
Long Noncoding RNA LncPGCR Mediated by TCF7L2 Regulates Primordial Germ Cell Formation in Chickens

 , ,
, ,
Abstract
:Simple Summary
Abstract
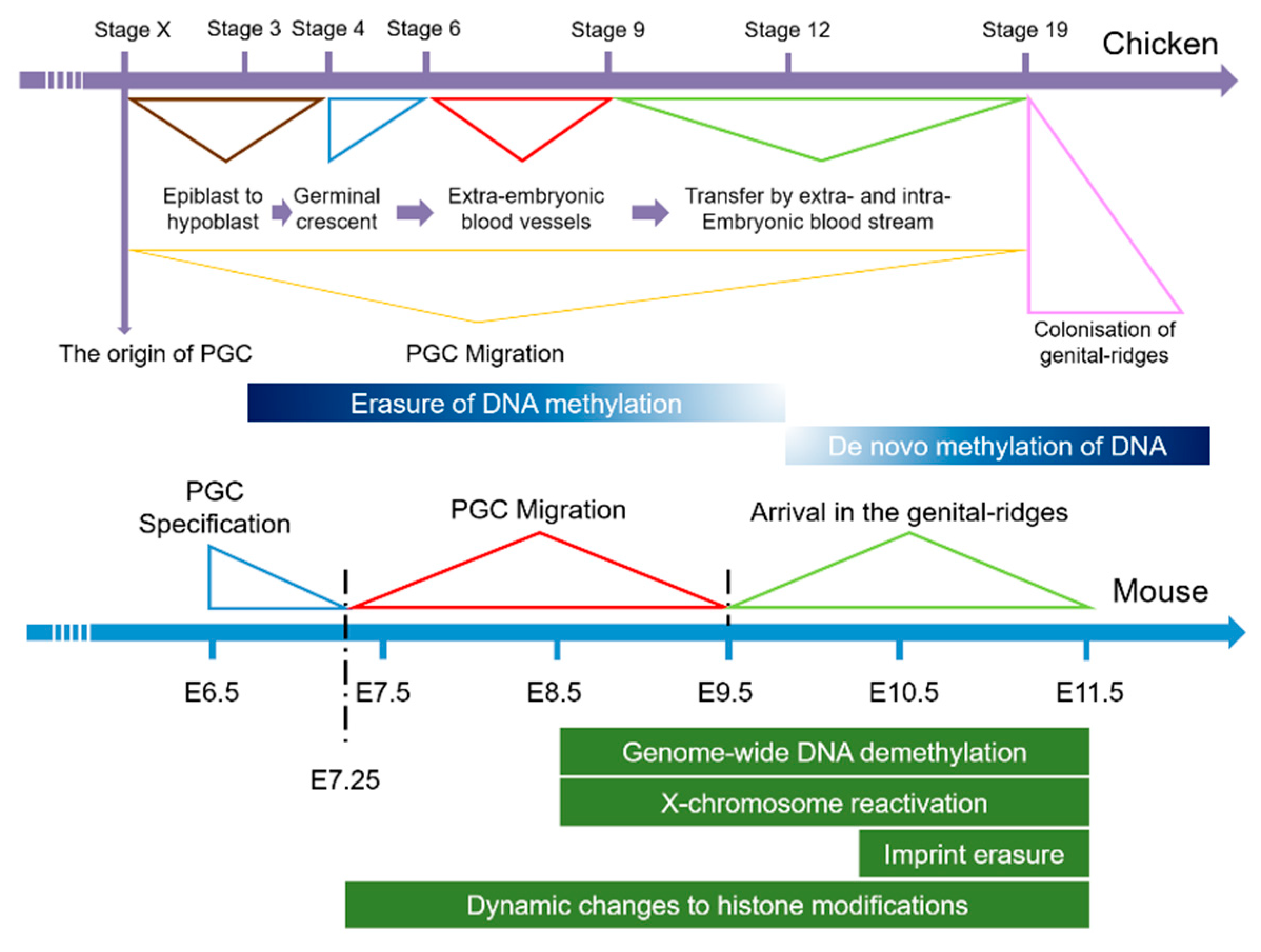
1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Antibody and Reagent
2.3. Experimental Animals
2.4. Cell Culture and Plasmids
2.5. FACS and RNA-seq
2.6. Isolation of Nuclear and Cytoplasmic RNA
2.7. Chicken Embryo Vascular Injection
2.8. qRT-PCR
2.9. Indirect Immunoinfluscent Assay (IFA)
2.10. Dual-Luciferase Assays
2.11. Data Analysis
3. Results
3.1. Searching for LncRNAs Involved in the Development of PGCs
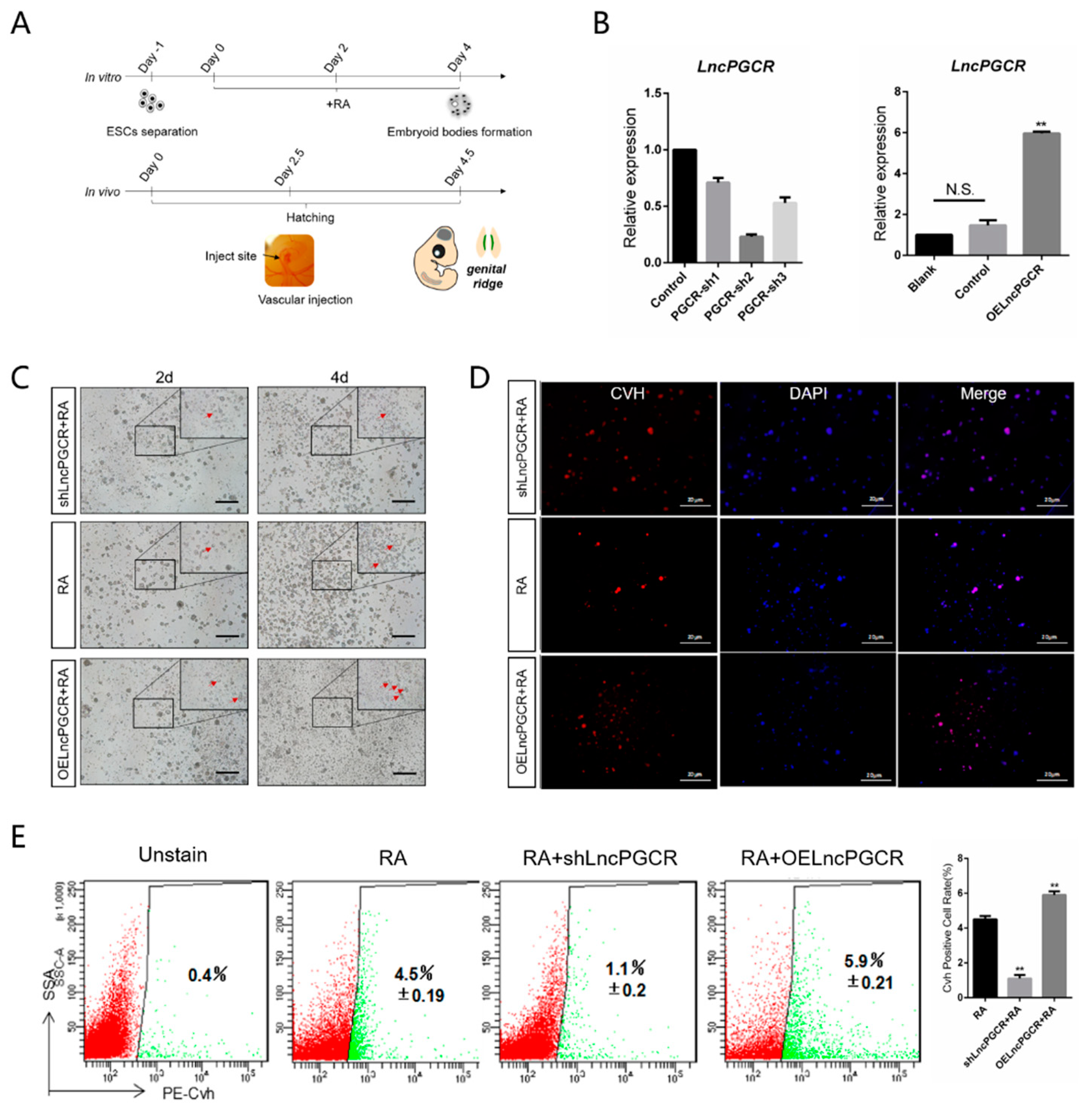
3.2. LncPGCR Promotes Development of PGCs In Vitro and In Vivo
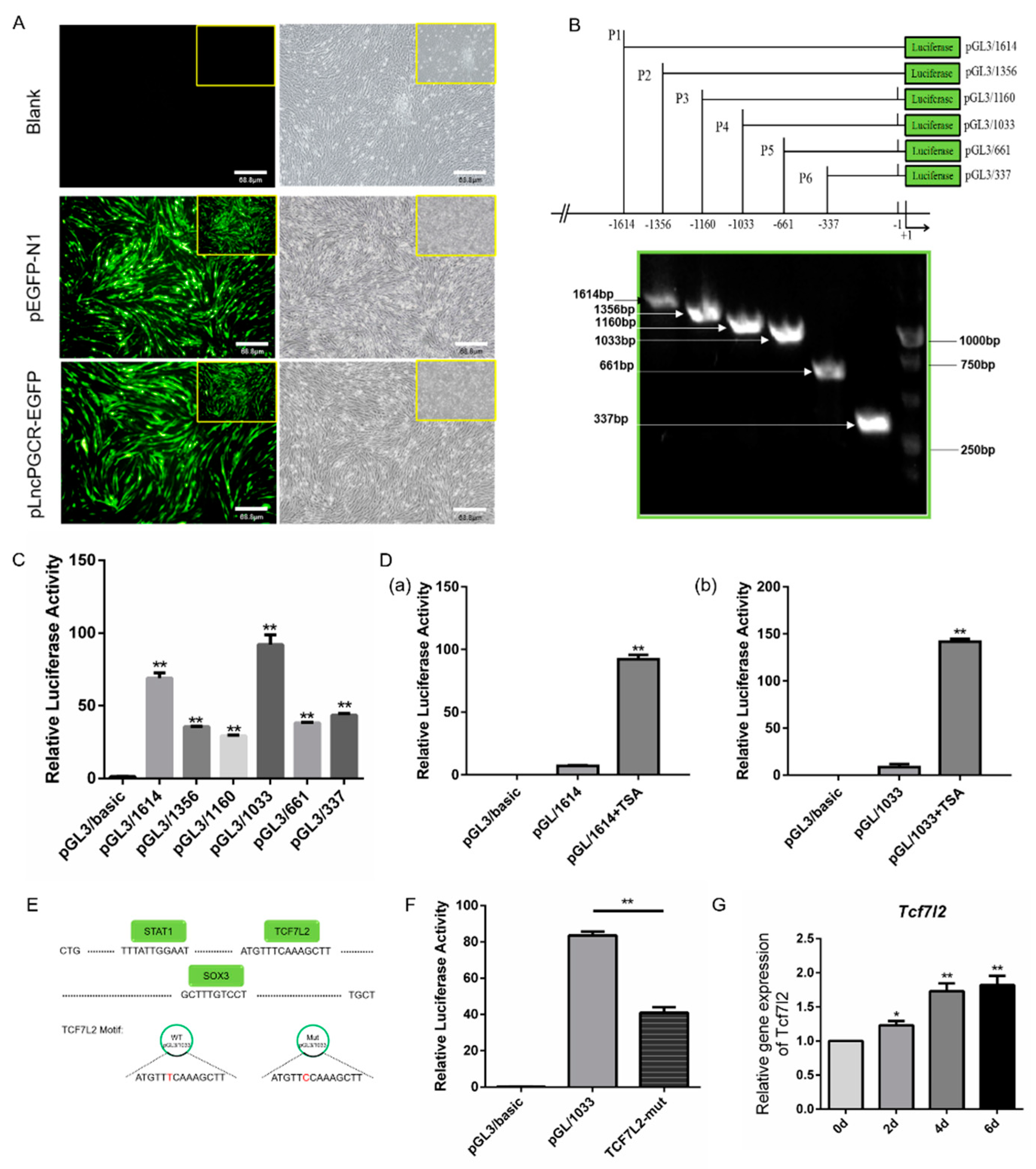
3.3. Deacetylation Downregulate LncPGCR Expression
3.4. Transcription Factor TCF7L2 Regulates the Expression of LncPGCR
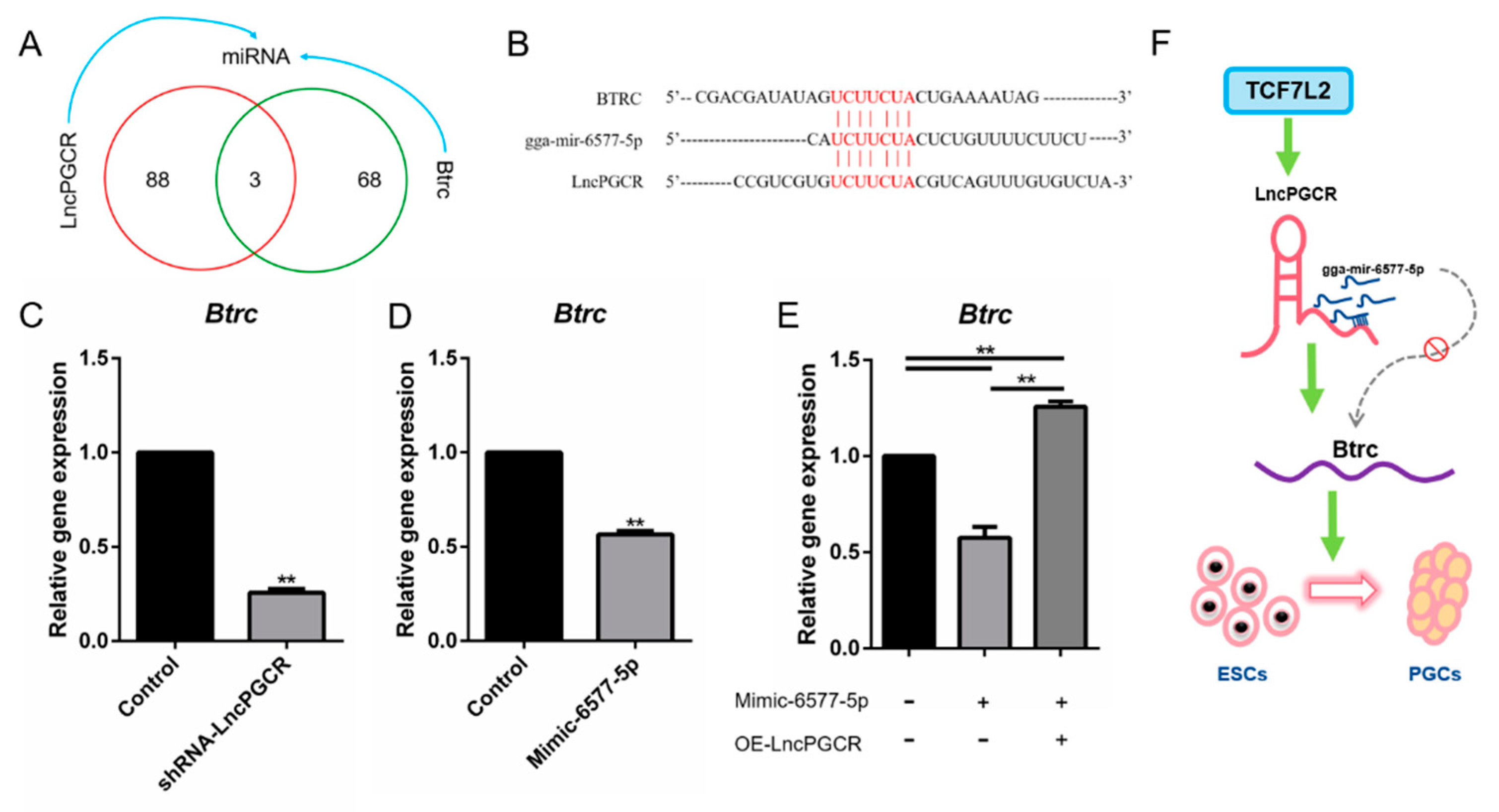
3.5. LncPGCR Functions as a ceRNA for gga-miR-6577-5p
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tsunekawa, N.; Naito, M.; Sakai, Y.; Nishida, T.; Noce, T. Isolation of chicken vasa homolog gene and tracing the origin of primordial germ cells. Development 2000, 127, 2741. [Google Scholar] [PubMed]
- Saitou, M.; Barton, S.C.; Surani, M.A. A molecular programme for the specification of germ cell fate in mice. Nat. Cell Biol. 2002, 418, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.Q.; Garbers, D.L. Male Germ Cell Specification and Differentiation. Dev. Cell 2002, 2, 537–547. [Google Scholar] [CrossRef] [Green Version]
- Ohinata, Y.; Payer, B.; O’Carroll, D.; Ancelin, K.; Ono, Y.; Sano, M.; Barton, S.C.; Obukhanych, T.; Nussenzweig, M.C.; Tarakhovsky, A.; et al. Blimp1 is a critical determinant of the germ cell lineage in mice. Nat. Cell Biol. 2005, 436, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Ancelin, K.; Lange, U.C.; Hajkova, P.; Schneider, R.J.; Bannister, A.J.; Kouzarides, T.; Surani, M.A. Blimp1 associates with Prmt5 and directs histone arginine methylation in mouse germ cells. Nat. Cell Biol. 2006, 8, 623–630. [Google Scholar] [CrossRef]
- Kehler, J.; Tolkunova, E.; Koschorz, B.; Pesce, M.; Gentile, L.; Boiani, M.; Lomelí, H.; Nagy, A.; McLaughlin, K.J.; Schöler, H.R.; et al. Oct4 is required for primordial germ cell survival. EMBO Rep. 2004, 5, 1078–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnúsdóttir, E.; Surani, M.A. How to make a primordial germ cell. Deelopment 2013, 141, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Yamaji, M.; Seki, Y.; Kurimoto, K.; Yabuta, Y.; Yuasa, M.; Shigeta, M.; Yamanaka, K.; Ohinata, Y.; Saitou, M. Critical function of Prdm14 for the establishment of the germ cell lineage in mice. Nat. Genet. 2008, 40, 1016–1022. [Google Scholar] [CrossRef]
- Kallayanee, C.; Svingen, T.; Ng, E.T.; Epp, T.; Spill, C.M.; Clark, C.; Cooper, H.; Koopman, P. Loss of Wnt5a Disrupts Primordial Germ Cell Migration and Male Sexual Development in Mice. Biol. Reprod. 2012, 86, 1–12. [Google Scholar]
- de Sousa Lopes, S.M.C.; Roelen, B.A.J.; Monteiro, R.M.; Emmens, R.; Lin, H.Y.; Li, E.; Lawson, K.A.; Mummery, C.L. BMP signaling mediated by ALK2 in the visceral endoderm is necessary for the generation of primordial germ cells in the mouse embryo. Genes Dev. 2004, 18, 1838–1849. [Google Scholar] [CrossRef] [Green Version]
- Margueron, R.; Trojer, P.; Reinberg, D. The key to development: Interpreting the histone code? Curr. Opin. Genet. Dev. 2005, 15, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Karagenc, L.; Petitte, J.N. Soluble factors and the emergence of chick primordial germ cells in vitro. Poult. Sci. 2000, 79, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Ying, Y.; Qi, X.; Zhao, G.-Q. Induction of primordial germ cells from murine epiblasts by synergistic action of BMP4 and BMP8B signaling pathways. Proc. Natl. Acad. Sci. USA 2001, 98, 7858–7862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seisenberger, S.; Andrews, S.; Krueger, F.; Arand, J.; Walter, J.; Santos, F.; Popp, C.; Thienpont, B.; Dean, W.; Reik, W. The Dynamics of Genome-wide DNA Methylation Reprogramming in Mouse Primordial Germ Cells. Mol. Cell 2012, 48, 849–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, H.; Matsui, Y. Epigenetic events in mammalian germ-cell development: Reprogramming and beyond. Nat. Rev. Genet. 2008, 9, 129–140. [Google Scholar] [CrossRef]
- Chong, S.; Whitelaw, E. Epigenetic germline inheritance. Curr. Opin. Genet. Dev. 2004, 14, 692–696. [Google Scholar] [CrossRef]
- Saitou, M.; Kagiwada, S.; Kurimoto, K. Epigenetic reprogramming in mouse pre-implantation development and primordial germ cells. Development 2012, 139, 15–31. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Sun, C.; Li, D.; Li, T.; Zhang, W.; Jin, J.; Qi-Sheng, Z.; Ya-Ni, Z.; Guo-Hong, C.; Bi-Chun, L. Construction of RNA Interference Vector of Chicken(Gallus domesticus) FGF8 Gene and Its Effect on the Formation of PGCs. J. Agric. Biotechnol. 2018, 26, 1457–1466. [Google Scholar]
- He, N.; Wang, Y.; Zhang, C.; Wang, M.; Wang, Y.; Zuo, Q.; Zhang, Y.; Li, B. Wnt signaling pathway regulates differentiation of chicken embryonic stem cells into spermatogonial stem cells via Wnt5a. J. Cell. Biochem. 2017, 119, 1689–1701. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, M.; Jin, J.; He, N.; Li, T.; Sun, X.; Cai, H.; Shu-Jian, Z.; Chang-Hua, S.; Qi-Sheng, Z.; et al. Construction of RNA Interference Vector of Hsd3β2 Gene and the Regulation of Chicken (Gallus gallus) ESCs Differentiation to PGCs. J. Agric. Biotechnol. 2018, 26, 1567–1576. [Google Scholar]
- Bao, J.; Wu, J.; Schuster, A.S.; Hennig, G.W.; Yan, W. Expression profiling reveals developmentally regulated lncRNA repertoire in the mouse male germline. Biol. Reprod. 2013, 89, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesana, M.; Cacchiarelli, D.; Legnini, I.; Santini, T.; Sthandier, O.; Chinappi, M.; Tramontano, A.; Bozzoni, I. A Long Noncoding RNA Controls Muscle Differentiation by Functioning as a Competing Endogenous RNA. Cell 2011, 147, 358–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, M.-C.; Manor, O.; Wan, Y.; Mosammaparast, N.; Wang, J.K.; Lan, F.; Shi, Y.; Segal, E.; Chang, H.Y. Long Noncoding RNA as Modular Scaffold of Histone Modification Complexes. Science 2010, 329, 689–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guttman, M.; Garber, M.; Levin, J.Z.; Donaghey, J.; Robinson, J.T.; Adiconis, X.; Fan, L.; Koziol, M.J.; Gnirke, A.; Nusbaum, C.; et al. Ab initio reconstruction of cell type–specific transcriptomes in mouse reveals the conserved multi-exonic structure of lincRNAs. Nat. Biotechnol. 2010, 28, 503–510. [Google Scholar] [CrossRef]
- Mohamed, J.S.; Gaughwin, P.M.; Lim, B.; Robson, P.; Lipovich, L. Conserved long noncoding RNAs transcriptionally regulated by Oct4 and Nanog modulate pluripotency in mouse embryonic stem cells. RNA 2009, 16, 324–337. [Google Scholar] [CrossRef] [Green Version]
- Batista, P.J.; Chang, H.Y. Long Noncoding RNAs: Cellular Address Codes in Development and Disease. Cell 2013, 152, 1298–1307. [Google Scholar] [CrossRef] [Green Version]
- Gong, N.; Teng, X.; Li, J.; Liangcd, X.-J. Antisense Oligonucleotide-Conjugated Nanostructure-Targeting lncRNA MALAT1 Inhibits Cancer Metastasis. ACS Appl. Mater. Interfaces 2018, 11, 37–42. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data using Real-Time Quantitative PCR. Method 2002, 25, 402–408. [Google Scholar] [CrossRef]
- Minucci, S.; Pelicci, P.G. Histone deacetylase inhibitors and the promise of epigenetic (and more) treatments for cancer. Nat. Rev. Cancer 2006, 6, 38–51. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, Q.; Wang, J.; Bai, P.; Shi, L.; Shen, W.; Zhou, M.; Zhou, X.; Zhang, Y.; Cai, M. Mit1 Transcription Factor Mediates Methanol Signaling and Regulates the Alcohol Oxidase 1 (AOX1) Promoter inPichia pastoris. J. Biol. Chem. 2016, 291, 6245–6261. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.-P.; Flavell, R.A. The Transcription Factor GATA-3 Is Necessary and Sufficient for Th2 Cytokine Gene Expression in CD4 T Cells. J. Immunol. 2016, 196, 4426–4435. [Google Scholar] [CrossRef] [Green Version]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA Hypothesis: The Rosetta Stone of a Hidden RNA Language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nightingale, K.P.; Wellinger, R.E.; Sogo, J.M.; Becker, P.B. Histone acetylation facilitates RNA polymerase II transcription of the Drosophila hsp26 gene in chromatin. Embo J. 2014, 17, 2865–2876. [Google Scholar] [CrossRef]
- Damcott, C.M.; Pollin, T.I.; Reinhart, L.J.; Ott, S.H.; Shen, H.; Silver, K.D.; Mitchell, B.D.; Shuldiner, A.R. Polymorphisms in the transcription factor 7-like 2 (TCF7L2) gene are associated with type 2 diabetes in the Amish: Replication and evidence for a role in both insulin secretion and insulin resistance. Diabetes 2006, 55, 2654–2659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak, S.H.; Cho, Y.; Moon, M.K.; Kim, J.; Park, B.; Cheong, H.; Shin, H.-D.; Jang, H.; Kim, S.; Lee, H.; et al. Association of polymorphisms in the insulin-degrading enzyme gene with type 2 diabetes in the Korean population. Diabetes Res. Clin. Pr. 2008, 79, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Nie, F.; Wang, Y.; Zhang, Z.; Hou, J.; He, D.; Xie, M.; Xu, L.; De, W.; Wang, Z.-X.; et al. LncRNA HOXA11-AS Promotes Proliferation and Invasion of Gastric Cancer by Scaffolding the Chromatin Modification Factors PRC2, LSD1, and DNMT1. Cancer Res. 2016, 76, 6299–6310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; Cao, G.; Jing, L.; Lin, S.; Wang, X.; Zhang, J.; Wang, M.; Liu, W.; Lv, C. Analysing the relationship between lncRNA and protein-coding gene and the role of lncRNA as ceRNA in pulmonary fibrosis. J. Cell. Mol. Med. 2014, 18, 991–1003. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, Z.; Jiang, J.; Xu, C.; Kang, J.; Xiao, L.; Wu, M.; Xiong, J.; Guo, X.; Liu, H. Endogenous miRNA Sponge lincRNA-RoR Regulates Oct4, Nanog, and Sox2 in Human Embryonic Stem Cell Self-Renewal. Dev. Cell 2013, 25, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Camp, N. WTX is a novel regulator of ubiquitination in the Wnt/beta-catenin and KEAP1/NRF2 pathways. J. Anal. Psychol. 2012, 20, 57–68. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′ to 3′) |
|---|---|
| mir-6577-5p-F | CCGGAATTCGGCTGCTAAGGTATAGCACAGCTGTG |
| mir-6577-5p-R | CCGGGATCCCCTTACCCAGCAGAGCAACTCAG |
| Gene | Primer Sequence |
|---|---|
| β-actin | F: CAGCCATCTTTCTTGGGTAT |
| R: CTGTGATCTCCTTCTGCATCC | |
| Gapdh | F: TCAAATGGGCAGATGCAGGT |
| R: TCAGCAGCAGCCTTCACTAC | |
| U1 | F: ACATGGTGTACAACAAGCGC |
| R: CTCACCGCTCATCGTATCGG | |
| LncPGCR | F: TGGATGGTCAAAGGGATGCC |
| R: ATGAGAAATGGCTGTGGGGG | |
| Cvh | F: CCACGGCTATTTCACACCTCTG |
| R: GCTCTTGGCAAGCATCCGTA | |
| C-kit | F: GCGAACTTCACCTTACCCGATTA |
| R: TGTCATTGCCGAGCATATCCA | |
| Nanog | F: TGGTTTCAGAACCAACGAATGAAG |
| R: TGCACTGGTCACAGCCTGAAG | |
| Btrc | F: TCAAATGGGCAGATGCAGGT |
| R: TCAGCAGCAGCCTTCACTAC | |
| gga-mir-6577-5p | F: acggcgcgCATCTTCTACTCTGTTTTCTTCT |
| R: CAGTGCGTGTCGTGGAGT |
| LncRNA | Target Gene | KEGG |
|---|---|---|
| TCONS_00612668 | BCL9L | -- |
| TCONS_00765266 | Lef-1 | Wnt signaling pathway, Adherens junction, Melanogenesis |
| TCONS_00874170 | BMP4 | TGF-beta signaling pathway, Hedgehog signaling pathway |
| TCONS_00072199 | LIG4 | Non-homologous end-joining |
| TCONS_00362261 | MAPK1 | TGF-beta signaling pathway, MAPK signaling pathway |
| TCONS_00362344 | TBX3 | -- |
| TCONS_00612668 | DDX6 | RNA degradation |
| TCONS_00659989 | NR2E1 | -- |
| TCONS_00946310 | TRIM8 | TGF-beta signaling pathway |
| TCONS_00580256 | SALL4 | -- |
| TCONS_00612668 | PHLDB1 | -- |
| TCONS_00947438 | TCF7L2 | Wnt signaling pathway, Adherens junction, Melanogenesis |
| TCONS_00978804 | ACVR2A | TGF-beta signaling pathway |
| TCONS_00948124 | BTRC | Wnt signaling pathway, Hedgehog signaling pathway, Oocyte meiosis |
| TCONS_00627195 | IGF2BP1 | -- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, J.; Chen, C.; Cheng, S.; Yuan, X.; Jin, J.; Zhang, C.; Sun, X.; Song, J.; Zuo, Q.; Zhang, Y.; et al. Long Noncoding RNA LncPGCR Mediated by TCF7L2 Regulates Primordial Germ Cell Formation in Chickens. Animals 2021, 11, 292. https://doi.org/10.3390/ani11020292
Jiang J, Chen C, Cheng S, Yuan X, Jin J, Zhang C, Sun X, Song J, Zuo Q, Zhang Y, et al. Long Noncoding RNA LncPGCR Mediated by TCF7L2 Regulates Primordial Germ Cell Formation in Chickens. Animals. 2021; 11(2):292. https://doi.org/10.3390/ani11020292
Chicago/Turabian StyleJiang, Jingyi, Chen Chen, Shaoze Cheng, Xia Yuan, Jing Jin, Chen Zhang, Xiaolin Sun, Jiuzhou Song, Qisheng Zuo, Yani Zhang, and et al. 2021. "Long Noncoding RNA LncPGCR Mediated by TCF7L2 Regulates Primordial Germ Cell Formation in Chickens" Animals 11, no. 2: 292. https://doi.org/10.3390/ani11020292