
Programming Effect of the Parental Obesity on the Skeletal System of Offspring at Weaning Day

 ,
,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Procedures
2.2. Dual X-ray Absorptiometry (DXA) Analysis of the Total Skeleton and Isolated Tibiae
2.3. Peripheral Quantitative Computed Tomography (pQCT) Measurements of the Isolated Tibiae
2.4. 3D Micro-Computed Tomography (µ-CT) Analysis of Trabecular Bone Tissue Morphometry
2.5. Mechanical Properties
2.6. Biochemical Analysis
2.7. Statistical Analysis
3. Results
3.1. Body Weight, Body Composition, Tibia Length and Mass
3.2. Mass of the Subcutaneous, Visceral and Mesenteric Fat Tissue
3.3. DXA Measurements of the Whole Skeleton and the Isolated Tibia
3.4. pQCT Analysis of Tibia
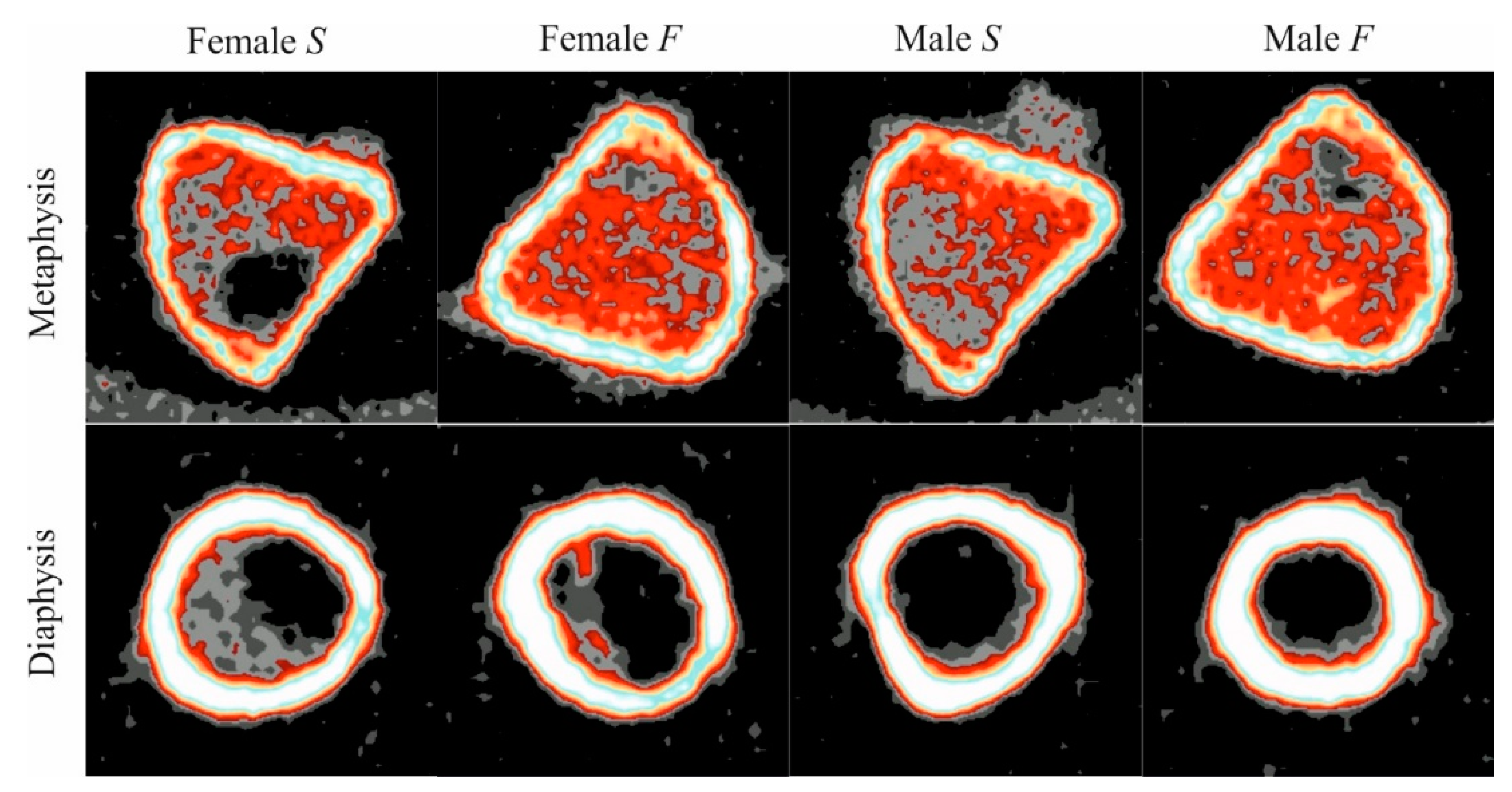
3.5. µCT Analysis of Trabecular Bone Tissue in the Proximal Metaphysis
3.6. Mechanical Analysis of Tibia
3.7. Biochemistry
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Moura, E.G.; Lisboa, P.C.; Passos, M.C. Neonatal programming of neuroimmunomodulation—Role of adipocytokines and neuropeptides. Neuroimmunomodulation 2008, 15, 176–188. [Google Scholar] [CrossRef]
- De Moura, E.G.; Passos, M.C. Neonatal programming of body weight regulation and energetic metabolism. Biosci. Rep. 2005, 25, 251–269. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.Y.; Yoon, K.H. Epidemic obesity in children and adolescents: Risk factors and prevention. Front. Med. 2018, 12, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Mohan, R.; Baumann, D.; Alejandro, E.U. Fetal undernutrition, placental insufficiency, and pancreatic beta-cell development programming in utero. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R867–R878. [Google Scholar] [CrossRef] [Green Version]
- Devlin, M.J.; Bouxsein, M.L. Influence of pre- and peri-natal nutrition on skeletal acquisition and maintenance. Bone 2012, 50, 444–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGown, C.; Birerdinc, A.; Younossi, Z.M. Adipose tissue as an endocrine organ. Clin. Liver Dis. 2014, 18, 41–58. [Google Scholar] [CrossRef]
- Bienko, M.; Lis, A.; Wolski, D.; Radzki, R.P. Relationships between bone tissue and fat tissue. Med. Weter. 2016, 72, 217–224. [Google Scholar]
- Calabro, P.; Yeh, E.T. Obesity, inflammation, and vascular disease: The role of the adipose tissue as an endocrine organ. Subcell Biochem. 2007, 42, 63–91. [Google Scholar] [PubMed]
- De Laet, C.; Kanis, J.A.; Oden, A.; Johanson, H.; Johnell, O.; Delmas, P.; Eisman, J.A.; Kroger, H.; Fujiwara, S.; Garnero, P.; et al. Body mass index as a predictor of fracture risk: A meta-analysis. Osteoporos Int. 2005, 16, 1330–1338. [Google Scholar] [CrossRef]
- Hwang, D.K.; Choi, H.J. The relationship between low bone mass and metabolic syndrome in Korean women. Osteoporos Int. 2010, 21, 425–431. [Google Scholar] [CrossRef]
- Sakata, T.; Halloran, B.P.; Elalieh, H.Z.; Munson, S.J.; Rudner, L.; Venton, L.; Ginzinger, D.; Rosen, C.J.; Bikle, D.D. Skeletal unloading induces resistance to insulin-like growth factor I on bone formation. Bone 2003, 32, 669–680. [Google Scholar] [CrossRef]
- Blum, M.; Harris, S.S.; Must, A.; Naumova, E.N.; Phillips, S.M.; Rand, W.M.; Dawson-Hughes, B. Leptin, body composition and bone mineral density in premenopausal women. Calcif. Tissue Int. 2003, 73, 27–32. [Google Scholar] [CrossRef]
- Hsu, Y.H.; Venners, S.A.; Terwedow, H.A.; Feng, Y.; Niu, T.; Li, Z.; Laird, N.; Brain, J.D.; Cummings, S.R.; Bouxsein, M.L.; et al. Relation of body composition, fat mass, and serum lipids to osteoporotic fractures and bone mineral density in Chinese men and women. Am. J. Clin. Nutr. 2006, 83, 146–154. [Google Scholar] [CrossRef] [Green Version]
- Dobrowolski, P.; Tomaszewska, E.; Bienko, M.; Radzki, R.P.; Pierzynowski, S.G. The effect of dietary administration of 2-oxoglutaric acid on the cartilage and bone of growing rats. Br. J. Nutr. 2013, 110, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Eastell, R.; Lambert, H. Diet and healthy bones. Calcif. Tissue Int. 2002, 70, 400–404. [Google Scholar] [CrossRef]
- Topolska, K.; Radzki, R.P.; Filipiak-Florkiewicz, A.; Florkiewicz, A.; Leszczynska, T.; Cieslik, E. Fructan-Enriched Diet Increases Bone Quality in Female Growing Rats at Calcium Deficiency. Plant Foods Hum. Nutr. 2018, 73, 172–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howie, G.J.; Sloboda, D.M.; Kamal, T.; Vickers, M.H. Maternal nutritional history predicts obesity in adult offspring independent of postnatal diet. J. Physiol. 2009, 587, 905–915. [Google Scholar] [CrossRef]
- Longhi, R.; Almeida, R.F.; Machado, L.; Duarte, M.; Souza, D.G.; Machado, P.; de Assis, A.M.; Quincozes-Santos, A.; Souza, D.O. Effect of a trans fatty acid-enriched diet on biochemical and inflammatory parameters in Wistar rats. Eur. J. Nutr. 2017, 56, 1003–1016. [Google Scholar] [CrossRef]
- Lac, G.; Cavalie, H.; Ebal, E.; Michaux, O. Effects of a high fat diet on bone of growing rats. Correlations between visceral fat, adiponectin and bone mass density. Lipids Health Dis. 2008, 7, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koolhaas, J.M. The laboratory rat. In The UFAW Handbook on the Care and Management of Laboratory and Other Research Animals, 8th ed.; Hubrecht, R., Kirkwood, J., Eds.; The Universities Federation for Animal Welfare: Hertfordshire, UK, 2010; pp. 311–326. [Google Scholar]
- Radzki, R.P.; Bienko, M.; Polak, P.; Szkucik, K.; Ziomek, M.; Ostapiuk, M.; Bienias, J. Is the consumption of snail meat actually healthy? An analysis of the osteotropic influence of snail meat as a sole source of protein in growing rats. J. Anim. Physiol. Anim. Nutr. 2018, 102, e885–e891. [Google Scholar] [CrossRef]
- Rodrigues, A.L.; de Moura, E.G.; Passos, M.C.; Dutra, S.C.; Lisboa, P.C. Postnatal early overnutrition changes the leptin signalling pathway in the hypothalamic-pituitary-thyroid axis of young and adult rats. J. Physiol. 2009, 587, 2647–2661. [Google Scholar] [CrossRef] [PubMed]
- Conceicao, E.P.; Franco, J.G.; Oliveira, E.; Resende, A.C.; Amaral, T.A.; Peixoto-Silva, N.; Passos, M.C.; Moura, E.G.; Lisboa, P.C. Oxidative stress programming in a rat model of postnatal early overnutrition—Role of insulin resistance. J. Nutr. Biochem. 2013, 24, 81–87. [Google Scholar] [CrossRef] [PubMed]
- De Albuquerque Maia, L.; Lisboa, P.C.; de Oliveira, E.; da Conceicao, E.P.; Lima, I.C.; Lopes, R.T.; Ruffoni, L.D.; Nonaka, K.O.; de Moura, E.G. Bone structure and strength are enhanced in rats programmed by early overfeeding. Horm. Metab. Res. 2014, 46, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Madeira, E.; Mafort, T.T.; Madeira, M.; Guedes, E.P.; Moreira, R.O.; de Mendonca, L.M.; Lima, I.C.; de Pinho, P.R.; Lopes, A.J.; Farias, M.L. Lean mass as a predictor of bone density and microarchitecture in adult obese individuals with metabolic syndrome. Bone 2014, 59, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Dimitri, P. Fat and bone in children—Where are we now? Ann. Pediatr. Endocrinol. Metab. 2018, 23, 62–69. [Google Scholar] [CrossRef]
- El Hage, R.; Bachour, F.; Sebaaly, A.; Issa, M.; Zakhem, E.; Maalouf, G. The influence of weight status on radial bone mineral density in Lebanese women. Calcif. Tissue Int. 2014, 94, 465–467. [Google Scholar] [CrossRef]
- Møgaard, C.; Hoppe, C.; Michaelsen, K.F. Bone mineral content (BMC) at 10 years of age is related to growth in infancy. Pediatric Res. 1999, 45, 915. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.R.; Bok, S.C.; Choi, S.C.; Lee, S.S.; Heo, M.S.; Huh, K.H.; Kim, T.I.; Yi, W.J. The relationship between dental implant stability and trabecular bone structure using cone-beam computed tomography. J. Periodontal Implant Sci. 2016, 46, 116–127. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Ahn, J.; Kim, H.K.; Kim, J.H. Obese children experience more extremity fractures than nonobese children and are significantly more likely to die from traumatic injuries. Acta Paediatr. 2016, 105, 1152–1157. [Google Scholar] [CrossRef]
- Goulding, A.; Grant, A.M.; Williams, S.M. Bone and body composition of children and adolescents with repeated forearm fractures. J. Bone Miner. Res. 2005, 20, 2090–2096. [Google Scholar] [CrossRef]
- Manias, K.; McCabe, D.; Bishop, N. Fractures and recurrent fractures in children; varying effects of environmental factors as well as bone size and mass. Bone 2006, 39, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Jones, I.E.; Taylor, R.W.; Williams, S.M.; Manning, P.J.; Goulding, A. Four-year gain in bone mineral in girls with and without past forearm fractures: A DXA study. Dual energy X-ray absorptiometry. J. Bone Miner. Res. 2002, 17, 1065–1072. [Google Scholar] [CrossRef]
- Wosje, K.S.; Khoury, P.R.; Claytor, R.P.; Copeland, K.A.; Hornung, R.W.; Daniels, S.R.; Kalkwarf, H.J. Dietary patterns associated with fat and bone mass in young children. Am. J. Clin. Nutr. 2010, 92, 294–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plesner, J.L.; Dahl, M.; Fonvig, C.E.; Nielsen, T.R.H.; Kloppenborg, J.T.; Pedersen, O.; Hansen, T.; Holm, J.C. Obesity is associated with vitamin D deficiency in Danish children and adolescents. J. Pediatr. Endocrinol. Metab. 2018, 31, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Black, R.E.; Williams, S.M.; Jones, I.E.; Goulding, A. Children who avoid drinking cow milk have low dietary calcium intakes and poor bone health. Am. J. Clin. Nutr. 2002, 76, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Sudhagoni, R.G.; Wey, H.E.; Djira, G.D.; Specker, B.L. Longitudinal effects of fat and lean mass on bone accrual in infants. Bone 2012, 50, 638–642. [Google Scholar] [CrossRef]
- Burrows, M.; Baxter-Jones, A.; Mirwald, R.; Macdonald, H.; McKay, H. Bone mineral accrual across growth in a mixed-ethnic group of children: Are Asian children disadvantaged from an early age? Calcif. Tissue Int. 2009, 84, 366–378. [Google Scholar] [CrossRef]
- Laddu, D.R.; Farr, J.N.; Laudermilk, M.J.; Lee, V.R.; Blew, R.M.; Stump, C.; Houtkooper, L.; Lohman, T.G.; Going, S.B. Longitudinal relationships between whole body and central adiposity on weight-bearing bone geometry, density, and bone strength: A pQCT study in young girls. Arch. Osteoporos. 2013, 8, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferretti, J.L.; Capozza, R.F.; Zanchetta, J.R. Mechanical validation of a tomographic (pQCT) index for noninvasive estimation of rat femur bending strength. Bone 1996, 18, 97–102. [Google Scholar] [CrossRef]
- Gajewska, J.; Ambroszkiewicz, J.; Radomyska, B.; Chelchowska, M.; Oltarzewski, M.; Laskowska-Klita, T.; Milanowski, A. Serum markers of bone turnover in children and adolescents with classic galactosemia. Adv. Med. Sci. 2008, 53, 214–220. [Google Scholar] [CrossRef] [Green Version]
- Bell, N.H.; Epstein, S.; Greene, A.; Shary, J.; Oexmann, M.J.; Shaw, S. Evidence for alteration of the vitamin D-endocrine system in obese subjects. J. Clin. Invest. 1985, 76, 370–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Albuquerque Maia, L.; Lisboa, P.C.; de Oliveira, E.; da Silva Lima, N.; da Costa, C.A.; de Moura, E.G. Two models of early weaning decreases bone structure by different changes in hormonal regulation of bone metabolism in neonate rat. Horm. Metab. Res. 2013, 45, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Cirulli, F.; Musillo, C.; Berry, A. Maternal Obesity as a Risk Factor for Brain Development and Mental Health in the Offspring. Neuroscience 2020, 447, 122–135. [Google Scholar] [CrossRef]
- McPherson, N.O.; Fullston, T.; Aitken, R.J.; Lane, M. Paternal obesity, interventions, and mechanistic pathways to impaired health in offspring. Ann. Nutr. Metab. 2014, 64, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Eberle, C.; Kirchner, M.F.; Herden, R.; Stichling, S. Paternal metabolic and cardiovascular programming of their offspring: A systematic scoping review. PLoS ONE 2020, 15, e0244826. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Ingredients | Standard Diet (S) | High-Energy Diet (F) |
|---|---|---|
| Metabolic Energy | 11.50 MJ/kg | 17.6 MJ/kg |
| Crude protein | 16.00% | 20.0% |
| Crude fat min. | 2.80% | 21.0% |
| Crude ash max. | 7.00% | 5.58% |
| Crude fiber max. | 5.00% | 3.86% |
| Lysine | 0.80% | 1.2% |
| Methionine + Cysteine | 0.50% | 0.76% |
| Tryptophan | 0.190% | 0.27% |
| Calcium | 1.10% | 1.32% |
| Phosphorus | 0.70% | 0.54% |
| Natrium | 0.22% | 0.19% |
| Vitamin A | 8000 IU/kg | 8000 IU/kg |
| Vitamin D3 | 1000 IU/kg | 1000 IU/kg |
| Vitamin E | 50 mg/kg | 76.9 mg/kg |
| Item | Female | Male | ||||
|---|---|---|---|---|---|---|
| S | F | p-Value | S | F | p-Value | |
| Body weight (birthday) (g) | 8.2 ± 0.2 | 9.5 ± 0.3 | 0.004 * | 8.5 ± 0.2 | 9.4 ± 0.2 | 0.002 * |
| Body weight (weaning) (g) | 60.0 ± 2.3 | 70.4 ± 1.4 | 0.002 * | 66.0 ± 0.9 | 85.3 ± 1.1 | <0.0001 * |
| Subcutaneous fat (g) | 1.03 ± 0.04 | 1.25 ± 0.09 | 0.036 * | 1.08 ± 0.08 | 1.35 ± 0.10 | 0.046 * |
| Visceral fat (g) | 0.21 ± 0.03 | 0.54 ± 0.07 | 0.008 * | 0.21 ± 0.03 | 0.51 ± 0.09 | 0.003 * |
| Mesenteric fat (g) | 0.38 ± 0.03 | 0.53 ± 0.03 | 0.007 * | 0.40 ± 0.04 | 0.51 ± 0.02 | 0.020 * |
| Tibia mass (g) | 0.104 ± 0.004 | 0.107 ± 0.003 | 0.243 | 0.104 ± 0.003 | 0.109 ± 0.003 | 0.187 |
| Tibia length (mm) | 23.2 ± 0.4 | 22.0 ± 0.2 | 0.301 | 22.9 ± 0.5 | 22.2 ± 0.2 | 0.277 |
| Soft tissue mass (ST) (g) | 53.8 ± 2.2 | 57.9 ± 2.4 | 0.227 | 57.5 ± 2.2 | 61.2 ± 2.2 | 0.267 |
| Fat tissue mass (FM) (g) | 0.060 ± 0.009 | 0.552 ± 0.079 | <0.0001 * | 0.072 ± 0.009 | 1.104 ± 0.085 | <0.0001 * |
| Lean mass (LM) (g) | 57.4 ± 1.8 | 58.4 ± 2.0 | 0.730 | 61.4 ± 1.4 | 63.5 ± 1.7 | 0.345 |
| Item | Female | Male | ||||
|---|---|---|---|---|---|---|
| S | F | p-Value | S | F | p-Value | |
| pQCT analysis of tibia diaphysis (50% of bone length) | ||||||
| Tot.BMC (mg/mm) | 0.86 ± 0.05 | 0.92 ± 0.06 | 0.523 | 0.86 ± 0.04 | 0.98 ± 0.03 | 0.053 |
| Tot.vBMD (mg/mm3) | 406 ± 13 | 422 ± 20 | 0.719 | 396 ± 18 | 423 ± 14 | 0.260 |
| Tot.Ar (mm2) | 2.15 ± 0.03 | 2.19 ± 0.08 | 0.734 | 2.20 ± 0.10 | 2.32 ± 0.08 | 0.378 |
| Peri.C (mm) | 5.2 ± 0.1 | 5.2 ± 0.1 | 0.749 | 5.3 ± 0.1 | 5.4 ± 0.1 | 0.366 |
| Endo.C (mm) | 3.8 ± 0.1 | 3.7 ± 0.1 | 0.890 | 3.8 ± 0.1 | 3.9 ± 0.1 | 0.444 |
| Ct.BMC (mg/mm) | 0.85 ± 0.03 | 0.93 ± 0.04 | 0.032 * | 0.86 ± 0.02 | 0.96 ± 0.02 | 0.006 * |
| (Ct.vBMD (mg/mm3) | 786 ± 7 | 841 ± 9 | 0.032 * | 814 ± 6 | 871 ± 9 | 0.016 * |
| Ct.Ar (mm2) | 1.02 ± 0.02 | 1.08 ± 0.03 | 0.133 | 1.06 ± 0.03 | 1.11 ± 0.02 | 0.250 |
| Ct.Th (mm) | 0.229 ± 0.002 | 0.242 ± 0.004 | 0.036 * | 0.234 ± 0.002 | 0.254 ± 0.003 | 0.001 * |
| pQCT analysis of tibia proximal metaphysis | ||||||
| Tot.BMC (mg/mm) | 1.6 ± 0.1 | 1.9 ± 0.1 | 0.017 * | 1.7 ± 0.1 | 2.1 ± 0.1 | 0.006 * |
| Tot.vBMD (mg/mm3) | 271 ± 8 | 322 ± 14 | 0.007 * | 254 ± 9 | 322 ± 11 | 0.0003 * |
| Tot.Ar (mm2) | 6.0 ± 0.2 | 6.0 ± 0.3 | 0.892 | 6.6 ± 0.4 | 6.5 ± 0.3 | 0.789 |
| Tb.BMC (mg/mm) | 0.28 ± 0.02 | 0.40 ± 0.03 | 0.005 * | 0.29 ± 0.03 | 0.42 ± 0.03 | 0.012 * |
| (Tb.vBMD (mg/mm3) | 92 ± 12 | 143 ± 14 | 0.018 * | 80 ± 16 | 140 ± 14 | 0.015 * |
| Tb.Ar (mm2) | 2.7 ± 0.1 | 2.7 ± 0.2 | 0.848 | 3.0 ± 0.2 | 2.9 ± 0.1 | 0.820 |
| Item | Female | Male | ||||
|---|---|---|---|---|---|---|
| S | F | p-Value | S | F | p-Value | |
| TV (mm3) | 14.8 ± 0.8 | 15.5 ± 0.1 | 0.35 | 15.0 ± 0.5 | 15.6 ± 0.4 | 0.43 |
| BV (mm3) | 8.8 ± 0.3 | 11.6 ± 0.1 | <0.0001 * | 8.1 ± 0.2 | 12.3 ± 0.3 | <0.0001 * |
| BS (mm2) | 372 ± 16 | 351 ± 6 | 0.280 * | 398 ± 9 | 369 ± 5 | 0.015 * |
| BS/TV (1/mm) | 25.3 ± 0.6 | 22.6 ± 0.4 | 0.004 * | 26.6 ± 0.5 | 23.7 ± 0.3 | 0.0006 * |
| BV/TV (%) | 59.2 ± 1.6 | 74.2 ± 0.9 | <0.0001 * | 58.1 ± 0.9 | 79.5 ± 2.6 | <0.0001 * |
| Tb.Th (mm) | 0.21 ± 0.02 | 0.30 ± 0.02 | 0.005 * | 0.22 ± 0.01 | 0.32 ± 0.01 | <0.0007 * |
| Tb.N (1/mm) | 3.0 ± 0.1 | 2.4 ± 0.1 | 0.003 * | 2.9 ± 0.1 | 2.4 ± 0.1 | 0.0007 * |
| Tb.Sp (mm) | 0.17 ± 0.01 | 0.13 ± 0.01 | 0.006 * | 0.17 ± 0.01 | 0.12 ± 0.01 | 0.014 * |
| SMI | −1.7 ± 0.1 | −3.0 ± 0.1 | <0.0001 * | −1.6 ± 0.1 | −3.0 ± 0.1 | <0.0001 * |
| Item | Female | Male | ||||
|---|---|---|---|---|---|---|
| S | F | p-Value | S | F | p-Value | |
| Osteocalcin (ng/mL) | 10.5 ± 0.1 | 11.2 ± 0.1 | 0.001 * | 10.1 ± 0.2 | 10.9 ± 0.1 | 0.005 * |
| bALP U/L | 517 ± 43 | 803 ± 59 | 0.002 * | 539 ± 27 | 670 ± 53 | 0.002 * |
| CTX-I (ng/mL) | 30.9 ± 1.2 | 26.6 ± 0.4 | 0.003 * | 28.0 ± 0.5 | 23.0 ± 0.9 | <0.0001 * |
| 25 OH D3 (ng/mL) | 49.0 ± 2.6 | 45.9 ± 2.9 | 0.424 | 44.3 ± 1.8 | 47.5 ± 1.9 | 0.042 * |
| Phosphorus (mmol/L) | 11.7 ± 0.3 | 12.6 ± 0.8 | 0.363 | 12.6 ± 0.6 | 12.2 ± 0.5 | 0.329 |
| Calcium (mmol/L) | 11.5 ± 0.1 | 11.6 ± 0.2 | 0.750 | 11.8 ± 0.3 | 11.5 ± 0.1 | 0.942 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radzki, R.P.; Bienko, M.; Wolski, D.; Ostapiuk, M.; Polak, P.; Manastyrska, M.; Kimicka, A.; Wolska, J. Programming Effect of the Parental Obesity on the Skeletal System of Offspring at Weaning Day. Animals 2021, 11, 424. https://doi.org/10.3390/ani11020424
Radzki RP, Bienko M, Wolski D, Ostapiuk M, Polak P, Manastyrska M, Kimicka A, Wolska J. Programming Effect of the Parental Obesity on the Skeletal System of Offspring at Weaning Day. Animals. 2021; 11(2):424. https://doi.org/10.3390/ani11020424
Chicago/Turabian StyleRadzki, Radoslaw Piotr, Marek Bienko, Dariusz Wolski, Monika Ostapiuk, Pawel Polak, Malgorzata Manastyrska, Aleksandra Kimicka, and Joanna Wolska. 2021. "Programming Effect of the Parental Obesity on the Skeletal System of Offspring at Weaning Day" Animals 11, no. 2: 424. https://doi.org/10.3390/ani11020424