 subunit) | rno-miR-133b-3p |
| 295347 | Rap1a | NM_001005765.1 | RAP1A, member of RAS oncogene family | rno-miR-133b-3p |
| 54296 | Rgs7 | NM_019343.1 | regulator of G-protein signaling 7 | rno-miR-133b-3p |
| 29726 | Slc22a5 | NM_019269.1 | solute carrier family 22 (organic cation/carnitine transporter), member 5 | rno-miR-133b-3p |
| 362691 | Tmem178 | NM_001004282.1 | transmembrane protein 178 | rno-miR-133b-3p |
| 24179 | Agt | NM_134432.2 | angiotensinogen (serpin peptidase inhibitor, clade A, member 8) | rno-miR-133a-5p |
| Arginine vs. Control | Target Genes | n | p-Value |
|---|---|---|---|
| No results | Slc22a2, Clic3, Kcnv1, Igfbp4, Sstr1, Ghrhr | 6 vs. 6 | |
| No difference | Mbd1, Naa11(Ard1b), Mras, Mrpl40, Rap1a, Rgs7, Abcc1, Pole3, Crebl2, Rabep1 | 6 vs. 6 | |
| Up gene | Mkrn2, Slfn3, Mex3c, Rnf185, Ndrg3, Tgoln2, Dhx15, Slc22a5, Casp3, Tmem178a, E1td1(Adgrl4), Hsp90b1, Tmem30a, Ppp1r2, Agt | 6 vs. 6 | 0.05 |
| Down gene | Atp5l, 15-Sep | 6 vs. 6 | 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, G.; Xu, Q.; Wu, F.; Wang, M.; Chen, L.; Hu, L.; Zhao, J.; Loor, J.J.; Zhang, J. Arginine Alters miRNA Expression Involved in Development and Proliferation of Rat Mammary Tissue. Animals 2021, 11, 535. https://doi.org/10.3390/ani11020535
Zhou G, Xu Q, Wu F, Wang M, Chen L, Hu L, Zhao J, Loor JJ, Zhang J. Arginine Alters miRNA Expression Involved in Development and Proliferation of Rat Mammary Tissue. Animals. 2021; 11(2):535. https://doi.org/10.3390/ani11020535
Chicago/Turabian StyleZhou, Gang, Qiaoyun Xu, Feifan Wu, Mengzhi Wang, Lianmin Chen, Liangyu Hu, Jingwen Zhao, Juan J. Loor, and Jun Zhang. 2021. "Arginine Alters miRNA Expression Involved in Development and Proliferation of Rat Mammary Tissue" Animals 11, no. 2: 535. https://doi.org/10.3390/ani11020535